
Regulatory Effects of 198-bp Structural Variants in the GSTA2 Promoter Region on Adipogenesis in Chickens
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Conservation and Protein Interactions of the GSTA2 Gene
2.2. Identification and Genotyping of the 198-bp SV
2.3. Genetic Diversity of the 198-bp SV
2.4. Differential Selection of the 198-bp SV
2.5. Association of the 198-bp SV with Chicken Carcass and Meat Traits
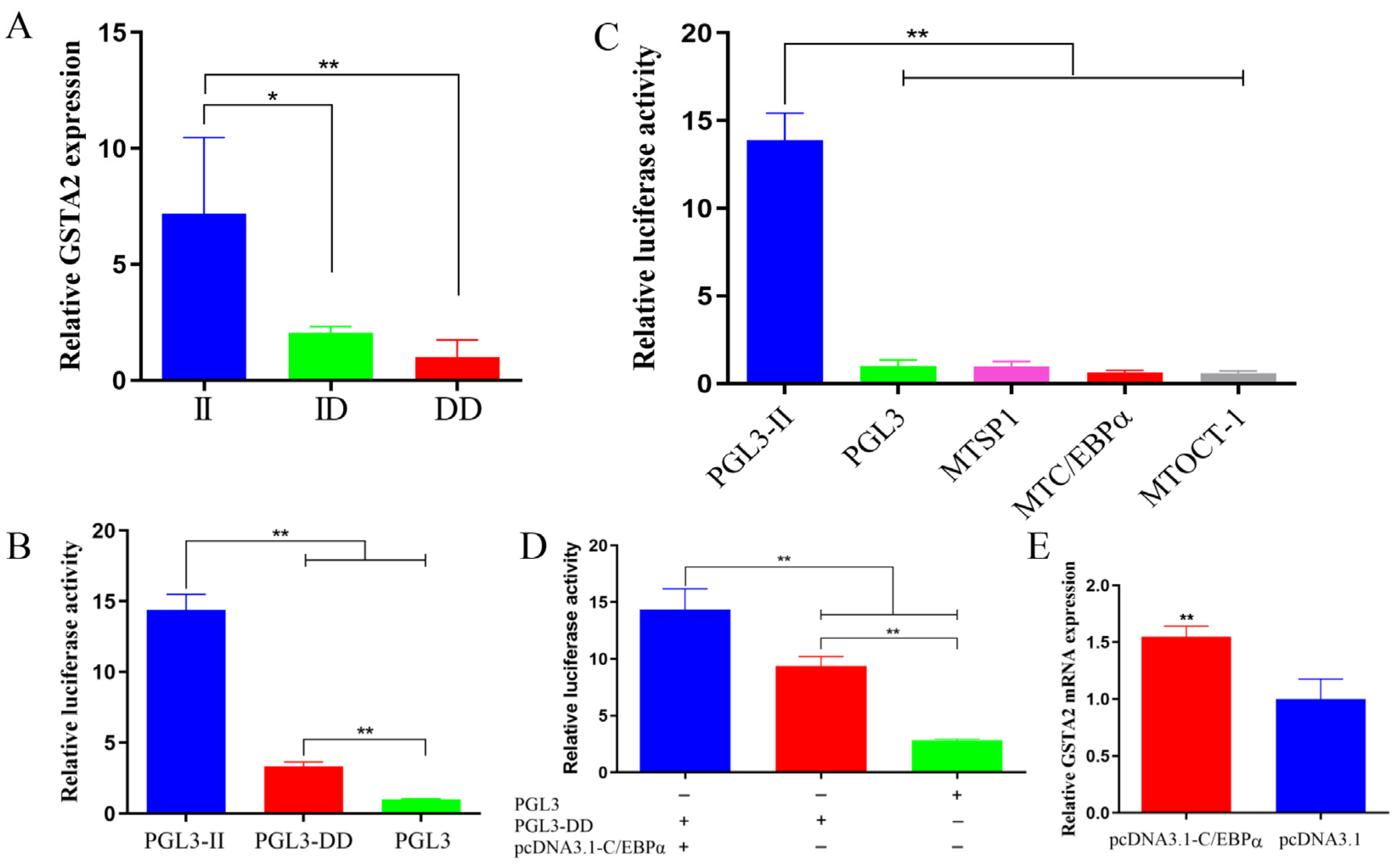
2.6. Expression of GSTA2 Gene
2.7. Relative Expression of GSTA2 in Different Genotypes
2.8. Promoter Activity of GSTA2
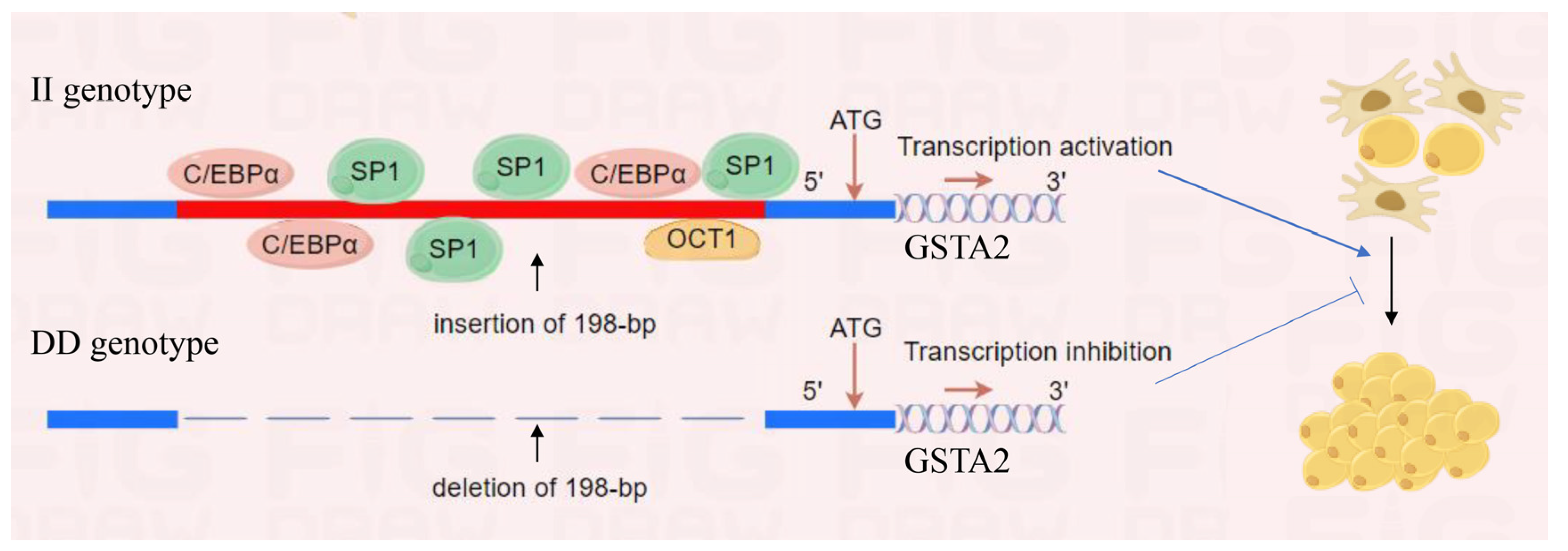
2.9. Prediction and Validation of Transcription Factors in the 198-bp SV
2.10. GSTA2 Promotes the Proliferation of ICP1
2.11. GSTA2 Promotes the Differentiation of ICP1
3. Discussion
4. Materials and Methods
4.1. Collection of Animal Samples
4.2. Bioinformatics Analysis
4.3. Genetic Variation and Genotyping
4.4. Cell Culture
4.5. cDNA Synthesis and Quantitative Real-Time PCR (qPCR)
4.6. Dual-Luciferase Reporter Assay
4.7. RNA Oligonucleotides, Plasmid Construction and Cell Transfection
4.8. Flow Cytometry Assays
4.9. 5-Ethynyl-2′-deoxyuridine (EdU) Assay
4.10. Oil Red O Staining
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pandit, R.J.; Hinsu, A.T.; Patel, N.V.; Koringa, P.G.; Jakhesara, S.J.; Thakkar, J.R.; Shah, T.M.; Limon, G.; Psifidi, A.; Guitian, J. Microbial diversity and community composition of caecal microbiota in commercial and indigenous Indian chickens determined using 16s rDNA amplicon sequencing. Microbiome 2018, 6, 115. [Google Scholar] [CrossRef]
- Lu, Q.; Wen, J.; Zhang, H. Effect of chronic heat exposure on fat deposition and meat quality in two genetic types of chicken. Poultry Sci. 2007, 86, 1059–1064. [Google Scholar] [CrossRef]
- Siegel, P.B. Evolution of the modern broiler and feed efficiency. Annu. Rev. Anim. Biosci. 2014, 2, 375–385. [Google Scholar] [CrossRef]
- Tallentire, C.W.; Leinonen, I.; Kyriazakis, I. Breeding for efficiency in the broiler chicken: A review. Agron. Sustain. Dev. 2016, 36, 66. [Google Scholar] [CrossRef]
