
Genome-Wide Identification of the Sulfate Transporters Gene Family in Blueberry (Vaccinium spp.) and Its Response to Ericoid Mycorrhizal Fungi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Identification of SULTR Genes in the Blueberry Genome
2.2. Phylogenetic Analysis of VcSULTRs
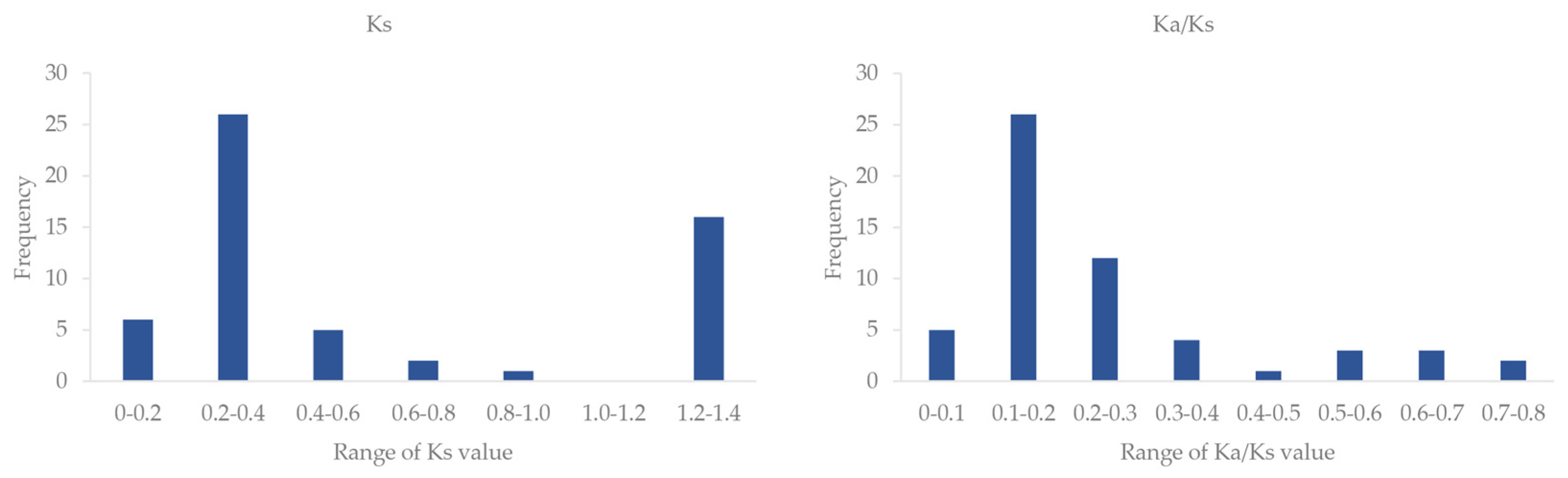
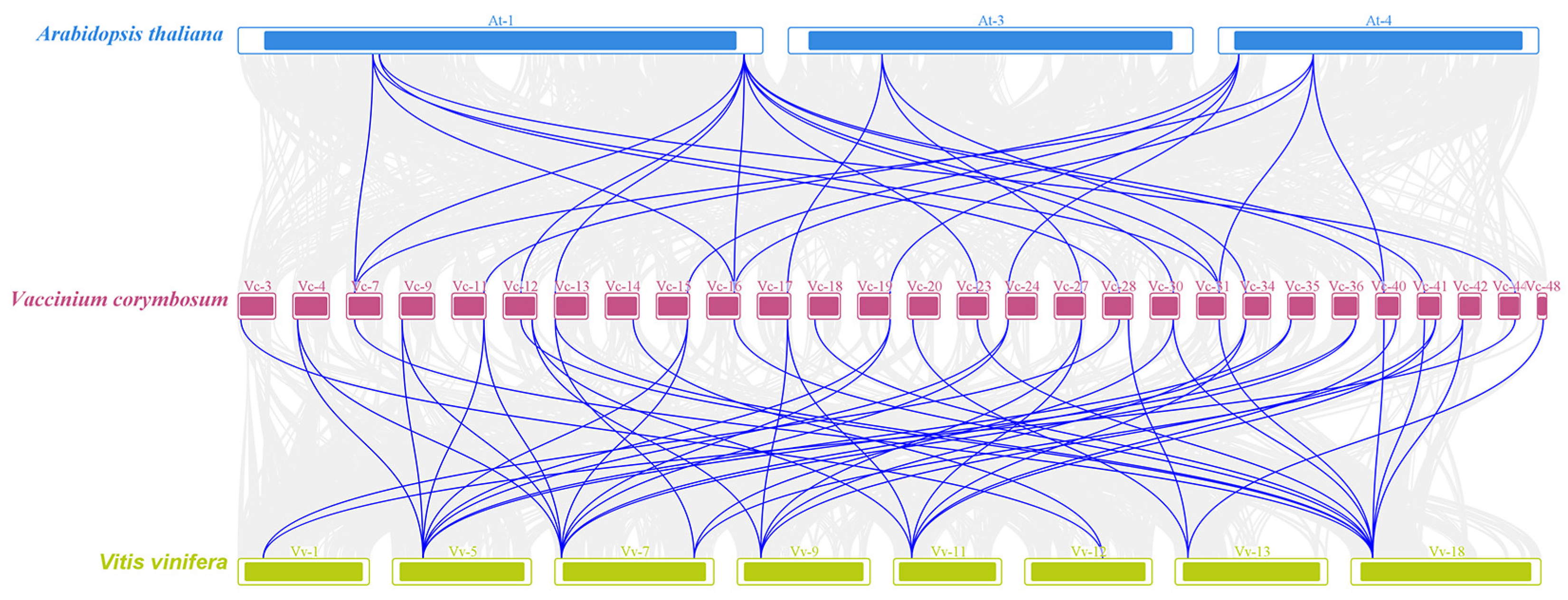
2.3. Chromosomal Location of VcSULTRs and Collinearity Analysis
2.4. Conserved Motifs and Gene Structure of VcSULTRs
2.5. Identification of Cis-Regulatory Elements in VcSULTR Promoter Regions
2.6. Expression Patterns of SULTR Genes in Blueberry
2.7. Expression Analysis of VcSULTR Genes in Response to ERMF Inoculation and Sulfate Treatments
2.8. Subcellular Localization of VcSULTR2;1c
2.9. Expression Analysis of VcSULTR2;1c and Its Involvement in Sulfate Absorption
2.10. Virus-Induced Gene Silencing of VcSULTR2;1c
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Identification of SULTR Gene Family Members in Blueberry
4.3. Gene Information and Phylogenetic Relationships
4.4. Chromosomal Location of Genes and Collinearity Analysis
4.5. Analysis of Domain Motifs and Gene Structures
4.6. Analysis of VcSULTR Gene Promoter Regions
4.7. Expression Pattern Analysis of VcSULTRs
4.8. Cloning of VcSULTR2;1c and Subcellular Localization
4.9. Prokaryotic Expression of VcSULTR2;1c
4.10. Transient Transformation of Blueberry Root
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Günther, C.S.; Dare, A.P.; McGhie, T.K.; Deng, C.; Lafferty, D.J.; Plunkett, B.J.; Grierson, E.R.P.; Turner, J.L.; Jaakola, L.; Albert, N.W.; et al. Spatiotemporal modulation of flavonoid metabolism in blueberries. Front. Plant Sci. 2020, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Cassidy, A.; Howard, L.R.; Krikorian, R.; Stull, A.J.; Tremblay, F.; Zamora-Ros, R. Recent research on the health benefits of blueberries and their anthocyanins. Adv. Nutr. 2020, 11, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Retamales, J.B.; Hancock, J.F. Blueberry taxonomy and breeding. In Blueberries, 2nd ed.; CABI: Wallington, NJ, USA, 2018; pp. 18–60. [Google Scholar]
- Edger, P.P.; Iorizzo, M.; Bassil, N.V.; Benevenuto, J.; Ferrão, L.F.V.; Giongo, L.; Hummer, K.; Lawas, L.M.F.; Leisner, C.P.; Li, C.; et al. There and back again; historical perspective and future directions for Vaccinium breeding and research studies. Hortic. Res. 2022, 9, uhac083. [Google Scholar] [CrossRef] [PubMed]
- Pescie, M.A.; Fradkin, M.; Lavado, R.S.; Chiocchio, V.M. Endophytic fungi in blueberry cultivars, in three production areas of Argentina. Physiol. Mol. Plant Pathol. 2021, 115, 101662. [Google Scholar] [CrossRef]
- Caspersen, S.; Svensson, B.; Håkansson, T.; Winter, C.; Khalil, S.; Asp, H. Blueberry—Soil interactions from an organic perspective. Sci. Hortic. 2016, 208, 78–91. [Google Scholar] [CrossRef]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 2023, 18, 2030082. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gao, Y.; Yang, A. Sulfur homeostasis in plants. Int. J. Mol. Sci. 2020, 21, 8926. [Google Scholar] [CrossRef] [PubMed]
