
Diagnostic and Prognostic Markers for Pancreatitis and Pancreatic Ductal Adenocarcinoma
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
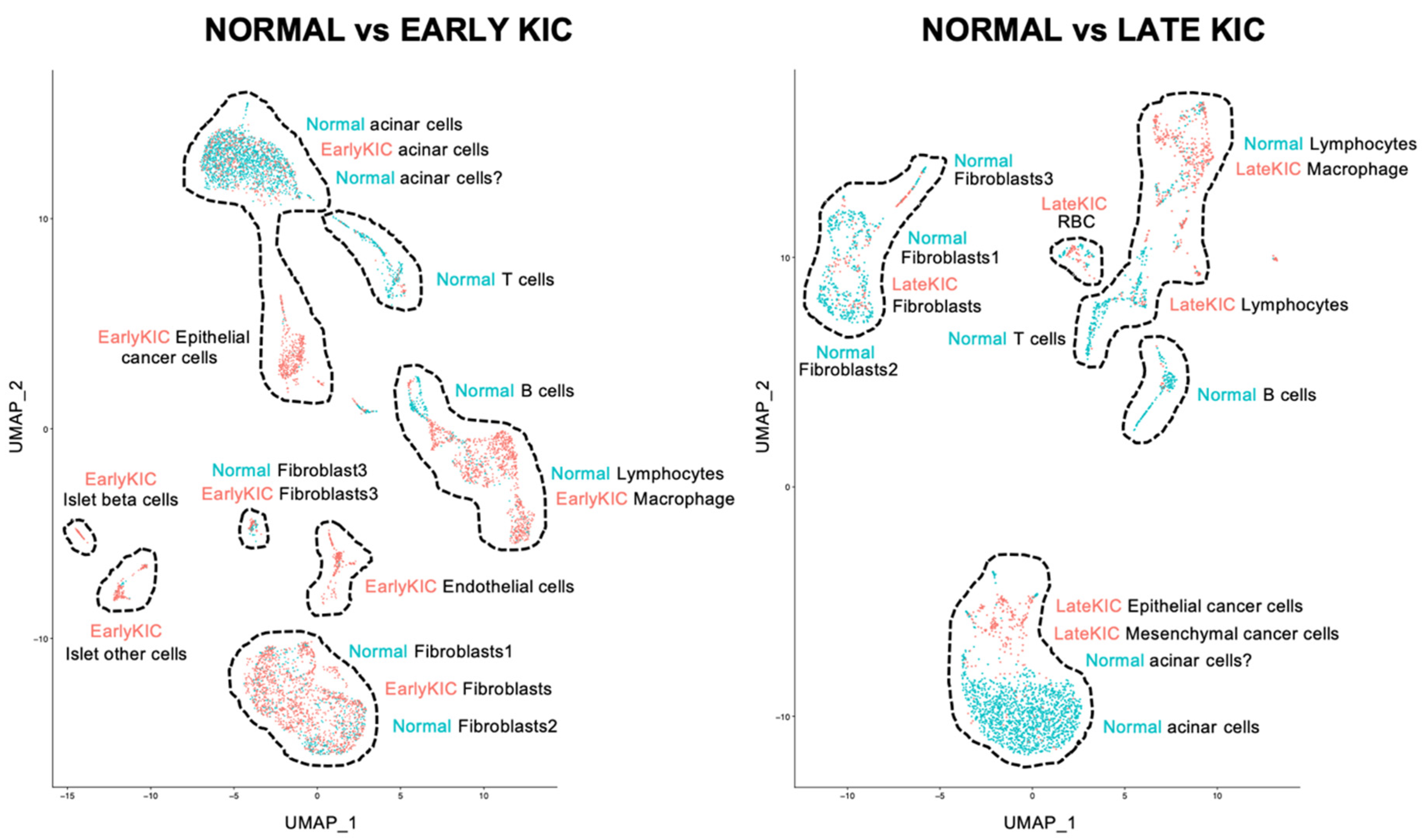
2.1. Single-Cell RNAseq Analysis of PDA in KIC Mice
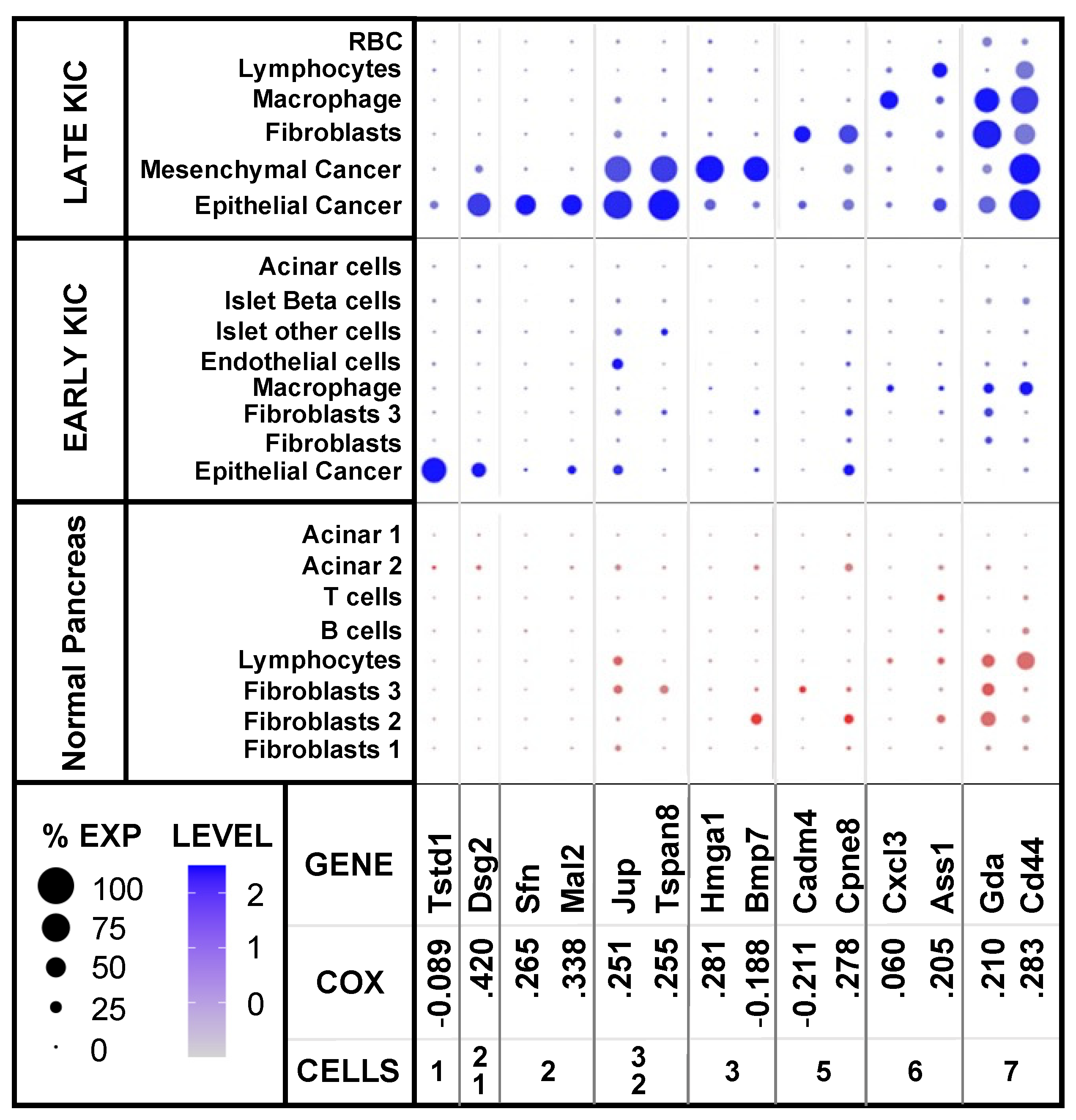
2.2. Cell-Type Expression of Diagnostic PDA Markers
2.3. Screenings of Pathways and Processes Implicated in PDA
2.4. Candidate Gene Identification
2.4.1. Approach (1): Unsupervised Screening for Differentially Expressed PDA Genes in KIC Mice
2.4.2. Approach (2): Mass Spectrometry Identification of Human and Mouse PDA Proteins
2.4.3. Approach (3): Pathways of PDA Initiation and Progression
Ras Pathway Genes
Mutated Genes
KEGG Cancer Genes
Pancreatitis Models
- Calcium pathway genes: Among 1750 genes associated with calcium signaling, 138 were preferentially expressed in specific tumor cell types (Table 1; gene names and cell type expression are in Supplementary Table S1_3N).