- Wen, C.; Yan, W.; Sun, C.; Ji, C.; Zhou, Q.; Zhang, D.; Zheng, J.; Yang, N. The gut microbiota is largely independent of host genetics in regulating fat deposition in chickens. ISME J. 2019, 13, 1422–1436. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Vigilanza, P.; Aquilano, K.; Baldelli, S.; Rotilio, G.; Ciriolo, M.R. Modulation of intracellular glutathione affects adipogenesis in 3T3-L1 cells. J. Cell. Physiol. 2011, 226, 2016–2024. [Google Scholar] [CrossRef]
- Morel, F.; Aninat, C. The glutathione transferase kappa family. Drug Metab. Rev. 2011, 43, 281–291. [Google Scholar] [CrossRef]
- Sheehan, D.; Meade, G.; Foley, V.M.; Dowd, C.A. Structure, function and evolution of glutathione transferases: Implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem J. 2001, 360, 1–16. [Google Scholar] [CrossRef]
- Choi, Y.J.; Shin, M.J.; Youn, G.S.; Park, J.H.; Yeo, H.J.; Yeo, E.J.; Kwon, H.J.; Lee, L.R.; Kim, N.Y.; Kwon, S.Y. Protective Effects of PEP-1-GSTA2 Protein in Hippocampal Neuronal Cell Damage Induced by Oxidative Stress. Int. J. Mol. Sci. 2023, 24, 2767. [Google Scholar] [CrossRef]
- Kang, K.W.; Lee, S.J.; Kim, S.G. Molecular mechanism of nrf2 activation by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Boušová, I.; Košťáková, Š.; Matoušková, P.; Bartikova, H.; Szotakova, B.; Skalova, L. Monosodium glutamate-induced obesity changed the expression and activity of glutathione S-transferases in mouse heart and kidney. Pharm. Int. J. Pharm. Sci. 2017, 72, 257–259. [Google Scholar]
- Guang-Qi, G.; Li-Shuang, S.; Bin, T.; Guang-Peng, L. Expression levels of GSTA2 and APOD genes might be associated with carotenoid coloration in golden pheasant (Chrysolophus pictus) plumage. Zool Res. 2016, 37, 144. [Google Scholar]
- Tripathy, P.S.; Devi, N.C.; Parhi, J.; Priyadarshi, H.; Patel, A.B.; Pandey, P.K.; Mandal, S.C. Molecular mechanisms of natural carotenoid-based pigmentation of queen loach, Botia dario (Hamilton, 1822) under captive condition. Sci. Rep. 2019, 9, 12585. [Google Scholar] [CrossRef] [PubMed]