- Narayan, O.P.; Verma, N.; Jogawat, A.; Dua, M.; Johri, A.K. Sulfur transfer from the endophytic fungus Serendipita indica improves maize growth and requires the sulfate transporter SiSulT. Plant Cell 2021, 33, 1268–1285. [Google Scholar] [CrossRef] [PubMed]
- Davidian, J.C.; Kopriva, S. Regulation of sulfate uptake and assimilation--the same or not the same? Mol. Plant 2010, 3, 314–325. [Google Scholar] [CrossRef]
- Casieri, L.; Ait Lahmidi, N.; Doidy, J.; Veneault-Fourrey, C.; Migeon, A.; Bonneau, L.; Courty, P.E.; Garcia, K.; Charbonnier, M.; Delteil, A.; et al. Biotrophic transportome in mutualistic plant-fungal interactions. Mycorrhiza 2013, 23, 597–625. [Google Scholar] [CrossRef]
- Garcia, K.; Doidy, J.; Zimmermann, S.D.; Wipf, D.; Courty, P.E. Take a trip through the plant and fungal transportome of mycorrhiza. Trends Plant Sci. 2016, 21, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Declerck, S.; Feng, G.; Zhang, L. Hyphosphere interactions between Rhizophagus irregularis and Rahnella aquatilis promote carbon-phosphorus exchange at the peri-arbuscular space in Medicago truncatula. Environ. Microbiol. 2023, 25, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, K.; Zhou, M. Structure and function of an Arabidopsis thaliana sulfate transporter. Nat. Commun. 2021, 12, 4455. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Geng, J.; Du, Y.; Li, S.; Yuan, X.; Zhu, J.; Zhou, Z.; Wang, Q.; Du, J. The common bean (Phaseolus vulgaris) SULTR gene family: Genome-wide identification, phylogeny, evolutionary expansion and expression patterns. Biotechnol. Biotechnol. Equip. 2022, 36, 724–736. [Google Scholar] [CrossRef]
- Xun, M.; Song, J.; Shi, J.; Li, J.; Shi, Y.; Yan, J.; Zhang, W.; Yang, H. Genome-wide identification of sultr genes in Malus domestica and low sulfur-induced MhSultr3;1a to increase cysteine-improving growth. Front. Plant Sci. 2021, 12, 74842. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hao, X.; Zhang, J.; Wang, L.; Wang, Y.; Li, N.; Guo, L.; Ren, H.; Zeng, J. Genome-wide identification of SULTR genes in tea plant and analysis of their expression in response to sulfur and selenium. Protoplasma 2022, 259, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Puresmaeli, F.; Heidari, P.; Lawson, S. Insights into the sulfate transporter gene family and its expression patterns in durum wheat seedlings under salinity. Genes 2023, 14, 333. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Long, W.; Hu, H.; Liang, T.; Luo, X.; Hu, Z.; Zhu, R.; Wu, X. Genome-wide identification and expansion patterns of SULTR gene family in gramineae crops and their expression profiles under abiotic stress in Oryza sativa. Genes 2021, 12, 634. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Filiz, E.; Aydın, M.; Koc, I. Genome-wide identification and expression analysis of sulphate transporter (SULTR) genes under sulfur deficiency in Brachypodium distachyon. J. Plant Biochem. Biot. 2017, 26, 263–273. [Google Scholar] [CrossRef]
