
Anti-Persisters Activity of Lacticaseibacillus rhamnosus Culture Filtrates against Pseudomonas aeruginosa in Artificial Sputum Medium





Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
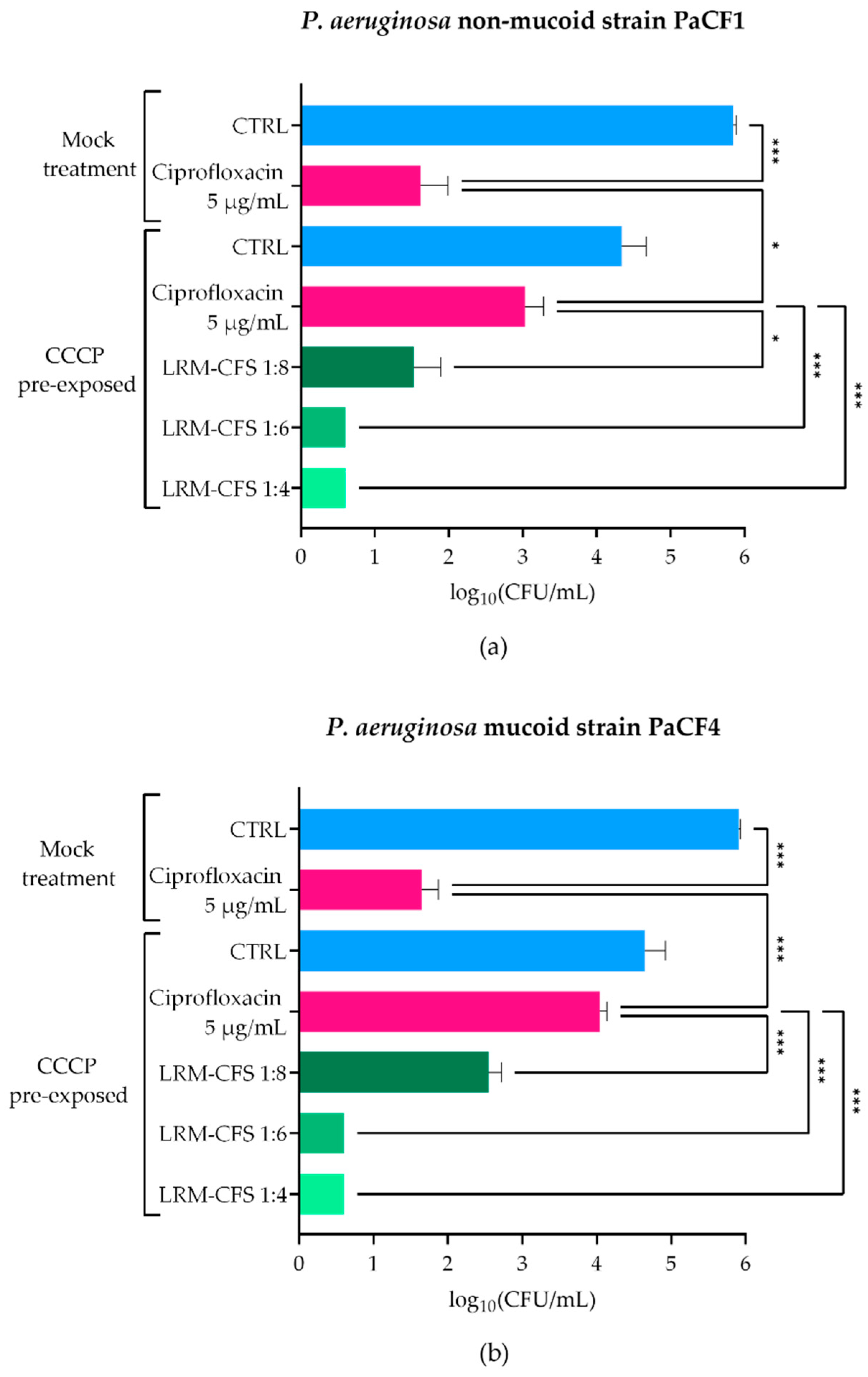
2.1. Bactericidal Activity of LRM-CFS in ASM against CCCP-Generated P. aeruginosa Persisters
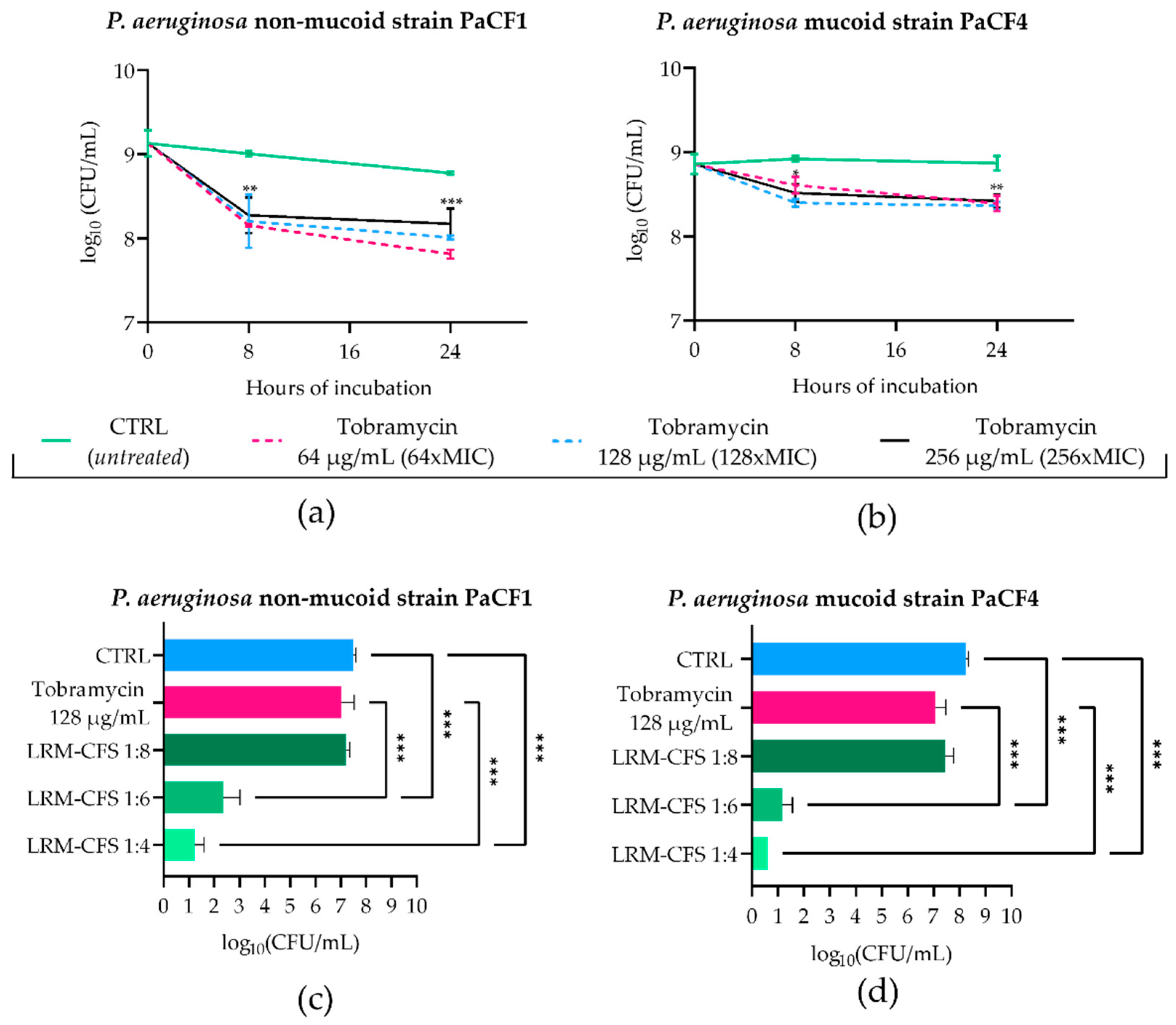
2.2. Bactericidal Activity of LRM-CFS in ASM against Tobramycin-Selected P. aeruginosa Persisters
2.3. Bactericidal Activity of LRM-CFS against P. aeruginosa Persisters from Biofilm
2.4. Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. L. rhamnosus Culture Filtrate (LRM-CFS) Preparation
4.3. Preparation of Artificial Sputum Medium (ASM)
4.4. Reagents
4.5. Determination of Minimum Inhibitory Concentrations (MICs)
4.6. CCCP Induction of P. aeruginosa Planktonic Persisters and Evaluation of LRM-CFS Activity
4.7. Tobramycin Selection of P. aeruginosa Planktonic Persisters in ASM and Evaluation of LRM-CFS Activity
4.8. Ciprofloxacin Selection of P. aeruginosa Biofilm Persisters in ASM and Evaluation of LRM-CFS Activity
4.9. Cytotoxicity
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turcios, N.L. Cystic Fibrosis Lung Disease: An Overview. Respir. Care 2020, 65, 233–251. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.; Weldon, S.; Elborn, S.; Downey, D.G.; Taggart, C. The Effect of CFTR Modulators on Airway Infection in Cystic Fibrosis. Int. J. Mol. Sci. 2022, 23, 3513. [Google Scholar] [CrossRef] [PubMed]
- Maselli, D.J.; Keyt, H.; Restrepo, M.I. Inhaled Antibiotic Therapy in Chronic Respiratory Diseases. Int. J. Mol. Sci. 2017, 18, 1062. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.R.; Burns, J.L.; Lory, S.; Lewis, K. Emergence of Pseudomonas aeruginosa Strains Producing High Levels of Persister Cells in Patients with Cystic Fibrosis. J. Bacteriol. 2010, 192, 6191–6199. [Google Scholar] [CrossRef] [PubMed]
- Cirilli, N.; Schiavoni, V.; Tagliabracci, V.; Gesuita, R.; Tiano, L.; Fabrizzi, B.; D’Antuono, A.; Peruzzi, A.; Cedraro, N.; Carle, F.; et al. Role of Viable but Non Culturable Cells in Patients with Cystic Fibrosis in the Era of Highly Effective Modulator Therapy. J. Cyst. Fibros. 2024; in press. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-T.; Lohia, G.K.; Chen, S.; Riquelme, S.A. Immunometabolic Regulation of Bacterial Infection, Biofilms, and Antibiotic Susceptibility. J. Innate Immun. 2024, 16, 143. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Billón, M.; Llambías-Cabot, A.E.; Jordana-Lluch, E.; Oliver, A.; Macià, M.D. Mechanisms of Antibiotic Resistance in Pseudomonas aeruginosa Biofilms. Biofilm 2023, 5, 100129. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of Bacterial Persistence during Stress and Antibiotic Exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef] [PubMed]
