

The Thioredoxin Fold Protein (TFP2) from Extreme Acidophilic Leptospirillum sp. CF-1 Is a Chaperedoxin-like Protein That Prevents the Aggregation of Proteins under Oxidative Stress

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Results
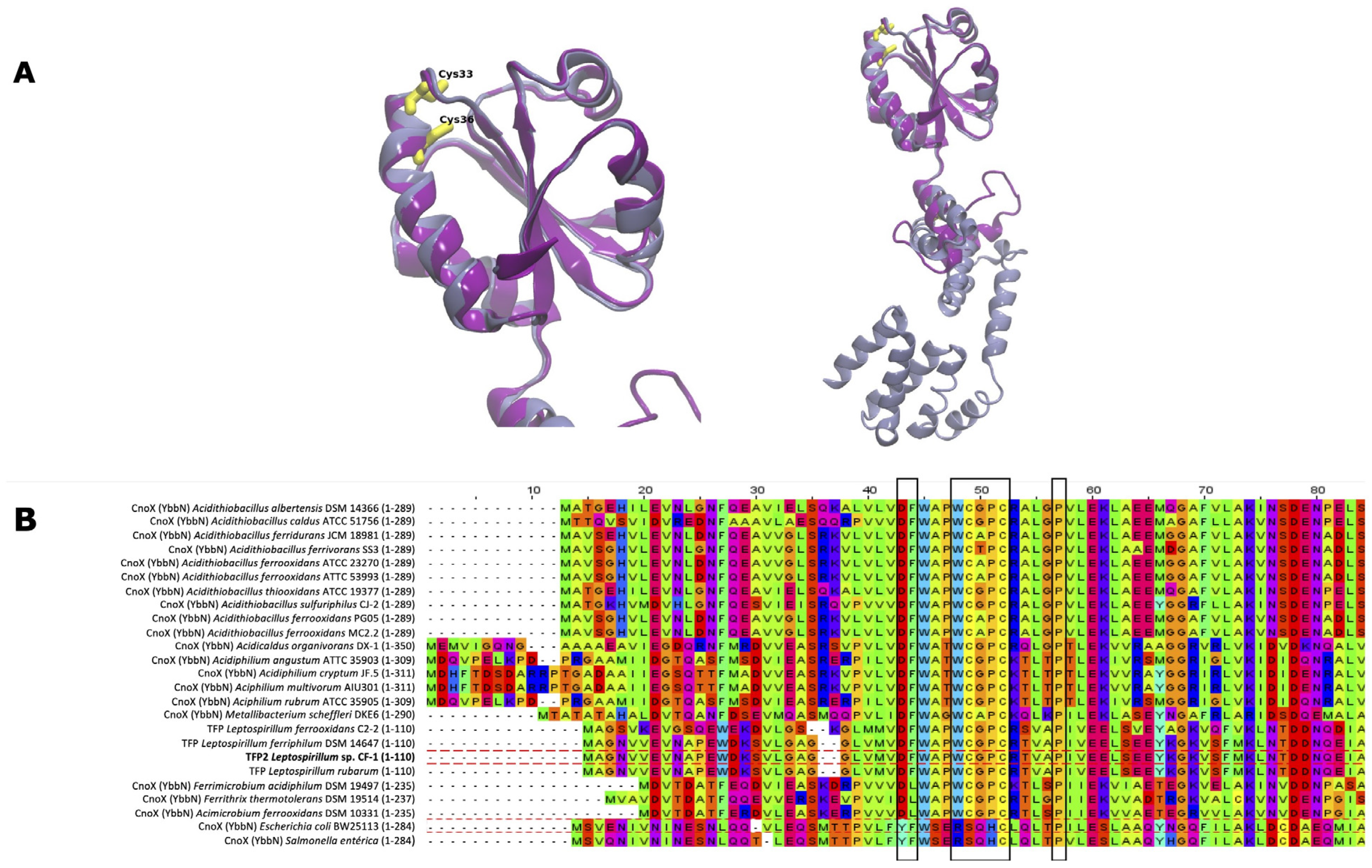
2.1. Structural and Multiple Sequence Alignment Analysis of tfp2 from Leptospirillum sp. CF-1
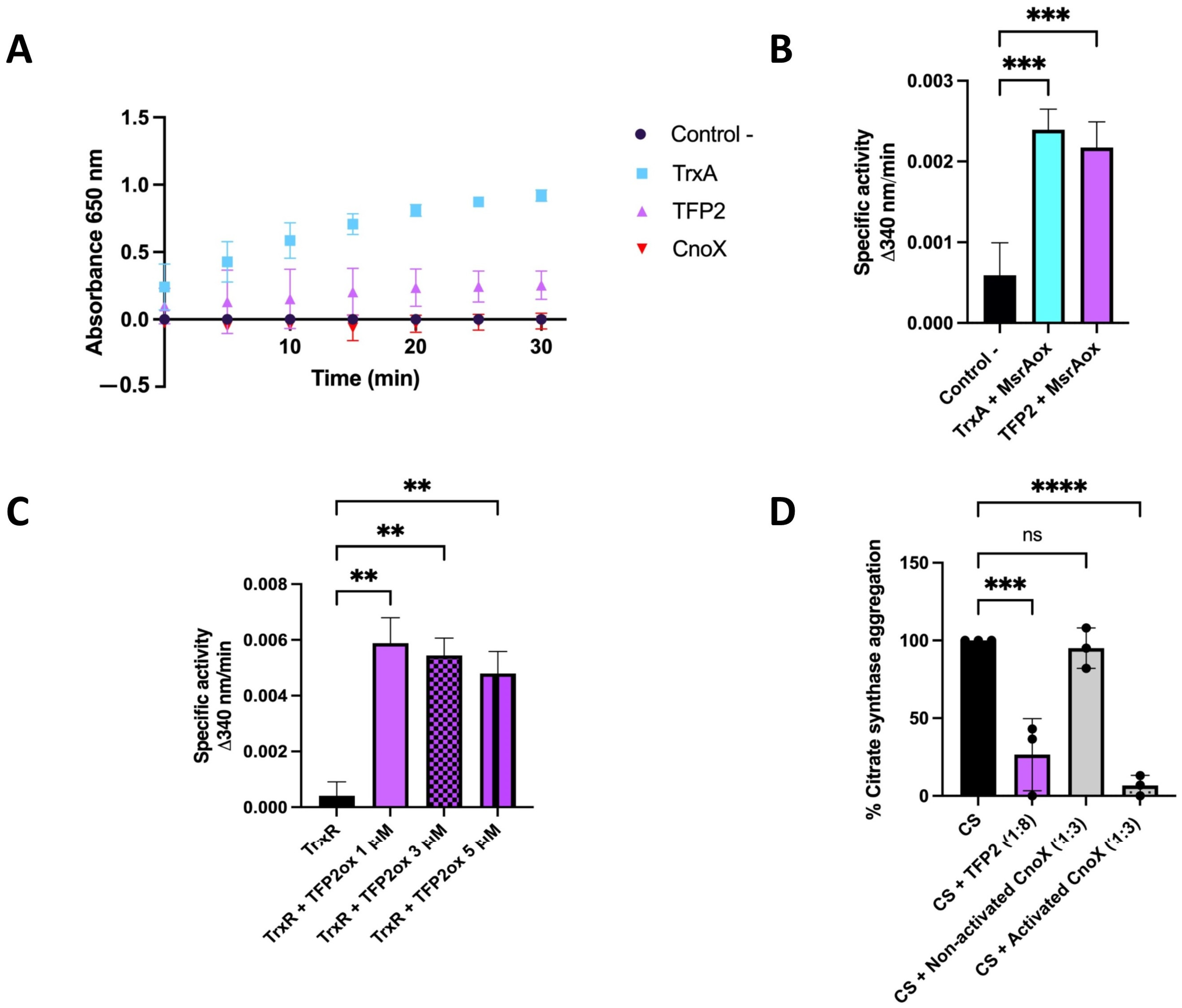
2.2. Evaluation of Reductase and Chaperone Activities of TFP2 In Vitro
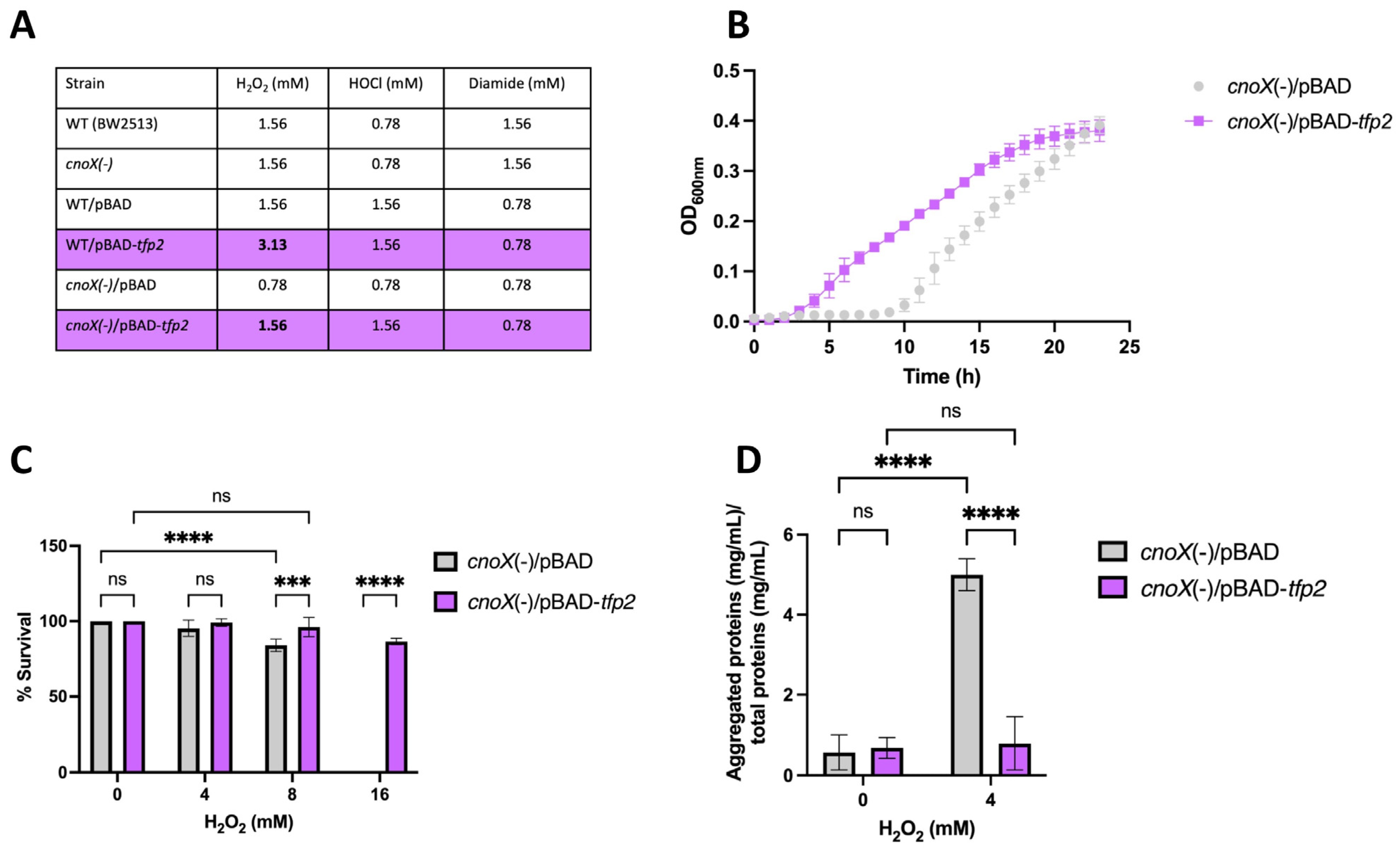
2.3. Role of TFP2 in Protecting E. coli against Oxidative Stress
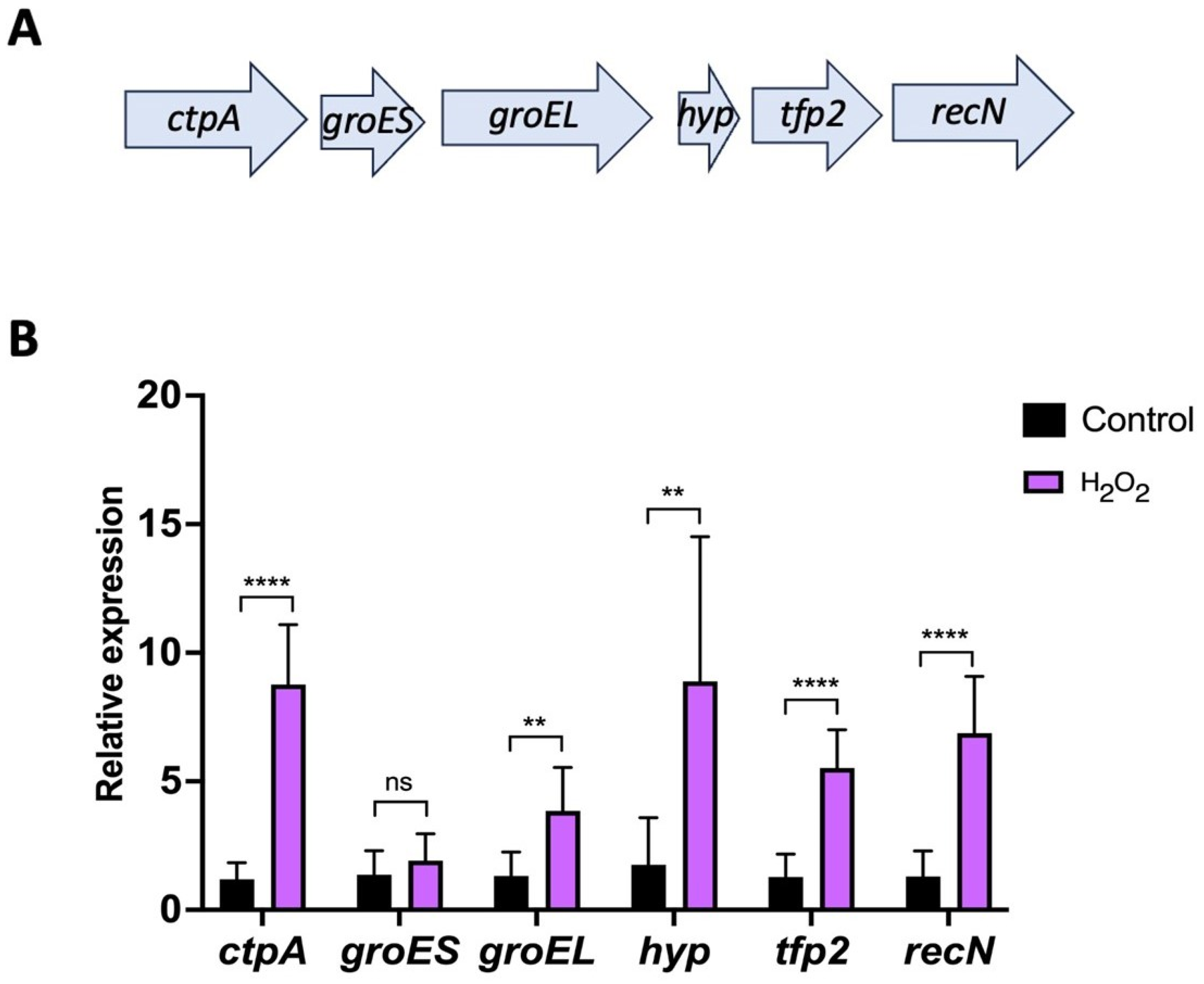
2.4. Genetic Organization and Expression of tfp2 Gene and Neighboring Genes
2.5. Identification of TFP2 Client Proteins
3. Discussion
4. Materials and Methods
4.1. Structural Analysis
4.2. Multiple Sequence Alignment Analysis
4.3. Bacterial Strains, Growth, and Viability
4.4. Susceptibility Assay of E. coli to H2O2
4.5. Cloning in pet21b Expression Vector
4.6. Protein Purification
4.7. Thioredoxin Activity Assay
4.8. Thiol Reductase Assay
4.9. Reduction of TFP2 by TrxR
4.10. Determination of Chaperone Activity
4.11. Cloning pBAD-tfp2 in E. coli
4.12. Protein Aggregation Measurement
4.13. Relative Expression of tfp2 and Neighboring Genes under Oxidative Conditions
4.14. Identification of TFP2 Client Proteins in E. coli
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imlay, J.A. Diagnosing oxidative stress in bacteria: Not as easy as you might think. Curr. Opin. Microbiol. 2015, 24, 124–131. [Google Scholar] [CrossRef]
- Davies, M.J. The oxidative environment and protein damage. Biochem. Biophys. Acta 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Reichmann, D.; Voth, W.; Jakob, U. Maintaining a healthy proteome during oxidative stress. Mol. Cell. 2018, 69, 203–213. [Google Scholar] [CrossRef]
- Dahl, J.U.; Gray, M.J.; Jakob, U. Protein quality control under oxidative stress conditions. J. Mol. Biol. 2015, 427, 1549–1563. [Google Scholar] [CrossRef]
- Balchin, D.; Hayer-Hartl, M.; Hartl, F.U. Recent advances in understanding catalysis of protein folding by molecular chap-erones. FEBS Lett. 2020, 594, 2770–2781. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Bröms, J.E.; Kumar, R.; Sjöstedt, A. The role of ClpB in bacterial stress responses and virulence. Front. Mol. Biosci. 2021, 8, 668910. [Google Scholar] [CrossRef]