- Takahashi, H. Sulfate transport systems in plants: Functional diversity and molecular mechanisms underlying regulatory coordination. J. Exp. Bot. 2019, 70, 4075–4087. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, X.; Zuo, L.; Wang, H.; Yu, D. Identification and functional characterization of the sulfate transporter gene GmSULTR1;2b in soybean. BMC Genom. 2016, 17, 373. [Google Scholar] [CrossRef]
- Yang, Z.; Hui, S.; Lv, Y.; Zhang, M.; Chen, D.; Tian, J.; Zhang, H.; Liu, H.; Cao, J.; Xie, W.; et al. miR395-regulated sulfate metabolism exploits pathogen sensitivity to sulfate to boost immunity in rice. Mol. Plant 2022, 15, 671–688. [Google Scholar] [CrossRef] [PubMed]
- Heidari, P.; Hasanzadeh, S.; Faraji, S.; Ercisli, S.; Mora-Poblete, F. Genome-wide characterization of the sulfate transporter gene family in oilseed crops: Camelina sativa and Brassica napus. Plants 2023, 12, 628. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, P.X.; Miao, Z.Q.; Qi, G.F.; Wang, Z.; Yuan, Y.; Ahmad, N.; Cao, M.J.; Hell, R.; Wirtz, M.; et al. SULTR3s function in chloroplast sulfate uptake and affect ABA biosynthesis and the stress response. Plant Physiol. 2019, 180, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, R.; Koc, I.; Ozyigit, I.I.; Sen, U.; Uras, M.E.; Anjum, N.A.; Pereira, E.; Filiz, E. Genome-wide identification and expression analysis of sulfate transporter (SULTR) genes in potato (Solanum tuberosum L.). Planta 2016, 244, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Akbudak, M.A.; Filiz, E.; Kontbay, K. Genome-wide identification and cadmium induced expression profiling of sulfate transporter (SULTR) genes in sorghum (Sorghum bicolor L.). Biometals 2018, 31, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Wang, M.; Xia, Z. The SULTR gene family in maize (Zea mays L.): Gene cloning and expression analyses under sulfate starvation and abiotic stress. J. Plant Physiol. 2018, 220, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Dürr, J.; Bücking, H.; Mult, S.; Wildhagen, H.; Palme, K.; Rennenberg, H.; Ditengou, F.; Herschbach, C. Seasonal and cell type specific expression of sulfate transporters in the phloem of Populus reveals tree specific characteristics for SO42− storage and mobilization. Plant Mol. Biol. 2010, 72, 499–517. [Google Scholar] [CrossRef]
- Casieri, L.; Gallardo, K.; Wipf, D. Transcriptional response of Medicago truncatula sulphate transporters to arbuscular mycorrhizal symbiosis with and without sulphur stress. Planta 2012, 235, 1431–1447. [Google Scholar] [CrossRef]
- Lin, L.; Chen, Q.; Yuan, K.; Xing, C.; Qiao, Q.; Huang, X.; Zhang, S. PbrATL18, an E3 ubiquitin ligase identified by genome-wide identification, is a positive factor in pear resistance to drought and Colletotrichum fructicola infection. Hortic. Plant J. 2023, 10, 698. [Google Scholar] [CrossRef]