- Batoni, G.; Catelli, E.; Kaya, E.; Pompilio, A.; Bianchi, M.; Ghelardi, E.; Di Bonaventura, G.; Esin, S.; Maisetta, G. Antibacterial and Antibiofilm Effects of Lactobacilli Strains against Clinical Isolates of Pseudomonas aeruginosa under Conditions Relevant to Cystic Fibrosis. Antibiotics 2023, 12, 1158. [Google Scholar] [CrossRef] [PubMed]
- Batoni, G.; Kaya, E.; Catelli, E.; Quinti, S.; Botti, M.; De Carli, A.; Bianchi, M.; Maisetta, G.; Esin, S. Lactobacillus Probiotic Strains Differ in Their Ability to Adhere to Human Lung Epithelial Cells and to Prevent Adhesion of Clinical Isolates of Pseudomonas aeruginosa from Cystic Fibrosis Lung. Microorganisms 2023, 11, 1707. [Google Scholar] [CrossRef] [PubMed]
- Fangous, M.-S.; Lazzouni, I.; Alexandre, Y.; Gouriou, S.; Boisramé, S.; Vallet, S.; Le Bihan, J.; Ramel, S.; Héry-Arnaud, G.; Le Berre, R. Prevalence and Dynamics of Lactobacillus sp. in the Lower Respiratory Tract of Patients with Cystic Fibrosis. Res. Microbiol. 2018, 169, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Glieca, S.; Quarta, E.; Bottari, B.; Bancalari, E.; Monica, S.; Scaltriti, E.; Tambassi, M.; Flammini, L.; Bertoni, S.; Bianchera, A.; et al. Development of Inhalation Powders Containing Lactic Acid Bacteria with Antimicrobial Activity against Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2024, 63, 107001. [Google Scholar] [CrossRef] [PubMed]
- Trindade, L.M.; Torres, L.; Matos, I.D.; Miranda, V.C.; de Jesus, L.C.L.; Cavalcante, G.; de Souza Oliveira, J.J.; Cassali, G.D.; Mancha-Agresti, P.; de Carvalho Azevedo, V.A.; et al. Paraprobiotic Lacticaseibacillus rhamnosus Protects Intestinal Damage in an Experimental Murine Model of Mucositis. Probiotics Antimicrob. Proteins 2023, 15, 338–350. [Google Scholar] [CrossRef]
- Piqué, N.; Berlanga, M.; Miñana-Galbis, D. Health Benefits of Heat-Killed (Tyndallized) Probiotics: An Overview. Int. J. Mol. Sci. 2019, 20, 2534. [Google Scholar] [CrossRef] [PubMed]