- Caerulein: To identify PDA markers that might be differentially expressed in pancreatitis, all candidate PDA marker genes identified above were screened for differential expression in the pancreas of normal mice treated with caerulein; 72 genes were up regulated by the caerulein treatment and 4 genes were down regulated in specific tumor cell types (Table 1; gene names and cell type expression are in Supplementary Table S1_3O and ST1_3P, respectively).
- Differentiation Transcription Factors (dTFs): A total of 154 and 88 genes that were differentially expressed in KIC tumor cells were also elevated or reduced two-fold, respectively, compared to a normal pancreas in dTF KO mice with the acinar-cell-specific deletion of either Ptf1a or Nr5a2 (Table 1; gene names and cell type expression are in Supplementary Table S1_3Q and ST1_3R, respectively).
Innate Immune Genes
2.4.4. Approach (4): Prognostic Markers
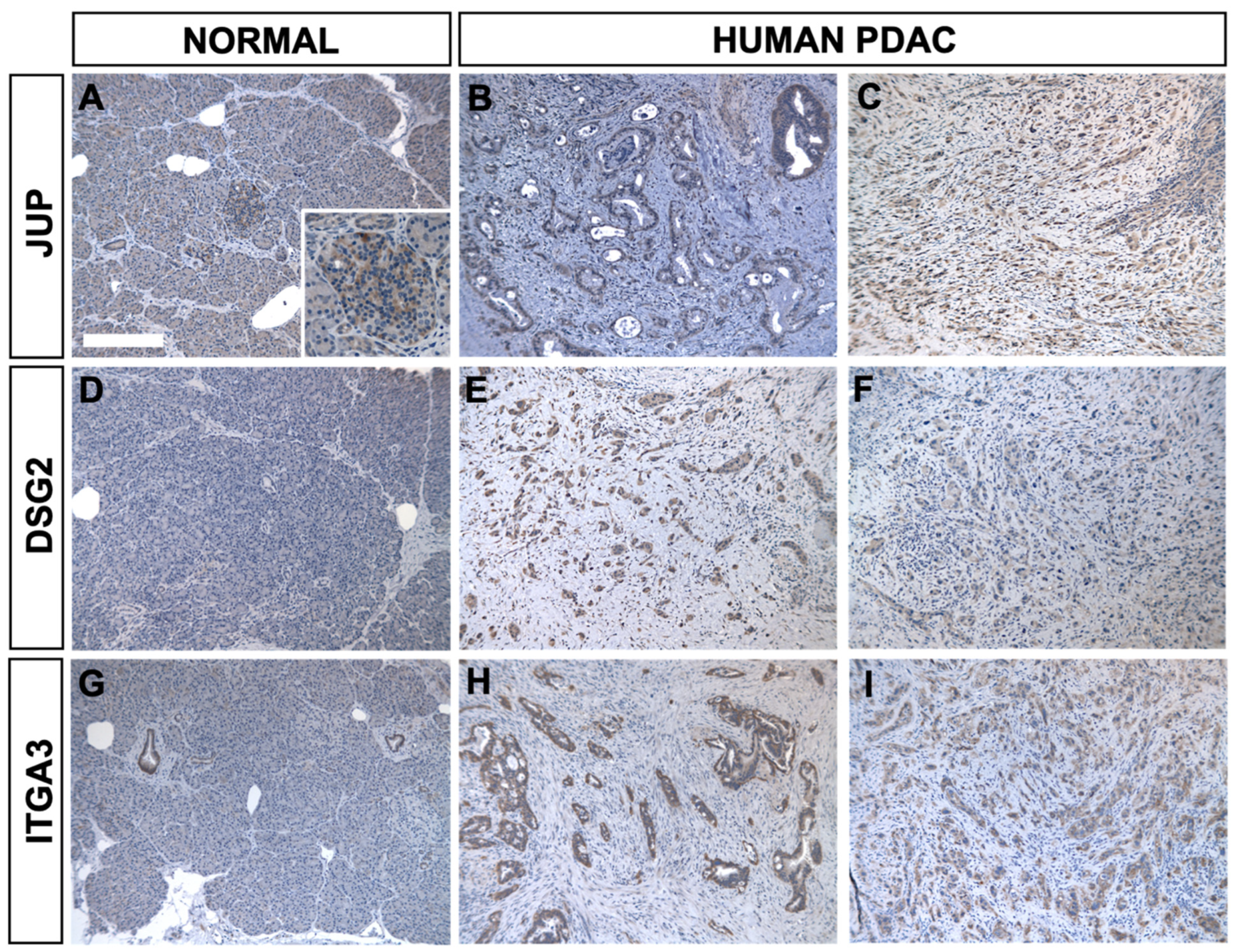
2.5. Immunohistochemistry Validation of PDA Markers
2.6. Pathway Analysis
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Principe, D.R.; Underwood, P.W.; Korc, M.; Trevino, J.G.; Munshi, H.G.; Rana, A. The Current Treatment Paradigm for Pancreatic Ductal Adenocarcinoma and Barriers to Therapeutic Efficacy. Front. Oncol. 2021, 11, 688377. [Google Scholar] [CrossRef] [PubMed]
- Yachida, S.; Iacobuzio-Donahue, C.A. Evolution and dynamics of pancreatic cancer progression. Oncogene 2013, 32, 5253–5260. [Google Scholar] [CrossRef]
- Jones, S.; Zhang, X.; Parsons, D.W.; Lin, J.C.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Kamiyama, H.; Jimeno, A.; et al. Core signaling pathways in human pancreatic cancers revealed by global genomic analyses. Science 2008, 321, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- McWilliams, R.R.; Maisonneuve, P.; Bamlet, W.R.; Petersen, G.M.; Li, D.; Risch, H.A.; Yu, H.; Fontham, E.T.; Luckett, B.; Bosetti, C.; et al. Risk Factors for Early-Onset and Very-Early-Onset Pancreatic Adenocarcinoma: A Pancreatic Cancer Case-Control Consortium (PanC4) Analysis. Pancreas 2016, 45, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.M.; Gingras, M.C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.; Quinn, M.C.; et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, R.A.; Marayati, R.; Flate, E.L.; Volmar, K.E.; Loeza, S.G.; Hoadley, K.A.; Rashid, N.U.; Williams, L.A.; Eaton, S.C.; Chung, A.H.; et al. Virtual microdissection identifies distinct tumor- and stroma-specific subtypes of pancreatic ductal adenocarcinoma. Nat. Genet. 2015, 47, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.J.; Bardeesy, N.; Sinha, M.; Lopez, L.; Tuveson, D.A.; Horner, J.; Redston, M.S.; DePinho, R.A. Activated Kras and Ink4a/Arf deficiency cooperate to produce metastatic pancreatic ductal adenocarcinoma. Genes Dev. 2003, 17, 3112–3126. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Bruick, R.K.; Yu, Y. Secreted IGFBP5 mediates mTORC1-dependent feedback inhibition of IGF-1 signalling. Nat. Cell Biol. 2016, 18, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Hu, R.; Ding, Q.; Savage, T.K.; Huffman, K.E.; Williams, N.; Cobb, M.H.; Minna, J.D.; Johnson, J.E.; Yu, Y. Subtype-specific secretomic characterization of pulmonary neuroendocrine tumor cells. Nat. Commun. 2019, 10, 3201. [Google Scholar] [CrossRef] [PubMed]
- Hosein, A.N.; Huang, H.; Wang, Z.; Parmar, K.; Du, W.; Huang, J.; Maitra, A.; Olson, E.; Verma, U.; Brekken, R.A. Cellular heterogeneity during mouse pancreatic ductal adenocarcinoma progression at single-cell resolution. JCI Insight 2019, 5, e129212. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.; Collado, M.; Navas, C.; Schuhmacher, A.J.; Hernandez-Porras, I.; Canamero, M.; Rodriguez-Justo, M.; Serrano, M.; Barbacid, M. Pancreatitis-induced inflammation contributes to pancreatic cancer by inhibiting oncogene-induced senescence. Cancer Cell 2011, 19, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Guerra, C.; Schuhmacher, A.J.; Canamero, M.; Grippo, P.J.; Verdaguer, L.; Perez-Gallego, L.; Dubus, P.; Sandgren, E.P.; Barbacid, M. Chronic pancreatitis is essential for induction of pancreatic ductal adenocarcinoma by K-Ras oncogenes in adult mice. Cancer Cell 2007, 11, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Hale, M.A.; Swift, G.H.; Hoang, C.Q.; Deering, T.G.; Masui, T.; Lee, Y.K.; Xue, J.; MacDonald, R.J. The nuclear hormone receptor family member NR5A2 controls aspects of multipotent progenitor cell formation and acinar differentiation during pancreatic organogenesis. Development 2014, 141, 3123–3133. [Google Scholar] [CrossRef] [PubMed]
- Xuan, S.; Borok, M.J.; Decker, K.J.; Battle, M.A.; Duncan, S.A.; Hale, M.A.; Macdonald, R.J.; Sussel, L. Pancreas-specific deletion of mouse Gata4 and Gata6 causes pancreatic agenesis. J. Clin. Investig. 2012, 122, 3516–3528. [Google Scholar] [CrossRef] [PubMed]
- Meredith, D.M.; Borromeo, M.D.; Deering, T.G.; Casey, B.H.; Savage, T.K.; Mayer, P.R.; Hoang, C.; Tung, K.C.; Kumar, M.; Shen, C.; et al. Program specificity for Ptf1a in pancreas versus neural tube development correlates with distinct collaborating cofactors and chromatin accessibility. Mol. Cell. Biol. 2013, 33, 3166–3179. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Cerulein pancreatitis: Oxidative stress, inflammation, and apoptosis. Gut Liver 2008, 2, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Molero, X.; Vaquero, E.C.; Flandez, M.; Gonzalez, A.M.; Ortiz, M.A.; Cibrian-Uhalte, E.; Servitja, J.M.; Merlos, A.; Juanpere, N.; Massumi, M.; et al. Gene expression dynamics after murine pancreatitis unveils novel roles for Hnf1alpha in acinar cell homeostasis. Gut 2012, 61, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Cobo, I.; Martinelli, P.; Flandez, M.; Bakiri, L.; Zhang, M.; Carrillo-de-Santa-Pau, E.; Jia, J.; Sanchez-Arevalo Lobo, V.J.; Megias, D.; Felipe, I.; et al. Transcriptional regulation by NR5A2 links differentiation and inflammation in the pancreas. Nature 2018, 554, 533–537. [Google Scholar] [CrossRef]
- Krah, N.M.; De La, O.J.; Swift, G.H.; Hoang, C.Q.; Willet, S.G.; Chen Pan, F.; Cash, G.M.; Bronner, M.P.; Wright, C.V.; MacDonald, R.J.; et al. The acinar differentiation determinant PTF1A inhibits initiation of pancreatic ductal adenocarcinoma. Elife 2015, 4, e07125. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.A.; Bednar, F.; Zhang, Y.; Brisset, J.C.; Galban, S.; Galban, C.J.; Rakshit, S.; Flannagan, K.S.; Adsay, N.V.; Pasca di Magliano, M. Oncogenic Kras is required for both the initiation and maintenance of pancreatic cancer in mice. J. Clin. Investig. 2012, 122, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Singhi, A.D.; McGrath, K.; Brand, R.E.; Khalid, A.; Zeh, H.J.; Chennat, J.S.; Fasanella, K.E.; Papachristou, G.I.; Slivka, A.; Bartlett, D.L.; et al. Preoperative next-generation sequencing of pancreatic cyst fluid is highly accurate in cyst classification and detection of advanced neoplasia. Gut 2018, 67, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Witkiewicz, A.K.; McMillan, E.A.; Balaji, U.; Baek, G.; Lin, W.C.; Mansour, J.; Mollaee, M.; Wagner, K.U.; Koduru, P.; Yopp, A.; et al. Whole-exome sequencing of pancreatic cancer defines genetic diversity and therapeutic targets. Nat. Commun. 2015, 6, 6744. [Google Scholar] [CrossRef] [PubMed]
- Biankin, A.V.; Waddell, N.; Kassahn, K.S.; Gingras, M.C.; Muthuswamy, L.B.; Johns, A.L.; Miller, D.K.; Wilson, P.J.; Patch, A.M.; Wu, J.; et al. Pancreatic cancer genomes reveal aberrations in axon guidance pathway genes. Nature 2012, 491, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.J.; Norris, A.L.; Petersen, G.M.; Bondy, M.L.; Brand, R.; Gallinger, S.; Kurtz, R.C.; Olson, S.H.; Rustgi, A.K.; Schwartz, A.G.; et al. Whole Genome Sequencing Defines the Genetic Heterogeneity of Familial Pancreatic Cancer. Cancer Discov. 2016, 6, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Mann, K.M.; Ward, J.M.; Yew, C.C.; Kovochich, A.; Dawson, D.W.; Black, M.A.; Brett, B.T.; Sheetz, T.E.; Dupuy, A.J.; Australian Pancreatic Cancer Genome, I.; et al. Sleeping Beauty mutagenesis reveals cooperating mutations and pathways in pancreatic adenocarcinoma. Proc. Natl. Acad. Sci. USA 2012, 109, 5934–5941. [Google Scholar] [CrossRef] [PubMed]