- Navarro Gallón, S.M.; Elejalde-Palmett, C.; Daudu, D.; Liesecke, F.; Jullien, F.; Papon, N.; Dugé de Bernonville, T.; Courdavault, V.; Lanoue, A.; Oudin, A.; et al. Virus-induced gene silencing of the two squalene synthase isoforms of apple tree (Malus × domestica L.) negatively impacts phytosterol biosynthesis, plastid pigmentation and leaf growth. Planta 2017, 246, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wei, Y.; Liang, C.; Guo, J.; Niu, T.; Zhang, P.; Wen, P. VvANR silencing promotes expression of VvANS and accumulation of anthocyanin in grape berries. Protoplasma 2022, 259, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Pei, J.; Li, Y.; Sun, H. Genome-wide identification and characterization of COMT gene family during the development of blueberry fruit. BMC Plant Biol. 2021, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Suzuki, K.; Ohtsu, I. Current understanding of sulfur assimilation metabolism to biosynthesize L-cysteine and recent progress of its fermentative overproduction in microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 8203–8211. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Hu, J.; Xu, G.; Song, H.; Zhang, L.; Cao, Y. An efficient transformation system for fast production of VcCHS transgenic blueberry callus and its expressional analysis. Plants 2023, 12, 2905. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Vancov, T.; Si, H.; Yang, W.; Tong, K.; Chen, W.; Fang, Y. Isolation and characterization of endomycorrhizal fungi associated with growth promotion of blueberry plants. J. Fungi 2021, 7, 584. [Google Scholar] [CrossRef] [PubMed]
- Sieh, D.; Watanabe, M.; Devers, E.A.; Brueckner, F.; Hoefgen, R.; Krajinski, F. The arbuscular mycorrhizal symbiosis influences sulfur starvation responses of Medicago truncatula. New Phytol. 2013, 197, 606–616. [Google Scholar] [CrossRef]
- Colle, M.; Leisner, C.P.; Wai, C.M.; Ou, S.; Bird, K.A.; Wang, J.; Wisecaver, J.H.; Yocca, A.E.; Alger, E.I.; Tang, H.; et al. Haplotype-phased genome and evolution of phytonutrient pathways of tetraploid blueberry. Gigascience 2019, 8, giz012. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.S.; Eddy, S.R.; Portugaly, E. Hidden Markov model speed heuristic and iterative HMM search procedure. BMC Bioinform. 2010, 11, 431. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, C.; Takamiya-Wik, M.; Schmelzer, E.; Hahlbrock, K.; Somssich, I.E. A novel regulatory element involved in rapid activation of parsley ELI7 gene family members by fungal elicitor or pathogen infection. Mol. Plant Pathol. 2000, 1, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Gu, M.; Sun, S.; Zhu, L.; Hong, S.; Xu, G. Identification of two conserved cis-acting elements, MYCS and P1BS, involved in the regulation of mycorrhiza-activated phosphate transporters in eudicot species. New Phytol. 2011, 189, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zan, X.L.; Wu, X.F.; Yao, L.; Chen, Y.L.; Jia, S.W.; Zhao, K.J. Identification of fungus-responsive cis-acting element in the promoter of Brassica juncea chitinase gene, BjCHI1. Plant Sci. 2014, 215–216, 190–198. [Google Scholar] [CrossRef]