- Weischenfeldt, J.; Symmons, O.; Spitz, F.; Korbel, J.O. Phenotypic impact of genomic structural variation: Insights from and for human disease. Nat. Rev. Genet. 2013, 14, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Ebert, P.; Audano, P.A.; Zhu, Q.; Rodriguez-Martin, B.; Porubsky, D.; Bonder, M.J.; Sulovari, A.; Ebler, J.; Zhou, W.; Serra Mari, R. Haplotype-resolved diverse human genomes and integrated analysis of structural variation. Science 2021, 372, eabf7117. [Google Scholar] [CrossRef] [PubMed]
- Roses, A.D.; Akkari, P.A.; Chiba-Falek, O.; Lutz, M.W.; Gottschalk, W.K.; Saunders, A.M.; Saul, B.; Sundseth, S.; Burns, D. Structural variants can be more informative for disease diagnostics, prognostics and translation than current SNP mapping and exon sequencing. Expert Opin. Drug Metab. 2016, 12, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.; Scott, A.J.; Davis, J.R.; Tsang, E.K.; Li, X.; Kim, Y.; Hadzic, T.; Damani, F.N.; Ganel, L.; Consortium, G. The impact of structural variation on human gene expression. Nat. Genet. 2017, 49, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, D.; Sebat, J. CNVs: Harbingers of a rare variant revolution in psychiatric genetics. Cell 2012, 148, 1223–1241. [Google Scholar] [CrossRef]
- Sudmant, P.H.; Rausch, T.; Gardner, E.J.; Handsaker, R.E.; Abyzov, A.; Huddleston, J.; Zhang, Y.; Ye, K.; Jun, G.; Hsi-Yang Fritz, M. An integrated map of structural variation in 2,504 human genomes. Nature 2015, 526, 75–81. [Google Scholar] [CrossRef]
- Kou, Y.; Liao, Y.; Toivainen, T.; Lv, Y.; Tian, X.; Emerson, J.; Gaut, B.S.; Zhou, Y. Evolutionary genomics of structural variation in Asian rice (Oryza sativa) domestication. Mol. Biol. Evol. 2020, 37, 3507–3524. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhang, W.-Y.; Yang, B.-G.; Zhou, D.-K.; Xu, L.; He, Y.-M.; Zhang, H.-Y.; Liu, C.-L.; Ma, Y.-H.; Chu, M.