- Khor, H.K.; Fisher, M.T.; Schoneich, C. Potential role of methionine sulfoxide in the inactivation of the chaperone GroEL by hypochlorous acid (HOCl) and peroxynitrite (ONOO−). J. Biol. Chem. 2004, 279, 19486–19493. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Linke, K.; Jatzek, A.; Jakob, U. Severe oxidative stress causes inactivation of DnaK and activation of the re-dox-regulated chaperone Hsp33. Mol Cell. 2005, 17, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Goemans, C.V.; Collet, J.F. Stress-induced chaperones: A first line of defense against the powerful oxidant hypochlorous acid. F1000Research 2019, 8. [Google Scholar] [CrossRef]
- Goemans, C.V.; Vertommen, D.; Agrebi, R.; Collet, J. CnoX is a chaperedoxin: A holdase that protects its substrates from irreversible oxidation. Mol. Cell 2018, 70, 614–627. [Google Scholar] [CrossRef]
- Lin, J.; Wilson, M.A. Escherichia coli thioredoxin-like protein YbbN contains an atypical tetratricopeptide repeat motif and is negative regulator of GroEL. J. Biol. Chem. 2011, 286, 19459–19469. [Google Scholar] [CrossRef] [PubMed]
- Collet, J.F.; Messens, J. Structure, function, and mechanism of thioredoxin proteins. Antioxid. Redox Signal 2010, 13, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Allan, R.K.; Ratajczak, T. Versatile TPR domains accommodate different modes of target protein recognition and function. Cell Stress Chaperones 2011, 16, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.; Schippers, A.; Hedrich, S.; Sand, W. Progress in bioleaching: Fundamentals and mechanisms of microbial metal sulfide oxidation-part A. App. Microbiol. Biotechnol. 2002, 106, 6933–6952. [Google Scholar] [CrossRef] [PubMed]
- Dopson, M.; Ossandon, F.J.; Lövgren, L.; Holmes, D.S. Metal resistance or tolerance? Acidophiles confront high metal loads via both abiotic and biotic mechanisms. Front. Microbiol. 2014, 5, 157. [Google Scholar] [CrossRef] [PubMed]
- Schoonen, M.A.A.; Harrington, A.D.; Laffers, R.; Strongin, D.R. Role of hydrogen peroxide and hydroxyl radical in pyrite oxidtion by molecular oxygen. Geochim. Cosmochim. 2010, 74, 4971–4987. [Google Scholar] [CrossRef]