- Batista, V.L.; De Jesus, L.C.L.; Tavares, L.M.; Barroso, F.L.A.; Fernandes, L.J.d.S.; Freitas, A.d.S.; Americo, M.F.; Drumond, M.M.; Mancha-Agresti, P.; Ferreira, E.; et al. Paraprobiotics and Postbiotics of Lactobacillus delbrueckii CIDCA 133 Mitigate 5-FU-Induced Intestinal Inflammation. Microorganisms 2022, 10, 1418. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Molaei, R.; Guimarães, J.T. A Review on Preparation and Chemical Analysis of Postbiotics from Lactic Acid Bacteria. Enzym. Microb. Technol. 2021, 143, 109722. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Kaya, E.; Lupetti, V.; Catelli, E.; Bianchi, M.; Maisetta, G.; Esin, S.; Di Bonaventura, G.; Batoni, G. Cell-Free Supernatants from Lactobacillus Strains Exert Antibacterial, Antibiofilm, and Antivirulence Activity against Pseudomonas aeruginosa from Cystic Fibrosis Patients. Microbes Infect. 2024, 26, 105301. [Google Scholar] [CrossRef] [PubMed]
- Kaya, E.; Bianchi, M.; Maisetta, G.; Esin, S.; Batoni, G. Strong Activity and No Resistance Induction Exerted by Cell-Free Supernatants from Lacticaseibacillus rhamnosus against Mono-Species and Dual-Species Biofilms of Wound Pathogens in In Vivo-like Conditions. Int. J. Mol. Sci. 2024, 25, 2087. [Google Scholar] [CrossRef] [PubMed]
- Grassi, L.; Di Luca, M.; Maisetta, G.; Rinaldi, A.C.; Esin, S.; Trampuz, A.; Batoni, G. Generation of Persister Cells of Pseudomonas aeruginosa and Staphylococcus aureus by Chemical Treatment and Evaluation of Their Susceptibility to Membrane-Targeting Agents. Front. Microbiol. 2017, 8, 1917. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Bicker, J.; Falcão, A.; Fortuna, A. Air-Liquid Interface (ALI) Impact on Different Respiratory Cell Cultures. Eur. J. Pharm. Biopharm. 2023, 184, 62–82. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent Bacterial Infections and Persister Cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.; Buchad, H.; Gajjar, D. Pseudomonas aeruginosa Persister Cell Formation upon Antibiotic Exposure in Planktonic and Biofilm State. Sci. Rep. 2022, 12, 16151. [Google Scholar] [CrossRef] [PubMed]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister Formation in Staphylococcus aureus Is Associated with ATP Depletion. Nat. Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef] [PubMed]
- Torshabi, M.; Bardouni, M.M.; Hashemi, A. Evaluation of Antioxidant and Antibacterial Effects of Lyophilized Cell-Free Probiotic Supernatants of Three Lactobacillus spp. and Their Cytocompatibility against Periodontal Ligament Stem Cells. Iran. J. Pharm. Res. 2023, 22, e136438. [Google Scholar] [CrossRef] [PubMed]