- Krah, N.M.; Narayanan, S.M.; Yugawa, D.E.; Straley, J.A.; Wright, C.V.E.; MacDonald, R.J.; Murtaugh, L.C. Prevention and Reversion of Pancreatic Tumorigenesis through a Differentiation-Based Mechanism. Dev. Cell 2019, 50, 744–754 e744. [Google Scholar] [CrossRef]
- Petersen, O.H.; Tepikin, A.V. Polarized calcium signaling in exocrine gland cells. Annu. Rev. Physiol. 2008, 70, 273–299. [Google Scholar] [CrossRef]
- Flandez, M.; Cendrowski, J.; Canamero, M.; Salas, A.; del Pozo, N.; Schoonjans, K.; Real, F.X. Nr5a2 heterozygosity sensitises to, and cooperates with, inflammation in KRas(G12V)-driven pancreatic tumourigenesis. Gut 2014, 63, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Azevedo-Pouly, A.; Hale, M.A.; Swift, G.H.; Hoang, C.Q.; Deering, T.G.; Xue, J.; Wilkie, T.M.; Murtaugh, L.C.; MacDonald, R.J. Key transcriptional effectors of the pancreatic acinar phenotype and oncogenic transformation. PLoS ONE 2023, 18, e0291512. [Google Scholar] [CrossRef] [PubMed]
- Cobo, I.; Iglesias, M.; Flandez, M.; Verbeke, C.; Del Pozo, N.; Llorente, M.; Lawlor, R.; Luchini, C.; Rusev, B.; Scarpa, A.; et al. Epithelial Nr5a2 heterozygosity cooperates with mutant Kras in the development of pancreatic cystic lesions. J. Pathol. 2021, 253, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Reichert, M.; Rustgi, A.K. Pancreatic ductal cells in development, regeneration, and neoplasia. J. Clin. Investig. 2011, 121, 4572–4578. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Curbelo, D.; Ho, Y.J.; Burdziak, C.; Maag, J.L.V.; Morris, J.P.t.; Chandwani, R.; Chen, H.A.; Tsanov, K.M.; Barriga, F.M.; Luan, W.; et al. A gene-environment-induced epigenetic program initiates tumorigenesis. Nature 2021, 590, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wang, M.; Wang, C. Qingyihuaji formula reverses gemcitabine resistant human pancreatic cancer through regulate lncRNA AB209630/miR-373/EphB2-NANOG signals. Biosci. Rep. 2019, 39, BSR20190610. [Google Scholar] [CrossRef] [PubMed]
- Oweida, A.; Bhatia, S.; Hirsch, K.; Calame, D.; Griego, A.; Keysar, S.; Pitts, T.; Sharma, J.; Eckhardt, G.; Jimeno, A.; et al. Ephrin-B2 overexpression predicts for poor prognosis and response to therapy in solid tumors. Mol. Carcinog. 2017, 56, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zhang, Y.; Li, Z.; Yu, S.; Zhao, G.; Li, M.; Wang, Z.; Wang, Q.; Yang, Y. Overexpression of the B-type Eph and ephrin genes correlates with progression and pain in human pancreatic cancer. Oncol. Lett. 2012, 3, 1207–1212. [Google Scholar] [CrossRef] [PubMed]