- Nooshabadi, A.J.; Larsson, A.-C.; Kota, H.R. Formation of hydrogen peroxide by chalcopyrite and its influence on flotation. Min. Eng. 2013, 49, 128–134. [Google Scholar] [CrossRef]
- Zhou, S.; Gan, M.; Wang, X.; Zhang, Y.; Fang, Y.; Gu, G.; Wang, Y.; Qiu, G. ROS formation driven by pyrite-mediated arse-nopyrite oxidation and its potential role on arsenic transformation. J. Hazard. Mater. 2023, 443, 130151. [Google Scholar] [CrossRef]
- Rivera-Araya, J.; Pollender, A.; Huynh, D.; Schlömann, M.; Chávez, R.; Levicán, G. Osmotic imbalance, cytoplasm acidification and oxidative stress induction support the high toxicity of chloride in acidophilic bacteria. Front. Microbiol. 2019, 10, 2455. [Google Scholar] [CrossRef]
- Norambuena, J.; Flores, R.; Cárdenas, J.P.; Quatrini, R.; Chávez, R.; Levicán, G. Thiol/disulfide system plays a crucial role in redox protection in the acidophilic iron-oxidizing bacterium Leptospirillum ferriphilum. PLoS ONE 2012, 7, e44576. [Google Scholar] [CrossRef]
- González, D.; Álamos, P.; Rivero, M.; Orellana, O.; Norambuena, J.; Chávez, R.; Levicán, G. Deciphering the role of multiple thioredoxin fold proteins of Leptospirillum sp. in oxidative stress tolerance. Int. J. Mol. Sci. 2020, 21, 1880. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Zhang, Y.; Trinidad, J.C.; Giedroc, D.P. Thioredoxin profiling of multiple thioredoxin-like proteins in Staphylococcus aureus. Front. Microbiol. 2018, 9, 410891. [Google Scholar] [CrossRef] [PubMed]
- Arnér, E.S.; Holmgren, A. Measurement of thioredoxin and thioredoxin reductase. Curr. Protoc. Toxicol. 2001, 24, 7.4.4–7.4.14. [Google Scholar] [CrossRef] [PubMed]
- Goemans, C.V.; Beaufay, F.; Arts, I.S.; Agrebi, R.; Vertommen, D.; Collet, J.F. The chaperone and redox properties of CnoX chaperedoxins are tailored to the proteostatic needs of bacterial species. mBio 2018, 9, e01541-18. [Google Scholar] [CrossRef] [PubMed]
- Le, H.T.; Gautier, V.; Kthiri, F.; Kohiyama, M.; Katayama, T.; Rich-arme, G. DNA replication defects in a mutant deficient in the thioredoxin homolog YbbN. Biochem. Biophys. Res. Commun 2011, 405, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Zapata, C.; Paillavil, B.; Chávez, R.; Álamos, P.; Levicán, G. Cytochrome c peroxidase (CcP) is a molecular determinant of the oxidative stress response in the extreme acidophilic Leptospirillum sp. CF-1. FEMS Microbiol Ecol. 2017, 93, fix001. [Google Scholar] [CrossRef] [PubMed]