- Zanetta, P.; Squarzanti, D.F.; di Coste, A.; Amoruso, A.; Pane, M.; Azzimonti, B. Growth Conditions Influence Lactobacillus Cell-Free Supernatant Impact on Viability, Biofilm Formation, and Co-Aggregation of the Oral Periodontopathogens Fusobacterium nucleatum and Porphyromonas gingivalis. Biomedicines 2023, 11, 859. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.; Alexandre, K.; Etienne, M. Tolerance and Persistence of Pseudomonas aeruginosa in Biofilms Exposed to Antibiotics: Molecular Mechanisms, Antibiotic Strategies and Therapeutic Perspectives. Front. Microbiol. 2020, 11, 423. [Google Scholar] [CrossRef] [PubMed]
- Drumond, M.M.; Tapia-Costa, A.P.; Neumann, E.; Nunes, Á.C.; Barbosa, J.W.; Kassuha, D.E.; Mancha-Agresti, P. Cell-Free Supernatant of Probiotic Bacteria Exerted Antibiofilm and Antibacterial Activities against Pseudomonas aeruginosa: A Novel Biotic Therapy. Front. Pharmacol. 2023, 14, 1152588. [Google Scholar] [CrossRef] [PubMed]
- Brul, S.; Coote, P. Preservative Agents in Foods. Mode of Action and Microbial Resistance Mechanisms. Int. J. Food Microbiol. 1999, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Salmond, C.V.; Kroll, R.G.; Booth, I.R. The Effect of Food Preservatives on pH Homeostasis in Escherichia coli. J. Gen. Microbiol. 1984, 130, 2845–2850. [Google Scholar] [CrossRef] [PubMed]
- Roe, A.J.; McLaggan, D.; Davidson, I.; O’Byrne, C.; Booth, I.R. Perturbation of Anion Balance during Inhibition of Growth of Escherichia coli by Weak Acids. J. Bacteriol. 1998, 180, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Alakomi, H.L.; Skyttä, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I.M. Lactic Acid Permeabilizes Gram-Negative Bacteria by Disrupting the Outer Membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of Culture Media, pH and Temperature on Growth and Bacteriocin Production of Bacteriocinogenic Lactic Acid Bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.M.; Pereira, M.O. Pseudomonas aeruginosa Diversification during Infection Development in Cystic Fibrosis Lungs-A Review. Pathogens 2014, 3, 680–703. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Ciofu, O.; Bjarnsholt, T. Pseudomonas aeruginosa Biofilms in Cystic Fibrosis. Future Microbiol. 2010, 5, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, C.; Carbone, D.; Parrino, B.; Cascioferro, S.; Diana, P. Recent Developments in the Inhibition of Bacterial Adhesion as Promising Anti-Virulence Strategy. Int. J. Mol. Sci. 2023, 24, 4872. [Google Scholar] [CrossRef] [PubMed]