- Papadakos, S.P.; Dedes, N.; Gkolemi, N.; Machairas, N.; Theocharis, S. The EPH/Ephrin System in Pancreatic Ductal Adenocarcinoma (PDAC): From Pathogenesis to Treatment. Int. J. Mol. Sci. 2023, 24, 3015. [Google Scholar] [CrossRef] [PubMed]
- Yachida, S.; White, C.M.; Naito, Y.; Zhong, Y.; Brosnan, J.A.; Macgregor-Das, A.M.; Morgan, R.A.; Saunders, T.; Laheru, D.A.; Herman, J.M.; et al. Clinical significance of the genetic landscape of pancreatic cancer and implications for identification of potential long-term survivors. Clin. Cancer Res. 2012, 18, 6339–6347. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Li, Y.; Liu, S.; Chen, Q.; Liu, Y. ITGA3 serves as a diagnostic and prognostic biomarker for pancreatic cancer. Onco. Targets Ther. 2019, 12, 4141–4152. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Fan, J.; Yang, J.; Zhu, G.Z. Characterization of GPR56 protein and its suppressed expression in human pancreatic cancer cells. Mol. Cell. Biochem. 2008, 308, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Navas, C.; Hernandez-Porras, I.; Schuhmacher, A.J.; Sibilia, M.; Guerra, C.; Barbacid, M. EGF receptor signaling is essential for k-ras oncogene-driven pancreatic ductal adenocarcinoma. Cancer Cell 2012, 22, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Wang, Y.; Cao, L.; Ouellette, M.M.; Freeman, J.W. Expression of oncogenic K-ras and loss of Smad4 cooperate to induce the expression of EGFR and to promote invasion of immortalized human pancreas ductal cells. Int. J. Cancer 2010, 127, 2076–2087. [Google Scholar] [CrossRef] [PubMed]
- Ardito, C.M.; Gruner, B.M.; Takeuchi, K.K.; Lubeseder-Martellato, C.; Teichmann, N.; Mazur, P.K.; Delgiorno, K.E.; Carpenter, E.S.; Halbrook, C.J.; Hall, J.C.; et al. EGF receptor is required for KRAS-induced pancreatic tumorigenesis. Cancer Cell 2012, 22, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Balci, H.; Hanspers, K.; Coort, S.L.; Martens, M.; Slenter, D.N.; Ehrhart, F.; Digles, D.; Waagmeester, A.; Wassink, I.; et al. WikiPathways 2024: Next generation pathway database. Nucleic Acids Res. 2024, 52, D679–D689. [Google Scholar] [CrossRef] [PubMed]
- Milacic, M.; Beavers, D.; Conley, P.; Gong, C.; Gillespie, M.; Griss, J.; Haw, R.; Jassal, B.; Matthews, L.; May, B.; et al. The Reactome Pathway Knowledgebase 2024. Nucleic Acids Res. 2024, 52, D672–D678. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.G.; Yan, G.R.; He, Q.Y. DOSE: An R/Bioconductor package for disease ontology semantic and enrichment analysis. Bioinformatics 2015, 31, 608–609. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Supplementary Table S1 | Query a | Total Genes # | KIC Cell Type Classification | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| E.epi | EL.epi | EL.epi + mes | L.epi | L.e + m | L.mes | EL.fib | EL.h | EL.all | |||