- Sousa, S.F.; Neves, R.P.P.; Waheed, S.O.; Fernandes, P.P.; Ramos, M.J. Structural and mechanistic aspects of S-S bonds in the thioredoxin-like family of proteins. Biol. Chem. 2019, 400, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Arts, I.S.; Vertommen, D.; Baldin, F.; Laloux, G.; Collet, J.F. Comprehensively characterizing the thioredoxin interactome in vivo highlights the central role played by this ubiquitous oxidoreductase in redox control. Mol. Cell Proteom. 2016, 15, 2125–2140. [Google Scholar] [CrossRef] [PubMed]
- Garcia–Pino, A.; Martinez–Rodriguez, S.; Wahni, K.; Wyns, L.; Loris, R.; Messens, J. Coupling of domain swapping to kinetic stability in a thioredoxin mutant. J. Mol. Biol. 2009, 385, 1590–1599. [Google Scholar] [CrossRef]
- Eklund, H.; Gleason, F.K.; Holmgren, A. Structural and functional relations among thioredoxins of different species. Proteins 1991, 11, 13–28. [Google Scholar] [CrossRef]
- Moskovitz, J.; Bar-Noy, S.; Williams, W.M.; Requena, J.; Berlett, B.S.; Stadtman, E.R. Methionine sulfoxide reductase (MsrA) is a regulator of antioxidant defense and lifespan in mammals. Proc. Natl. Acad. Sci. USA 2001, 98, 12920–12925. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-induced damage to proteins: Formation of nitrogen-centred radicals from lysine residues and their role in protein fragmentation. Biochem. J. 1998, 332, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.L.; Hjerde, E.; Paulsen, S.M.; Hansen, H.; Olsen, L.; Thode, S.K.; Dos Santos, M.T.; Paulssen, R.H.; Willassen, N.-P.; Haugen, P. Global responses of Aliivibrio salmonicida to hydrogen peroxide as revealed by microarray analysis. Mar. Genom. 2010, 3, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Uranga, L.; Reyes, E.; Patidar, P.; Reedman, L.N.; Lusetti, S.L. The cohesin-like RecN protein stimulates RecA-mediated recom-binational repair of DNA double-strand breaks. Nat. Commun. 2017, 8, 15282. [Google Scholar] [CrossRef]
- Hsu, H.C.; Wang, M.; Kovach, A.; Darwin, A.J.; Li, H. Pseudomonas aeruginosa C-terminal processing protease CtpA assembles into a hexameric structure that requires activation by a spiral-shaped lipoprotein-binding partner. Mbio 2022, 13, e0368021. [Google Scholar] [CrossRef]