- Dong, M.; Wang, B.; Tian, Y.; Chen, L.; Li, Y.; Sun, H. Diversity of fungal assemblages in rhizosphere and endosphere of blueberry (Vaccinium spp.) under field conditions revealed by culturing and culture-independent molecular methods. Can. J. Microbiol. 2022, 68, 622–632. [Google Scholar] [CrossRef]
- Deng, Y.; Li, Y.; Sun, H. Selection of reference genes for RT-qPCR normalization in blueberry (Vaccinium corymbosum × angustifolium) under various abiotic stresses. FEBS Open Bio. 2020, 10, 1418–1435. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Liu, H.; Kang, L.; Geng, J.; Gai, Y.; Ding, Y.; Sun, H.; Li, Y. Identification and expression analysis of MATE genes involved in flavonoid transport in blueberry plants. PLoS ONE 2015, 10, e0118578. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.M.; Lin, K.C.; Liau, W.S.; Chao, Y.Y.; Yang, L.H.; Chen, S.Y.; Lu, C.A.; Hong, C.Y. A set of GFP-based organelle marker lines combined with DsRed-based gateway vectors for subcellular localization study in rice (Oryza sativa L.). Plant Mol. Biol. 2016, 90, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Lv, W.; Jiang, H.; Chen, X.; Wang, X.; Wang, Y. Genome-wide identification of glutathione S-transferase gene family members in tea plant (Camellia sinensis) and their response to environmental stress. Int. J. Biol. Macromol. 2022, 205, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Mo, R.; Yang, S.; Zhang, Q.; Xu, L.; Luo, Z. Vacuum infiltration enhances the Agrobacterium-mediated transient transformation for gene functional analysis in persimmon (Diospyros kaki Thunb.). Sci. Hortic. 2019, 251, 174–180. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, M.; He, J.; Tang, X.; Liu, S.; Xing, J.; Chen, X.; Chen, L.; Li, Y.; Sun, H. Genome-Wide Identification of the Sulfate Transporters Gene Family in Blueberry (Vaccinium spp.) and Its Response to Ericoid Mycorrhizal Fungi. Int. J. Mol. Sci. 2024, 25, 6980. https://doi.org/10.3390/ijms25136980
Dong M, He J, Tang X, Liu S, Xing J, Chen X, Chen L, Li Y, Sun H. Genome-Wide Identification of the Sulfate Transporters Gene Family in Blueberry (Vaccinium spp.) and Its Response to Ericoid Mycorrhizal Fungi. International Journal of Molecular Sciences. 2024; 25(13):6980. https://doi.org/10.3390/ijms25136980
Chicago/Turabian StyleDong, Mei, Jiawei He, Xiaoxuan Tang, Siwen Liu, Jinjie Xing, Xuyang Chen, Li Chen, Yadong Li, and Haiyue Sun. 2024. "Genome-Wide Identification of the Sulfate Transporters Gene Family in Blueberry (Vaccinium spp.) and Its Response to Ericoid Mycorrhizal Fungi" International Journal of Molecular Sciences 25, no. 13: 6980. https://doi.org/10.3390/ijms25136980
APA StyleDong, M., He, J., Tang, X., Liu, S., Xing, J., Chen, X., Chen, L., Li, Y., & Sun, H. (2024). Genome-Wide Identification of the Sulfate Transporters Gene Family in Blueberry (Vaccinium spp.) and Its Response to Ericoid Mycorrhizal Fungi. International Journal of Molecular Sciences, 25(13), 6980. https://doi.org/10.3390/ijms25136980