| 1 | 1 + 2 | 1 + 2 + 3 | 2 | 2 + 3 | 3 | 5 | 6 | 7 | |||
| ST1_1C | NvEvL unsup | 503 | 7 | 34 | 9 | 12 | 98 | 56 | 7 | 47 | 233 |
| ST1_2D | mMS | 53 | 2 | 4 | 0 | 5 | 5 | 2 | 5 | 30 | nd |
| ST1_2E | msMS | 209 | 4 | 9 | 5 | 37 | 61 | 10 | 10 | 73 | nd |
| ST1_2F | hMS | 187 | 4 | 15 | 7 | 34 | 57 | 17 | 16 | 37 | nd |
| ST1_3G | Ras Path | 197 | 0 | 7 | 3 | 25 | 65 | 18 | 18 | 62 | nd |
| ST1_3H | Mutant cBP | 16 | 0 | 2 | 0 | 3 | 7 | 4 | 0 | 0 | nd |
| ST1_3I | AMP cBP | 13 | 0 | 4 | 1 | 0 | 4 | 4 | 0 | 0 | nd |
| ST1_3J | Del cBP | 17 | 1 | 4 | 1 | 1 | 4 | 6 | 0 | 0 | nd |
| ST1_3K | SB | 17 | 0 | 1 | 2 | 4 | 9 | 0 | 0 | 1 | nd |
| ST1_3L | Tempus + FM | 191 | 12 | 6 | 16 | 47 | 50 | 23 | 9 | 28 | nd |
| ST1_3M | KEGG cancer | 66 | 0 | 0 | 1 | 5 | 33 | 9 | 4 | 14 | nd |
| ST1_3N | Ca2+ Path | 138 | 1 | 5 | 3 | 24 | 54 | 19 | 8 | 24 | nd |
| ST1_3O | Caer Up | 72 | 1 | 7 | 3 | 9 | 26 | 2 | 5 | 15 | nd |
| ST1_3P | Caer Down | 4 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | nd |
| ST1_3Q | ko dTF Up | 154 | 5 | 20 | 6 | 25 | 40 | 15 | 5 | 38 | nd |
| ST1_3R | ko dTF Down | 88 | 4 | 6 | 5 | 12 | 23 | 10 | 8 | 20 | nd |
| ST1_3S | Innate Imm | 46 | 2 | 1 | 0 | 8 | 3 | 5 | 5 | 22 | nd |
| ST1_3T | ITGA3-Gpr56 | 128 | 1 | 5 | 1 | 20 | 54 | 14 | 14 | 19 | nd |
| ST1_4U | Cox Hi_500 | 202 | 0 | 4 | 2 | 21 | 53 | 6 | 3 | 11 | 102 |
| ST1_4W | Cox Lo_500 | 191 | 0 | 3 | 0 | 13 | 21 | 3 | 4 | 7 | 140 |
| ST1_Y | Panc CT_ID | 41 | 0 | 1 | 2 | 0 | 0 | 0 | 4 | 12 | 22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kantheti, H.S.; Hale, M.A.; Pal Choudhuri, S.; Huang, H.; Wang, X.-d.; Zolghadri, Y.; Innamorati, G.; Manikonda, S.P.R.; Reddy, N.; Reddy, S.; et al. Diagnostic and Prognostic Markers for Pancreatitis and Pancreatic Ductal Adenocarcinoma. Int. J. Mol. Sci. 2024, 25, 6619. https://doi.org/10.3390/ijms25126619
Kantheti HS, Hale MA, Pal Choudhuri S, Huang H, Wang X-d, Zolghadri Y, Innamorati G, Manikonda SPR, Reddy N, Reddy S, et al. Diagnostic and Prognostic Markers for Pancreatitis and Pancreatic Ductal Adenocarcinoma. International Journal of Molecular Sciences. 2024; 25(12):6619. https://doi.org/10.3390/ijms25126619
Chicago/Turabian StyleKantheti, Havish S., Michael A. Hale, Shreoshi Pal Choudhuri, Huocong Huang, Xu-dong Wang, Yalda Zolghadri, Giulio Innamorati, Sai Prasada Rao Manikonda, Naviya Reddy, Sarthak Reddy, and et al. 2024. "Diagnostic and Prognostic Markers for Pancreatitis and Pancreatic Ductal Adenocarcinoma" International Journal of Molecular Sciences 25, no. 12: 6619. https://doi.org/10.3390/ijms25126619