- Kerner, M.J.; Naylor, D.J.; Ishihama, Y.; Maier, T.; Chang, H.C.; Stines, A.P.; Georgopoulos, C.; Frishman, D.; Hayer-Hartl, M.; Mann, M.; et al. Proteome-wide analysis of chaperonin-dependent protein folding in Escherichia coli. Cell 2005, 122, 209–220. [Google Scholar] [CrossRef]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G. DnaK functions as a central hub in the E. coli chaperone network. Cell Rep. 2012, 1, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Kitagawa, M.; Ara, T.; Arifuzzaman, M.; Ioka-Nakamichi, T.; Inamoto, E.; Toyonaga, H.; Mori, H. Complete set of ORF clones of Escherichia coli ASKA library (A Complete Set of E. coli K-12 ORF Archive): Unique resources for biological research. DNA Res. 2005, 12, 291–299. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Con-struction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006-0008. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Haslbeck, M.; Buchner, J. Assays to characterize molecular chaperone function in vitro. Methods Mol. Biol. 2015, 1292, 39–51. [Google Scholar]
- Tomoyasu, T.; Mogk, A.; Langen, H.; Goloubinoff, P.; Bukau, B. Genetic dissection of the roles of chaperones and proteases in protein folding and degradation in the Escherichia coli cytosol. Mol. Microbiol. 2001, 40, 397–413. [Google Scholar] [CrossRef]
- Brauer, M.; Hotop, S.K.; Wurster, M.; Herrmann, J.; Miethke, M.; Schlüter, R.; Dittmann, S.; Zühlke, D.; Brönstrup, M.; Lalk, M.; et al. Clostridioides difficile modifies its aromatic compound metabolism in response to amidochelocardin-induced membrane stress. mSphere 2022, 7, e0030222. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Zaiontz, C. Real Statistics Resource Pack Software (Release 8.8). Copyright (2013–2021). Available online: www.real-statistics.com (accessed on 18 May 2023).
- Aguilan, J.T.; Kulej, K.; Sidoli, S. Guide for protein fold change and p-value calculation for non-experts in proteomics. Mol. Omics 2020, 16, 573–582. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.r-project.org/ (accessed on 18 May 2023).
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: https://www.rstudio.com/ (accessed on 18 May 2023).