- Parrino, B.; Carbone, D.; Cirrincione, G.; Diana, P.; Cascioferro, S. Inhibitors of Antibiotic Resistance Mechanisms: Clinical Applications and Future Perspectives. Future Med. Chem. 2020, 12, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, O.M.; Al-Gamal, M.S.; Ibrahim, G.A.; Dabiza, N.M.; Salem, S.S.; El-ssayad, M.F.; Youssef, A.M. Evaluation and Characterization of Some Protective Culture Metabolites in Free and Nano-Chitosan-Loaded Forms against Common Contaminants of Egyptian Cheese. Carbohydr. Polym. 2019, 223, 115094. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.; Grassiri, B.; Esin, S.; Kaya, E.; Maisetta, G.; Piras, A.M.; Batoni, G. Inhaled Lacticaseibacillus rhamnosus-Derived Postbiotics as Novel Strategy to Tackle Pseudomonas Aeruginosa Lung Infection in Cystic Fibrosis. In Proceedings of the Drug Delivery to the Lung Conference (DDL), as a Poster, Edimburgh, Scotland, 6–8 December 2023. [Google Scholar]
- Sriramulu, D.D.; Lünsdorf, H.; Lam, J.S.; Römling, U. Microcolony Formation: A Novel Biofilm Model of Pseudomonas aeruginosa for the Cystic Fibrosis Lung. J. Med. Microbiol. 2005, 54, 667–676. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, M.; Esin, S.; Kaya, E.; Batoni, G.; Maisetta, G. Anti-Persisters Activity of Lacticaseibacillus rhamnosus Culture Filtrates against Pseudomonas aeruginosa in Artificial Sputum Medium. Int. J. Mol. Sci. 2024, 25, 7113. https://doi.org/10.3390/ijms25137113
Bianchi M, Esin S, Kaya E, Batoni G, Maisetta G. Anti-Persisters Activity of Lacticaseibacillus rhamnosus Culture Filtrates against Pseudomonas aeruginosa in Artificial Sputum Medium. International Journal of Molecular Sciences. 2024; 25(13):7113. https://doi.org/10.3390/ijms25137113
Chicago/Turabian StyleBianchi, Marta, Semih Esin, Esingül Kaya, Giovanna Batoni, and Giuseppantonio Maisetta. 2024. "Anti-Persisters Activity of Lacticaseibacillus rhamnosus Culture Filtrates against Pseudomonas aeruginosa in Artificial Sputum Medium" International Journal of Molecular Sciences 25, no. 13: 7113. https://doi.org/10.3390/ijms25137113
APA StyleBianchi, M., Esin, S., Kaya, E., Batoni, G., & Maisetta, G. (2024). Anti-Persisters Activity of Lacticaseibacillus rhamnosus Culture Filtrates against Pseudomonas aeruginosa in Artificial Sputum Medium. International Journal of Molecular Sciences, 25(13), 7113. https://doi.org/10.3390/ijms25137113