- Ramakrishnan, R.; Houben, B.; Rousseau, F.; Schymkowitz, J. Differential proteostatic regulation of insoluble and abundant proteins. Bioinformatics 2019, 35, 4098–4107. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Uniprot ID | Protein | Gene | Log2 Fold Change | p-Value |
|---|---|---|---|---|
| C. Energy production and conversion | ||||
| P36683 | bifunctional aconitate hydratase AcnB | acnB | 6.77 | 0.015 |
| P77252 | putative lactate utilization oxidoreductase YkgE | ykgE | 4.59 | 0.021 |
| P0AFG3 | subunit of E1(0) component of 2-oxoglutarate dehydrogenase | sucA | 1.08 | 0.034 |
| P0AFG6 | dihydrolipoyltranssuccinylase | sucB | 6.63 | 0.045 |
| P09373 | pyruvate formate-lyase | pflB | 7.94 | 0.000 |
| P0AC33 | fumarase A | fumA | 9.99 | 0.002 |
| P0A9C0 | anaerobic glycerol-3-phosphate dehydrogenase subunit A | glpA | 2.34 | 0.037 |
| P33602 | NADH:quinone oxidoreductase subunit G | nuoG | 4.45 | 0.049 |
| P0AFC7 | NADH:quinone oxidoreductase subunit B | nuoB | 3.70 | 0.013 |
| P61889 | malate dehydrogenase | mdh | 8.27 | 0.003 |
| P22259 | phosphoenolpyruvate carboxykinase (ATP) | pckA | 6.48 | 0.001 |
| P0ABB4 | ATP synthase F1 complex subunit beta | atpD | 7.32 | 0.004 |
| P0ABA6 | ATP synthase F1 complex subunit gamma | atpG | 5.33 | 0.003 |
| P0ABA4 | ATP synthase F1 complex subunit delta | atpH | 4.71 | 0.027 |
| P0A9G6 | isocitrate lyase | aceA | 7.77 | 0.038 |
| P00363 | fumarate reductase flavoprotein subunit | frdA | 5.29 | 0.045 |
| E. Amino acid transport and metabolism | ||||
| P00968 | carbamoyl-phosphate synthetase large subunit | carB | 2.63 | 0.003 |
| P30750 | L-methionine/D-methionine ABC transporter ATP binding subunit | metN | 4.13 | 0.049 |
| P15288 | peptidase D | pepD | 3.60 | 0.027 |
| P00509 | aspartate aminotransferase | aspC | 5.12 | 0.012 |
| P04825 | aminopeptidase N | pepN | 6.49 | 0.006 |
| P75914 | zinc-binding phosphatase YcdX | ycdX | 4.92 | 0.040 |
| P16095 | L-serine deaminase I | sdaA | 7.14 | 0.003 |
| P0A9S3 | galactitol-1-phosphate 5-dehydrogenase | gatD | 5.87 | 0.026 |
| P07913 | threonine dehydrogenase | tdh | 7.38 | 0.004 |
| P08142 | acetohydroxy acid synthase I subunit IlvB | ilvB | 5.96 | 0.004 |
| P00963 | asparagine synthetase A | asnA | 7.40 | 0.026 |
| P0AB80 | branched-chain-amino-acid aminotransferase | ilvE | 3.91 | 0.043 |
| F. Nucleotide transport and metabolism | ||||
| P22333 | adenosine deaminase | add | 5.63 | 0.019 |
| P0A8F0 | uracil phosphoribosyltransferase | upp | 7.77 | 0.024 |
| P0A763 | nucleoside diphosphate kinase | ndk | 5.03 | 0.004 |
| P0A7E5 | CTP synthetase | pyrG | 1.65 | 0.017 |
| G. Carbohydrate transport and metabolism | ||||
| P69797 | mannose-specific PTS enzyme IIAB component | manX | 7.37 | 0.000 |
| P37188 | galactitol-specific PTS enzyme IIB component | gatB | 9.14 | 0.002 |
| P0C8J8 | putative tagatose-1.6-bisphosphate aldolase 2 chaperone | gatZ | 9.20 | 0.003 |
| P0C8J6 | tagatose-1.6-bisphosphate aldolase 2 | gatY | 9.59 | 0.001 |
| P39829 | galactarate dehydratase GarD | garD | 4.30 | 0.032 |
| P04983 | ribose ABC transporter ATP binding subunit | rbsA | 6.05 | 0.018 |
| P0A9C9 | fructose-1.6-bisphosphatase 2 | glpX | 4.71 | 0.010 |
| P36672 | trehalose-specific PTS enzyme IIBC component | treB | 7.67 | 0.004 |
| P0A6K6 | phosphopentomutase | deoB | 5.02 | 0.027 |
| H. Coenzyme transport and metabolism | ||||
| P76085 | phenylacetate-CoA ligase | paaK | 5.49 | 0.049 |
| P0A8Y1 | pyrimidine 5-nucleotidase YjjG | yjjG | 3.95 | 0.017 |
| I. Lipid transport and metabolism | ||||
| P0AEK2 | 3-oxoacyl-[acyl-carrier-protein] reductase FabG | fabG | 7.02 | 0.015 |
| P0AAI5 | 3-oxoacyl-[acyl carrier protein] synthase 2 | fabF | 5.43 | 0.011 |
| P0AEK4 | enoyl-[acyl-carrier-protein] reductase | fabI | 7.56 | 0.048 |
| P0A9Q5 | acetyl-CoA carboxyltransferase subunit beta | accD | 4.80 | 0.038 |
| P37440 | oxidoreductase UcpA | ucpA | 1.55 | 0.039 |
| P. Inorganic ion transport and metabolism | ||||
| P13036 | ferric citrate outer membrane transporter | fecA | 4.28 | 0.009 |
| J. Translation. ribosomal structure and biogenesis | ||||
| P0A8I8 | 23S rRNA m(3)psi1915 methyltransferase | rlmH | 4.88 | 0.014 |
| P0AEI1 | isopentenyl-adenosine A37 tRNA methylthiolase | miaB | 7.01 | 0.020 |
| P0AEI4 | ribosomal protein S12 methylthiotransferase RimO | rimO | 7.88 | 0.006 |
| P75838 | ribosomal protein S12 methylthiotransferase accessory factor YcaO | ycaO | 3.38 | 0.047 |
| P0AG67 | 30S ribosomal subunit protein S1 | rpsA | 5.55 | 0.027 |
| P0ABU2 | redox-responsive ATPase YchF | ychF | 7.45 | 0.007 |
| P0A7I0 | peptide chain release factor RF1 | prfA | 4.40 | 0.006 |
| P0A7L3 | 50S ribosomal subunit protein L20 | rplT | 6.56 | 0.008 |
| P39199 | ribosomal protein L3 N(5)-glutamine methyltransferase | prmB | 2.82 | 0.024 |
| P0A6P5 | 50S ribosomal subunit stability factor | der | 2.50 | 0.027 |
| P0A7K6 | 50S ribosomal subunit protein L19 | rplS | 4.46 | 0.049 |
| P00957 | alanine--tRNA ligase/DNA-binding transcriptional repressor | alaS | 3.31 | 0.031 |
| P07012 | peptide chain release factor RF2 | prfB | 7.24 | 0.005 |
| P05055 | polynucleotide phosphorylase | pnp | 2.95 | 0.036 |
| P0A705 | translation initiation factor IF-2beta | infB | 8.08 | 0.003 |
| P0C0R7 | 23S rRNA 2-O-ribose U2552 methyltransferase | rlmE | 4.16 | 0.037 |
| P0A7X3 | 30S ribosomal subunit protein S9 | rpsI | 8.27 | 0.036 |
| P0A6K3 | peptide deformylase | def | 4.92 | 0.027 |
| P0AG44 | 50S ribosomal subunit protein L17 | rplQ | 5.86 | 0.044 |
| P0A7S9 | 30S ribosomal subunit protein S13 | rpsM | 6.68 | 0.028 |
| P0AG55 | 50S ribosomal subunit protein L6 | rplF | 3.05 | 0.049 |
| P0A7W7 | 30S ribosomal subunit protein S8 | rpsH | 4.59 | 0.016 |
| P62399 | 50S ribosomal subunit protein L5 | rplE | 3.92 | 0.010 |
| P0A7V3 | 30S ribosomal subunit protein S3 | rpsC | 9.26 | 0.021 |
| P61175 | 50S ribosomal subunit protein L22 | rplV | 7.24 | 0.024 |
| P0A7U3 | 30S ribosomal subunit protein S19 | rpsS | 7.63 | 0.016 |
| P60422 | 50S ribosomal subunit protein L2 | rplB | 8.76 | 0.006 |
| P0ADZ0 | 50S ribosomal subunit protein L23 | rplW | 6.85 | 0.021 |
| P60438 | 50S ribosomal subunit protein L3 | rplC | 6.31 | 0.002 |
| P0A6M8 | elongation factor G | fusA | 6.81 | 0.024 |
| P0A6U3 | 5-carboxymethylaminomethyluridine-tRNA synthase subunit MnmG | mnmG | 5.09 | 0.027 |
| P0CE48 | translation elongation factor Tu 2 | tufB | 7.05 | 0.017 |
| P0A7J7 | 50S ribosomal subunit protein L11 | rplK | 9.30 | 0.024 |
| P0A7L0 | 50S ribosomal subunit protein L1 | rplA | 3.64 | 0.006 |
| K. Transcription | ||||
| P60240 | RNA polymerase-binding ATPase and RNAP recycling factor | rapA | 6.78 | 0.000 |
| P0A972 | transcription antiterminator and regulator of RNA stability CspE | cspE | 4.78 | 0.027 |
| P0A9F3 | DNA-binding transcriptional dual regulator CysB | cysB | 3.15 | 0.039 |
| P07604 | DNA-binding transcriptional dual regulator TyrR | tyrR | 4.31 | 0.016 |
| P0ACM2 | DNA-binding transcriptional repressor RspR | rspR | 3.77 | 0.014 |
| P31802 | DNA-binding transcriptional dual regulator NarP | narP | 4.35 | 0.015 |
| P0AA16 | DNA-binding transcriptional dual regulator OmpR | ompR | 1.62 | 0.007 |
| P46837 | putative RNA-binding protein YhgF | yhgF | 6.70 | 0.000 |
| P06993 | DNA-binding transcriptional activator MalT | malT | 6.59 | 0.006 |
| P0AFG0 | transcription termination/antitermination factor NusG | nusG | 5.00 | 0.039 |
| P0A8V2 | RNA polymerase subunit beta | rpoB | 6.38 | 0.043 |
| L. Replication. recombination and repair | ||||
| P0A812 | Holliday junction branch migration complex subunit RuvB | ruvB | 5.03 | 0.012 |
| D. Cell cycle control. cell division. chromosome partitioning | ||||
| P0AEZ3 | Z-ring positioning protein MinD | minD | 8.03 | 0.009 |
| M. Cell wall/membrane/envelope biogenesis | ||||
| P17952 | UDP-N-acetylmuramate--L-alanine ligase | murC | 1.90 | 0.021 |
| P02931 | outer membrane porin F | ompF | 6.44 | 0.020 |
| P0A910 | outer membrane protein A | ompA | 4.04 | 0.020 |
| P0AEP3 | UTP--glucose-1-phosphate uridylyltransferase | galU | 8.19 | 0.000 |
| P69776 | murein lipoprotein | lpp | 6.21 | 0.040 |
| P37751 | putative glycosyltransferase WbbK | wbbK | 5.64 | 0.000 |
| P37749 | beta-1.6-galactofuranosyltransferase WbbI | wbbI | 5.10 | 0.023 |
| P37747 | UDP-galactopyranose mutase | glf | 6.24 | 0.003 |
| P06996 | outer membrane porin C | ompC | 7.27 | 0.047 |
| P02930 | outer membrane channel TolC | tolC | 2.90 | 0.023 |
| P0A749 | UDP-N-acetylglucosamine 1-carboxyvinyltransferase | murA | 2.32 | 0.002 |
| P0A9V1 | lipopolysaccharide transport system ATP binding protein LptB | lptB | 4.70 | 0.023 |
| P25714 | membrane protein insertase YidC | yidC | 4.86 | 0.014 |
| P17169 | L-glutamine-D-fructose-6-phosphate aminotransferase | glmS | 4.88 | 0.018 |
| P22634 | glutamate racemase | murI | 4.40 | 0.025 |
| O. Posttranslational modification. protein turnover. chaperones | ||||
| P0ABZ6 | chaperone SurA | surA | 5.40 | 0.032 |
| P0ACA7 | glutathione S-transferase GstB | gstB | 3.75 | 0.012 |
| P63284 | chaperone protein ClpB | clpB | 3.70 | 0.042 |
| P0AAI3 | ATP-dependent zinc metalloprotease FtsH | ftsH | 2.44 | 0.017 |
| P39099 | periplasmic serine endoprotease | degQ | 3.77 | 0.002 |
| P0A6F5 | chaperonin GroEL | groEL | 2.27 | 0.018 |
| P0ABC3 | regulator of FtsH protease | hflC | 2.21 | 0.038 |
| T. Signal transduction mechanisms | ||||
| P0A964 | chemotaxis protein CheW | cheW | 5.91 | 0.011 |
| V. Defense mechanisms | ||||
| P0AE08 | alkyl hydroperoxide reductase. AhpC component | ahpC | 6.43 | 0.033 |
| P0ABT2 | DNA protection during starvation protein | dps | 1.86 | 0.010 |
| P06610 | thioredoxin/glutathione peroxidase BtuE | btuE | 4.96 | 0.026 |
| Poorly Characterized | ||||
| R. General function prediction only | ||||
| P24203 | P-loop guanosine triphosphatase YjiA | yjiA | 5.61 | 0.007 |
| Non categories | ||||
| P0DTT0 | 50S ribosomal subunit assembly factor BipA | bipA | 2.76 | 0.038 |
| Gene | GenBank Code | Primer Name | Sequence (5′-3′) | Amplicon (bp) |
|---|---|---|---|---|
| Pet21b cloning | ||||
| tfp2 | AKS23718 | pET21b_F_TFP2 | TTCCATATGGTGGAAGTAAATGCTCCG | 315 |
| pET21b_R_TFP2 | TTTAAGCTTGGACTTGAGAAGAGAGTCAA | |||
| RT-qPCR | ||||
| proP | AKS24793 | Prot_RT2_F | AGACCCTATCTTCACTTCCC | 105 |
| Prot_RT2_R | AATCGTCGCTCTCTTGTAATC | |||
| groES | AKS23720 | GroES_RT2_F | CAGAAAGGCAAGATCGAAAG | 96 |
| GroES_RT2_R | TGTGATCTTGGAACCAGAATA | |||
| groEL | AKS23719 | GroEL_RT2_F | CGTGGCTATATCTCTCCCTAT | 101 |
| GroEL_RT2_R | ATGCTGCTGACTTTCTTCTC | |||
| hyp | * | Hyp_RT2_F | TTAATTCTCTTCTGGCCTACA | 108 |
| Hyp_RT2-R | TTCTTGAATTCTTTGTATCGGA | |||
| tfp2 | AKS23718 | pTfp2-F | CTGGTAATGGTTGATTTCTGGG | 104 |
| pTfp2-R | GGAATGCCCATGACCTGATAT | |||
| recN | AKS23717 | RecN_RT_F | AGCAGCAAGTATGGTGTATG | 97 |
| RecN_RT_R | TACTTTCAGACGGTCCAAATC | |||
| rrsB | CP012147 | pRrsB-F | TACAAGCTTCCGCTCCTG | 288 |
| pRrsB-R | CCGGGCAAAAGTGGTTTACA | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Villagrán, C.; Acevedo-Arbunic, J.; Härtig, E.; Sievers, S.; Zühlke, D.; Issotta, F.; Mascayano, C.; Jahn, D.; Jahn, M.; Levicán, G. The Thioredoxin Fold Protein (TFP2) from Extreme Acidophilic Leptospirillum sp. CF-1 Is a Chaperedoxin-like Protein That Prevents the Aggregation of Proteins under Oxidative Stress. Int. J. Mol. Sci. 2024, 25, 6905. https://doi.org/10.3390/ijms25136905
Muñoz-Villagrán C, Acevedo-Arbunic J, Härtig E, Sievers S, Zühlke D, Issotta F, Mascayano C, Jahn D, Jahn M, Levicán G. The Thioredoxin Fold Protein (TFP2) from Extreme Acidophilic Leptospirillum sp. CF-1 Is a Chaperedoxin-like Protein That Prevents the Aggregation of Proteins under Oxidative Stress. International Journal of Molecular Sciences. 2024; 25(13):6905. https://doi.org/10.3390/ijms25136905
Chicago/Turabian StyleMuñoz-Villagrán, Claudia, Javiera Acevedo-Arbunic, Elisabeth Härtig, Susanne Sievers, Daniela Zühlke, Francisco Issotta, Carolina Mascayano, Dieter Jahn, Martina Jahn, and Gloria Levicán. 2024. "The Thioredoxin Fold Protein (TFP2) from Extreme Acidophilic Leptospirillum sp. CF-1 Is a Chaperedoxin-like Protein That Prevents the Aggregation of Proteins under Oxidative Stress" International Journal of Molecular Sciences 25, no. 13: 6905. https://doi.org/10.3390/ijms25136905
APA StyleMuñoz-Villagrán, C., Acevedo-Arbunic, J., Härtig, E., Sievers, S., Zühlke, D., Issotta, F., Mascayano, C., Jahn, D., Jahn, M., & Levicán, G. (2024). The Thioredoxin Fold Protein (TFP2) from Extreme Acidophilic Leptospirillum sp. CF-1 Is a Chaperedoxin-like Protein That Prevents the Aggregation of Proteins under Oxidative Stress. International Journal of Molecular Sciences, 25(13), 6905. https://doi.org/10.3390/ijms25136905