-X. A 1.1 Mb duplication CNV on chromosome 17 contributes to skeletal muscle development in Boer goats. Zool Res. 2023, 44, 303. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Wang, J.; Zhao, R.; Niu, N.; Su, Y.; Hu, Z.; Liu, X.; Hou, X.; Wang, L.; Wang, L. Associations of genome-wide structural variations with phenotypic differences in cross-bred Eurasian pigs. J. Anim. Sci. Biotechnol. 2023, 14, 136. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, D.; Tang, S.; Li, D.; Han, R.; Tian, Y.; Li, H.; Li, G.; Li, W.; Liu, X. A multiallelic indel in the promoter region of the Cyclin-dependent kinase inhibitor 3 gene is significantly associated with body weight and carcass traits in chickens. Poultry Sci. 2019, 98, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Shahmuradov, I.A.; Gammerman, A.J.; Hancock, J.M.; Bramley, P.M.; Solovyev, V.V. PlantProm: A database of plant promoter sequences. Nucleic Acids Res. 2003, 31, 114–117. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Li, W.; Liu, D.; Liang, K.; Wang, X.; Li, H.; Jiang, R.; Tian, Y.; Kang, X.; Li, Z. Two insertion/deletion variants in the promoter region of the QPCTL gene are significantly associated with body weight and carcass traits in chickens. Anim. Genet. 2019, 50, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Lin, W.; Yang, X.; Zhang, Z.; He, S.; Li, W.; Li, Z.; Zhang, X. QPCTL Affects the Daily Weight Gain of the F2 Population and Regulates Myogenic Cell Proliferation and Differentiation in Chickens. Animals 2022, 12, 3535. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, T.; Zhou, Y.; Liu, M.; Klaus, S.; Lan, X.; Lei, C.; Chen, H. A novel PAX7 10-bp indel variant modulates promoter activity, gene expression and contributes to different phenotypes of Chinese cattle. Sci. Rep. 2018, 8, 1724. [Google Scholar] [CrossRef] [PubMed]
- Wiener, P.; Wilkinson, S. Deciphering the genetic basis of animal domestication. Proc. R. Soc. B-Biol. Sci. 2011, 278, 3161–3170. [Google Scholar] [CrossRef]
- Jia, X.; Lin, H.; Nie, Q.; Zhang, X.; Lamont, S.J. A short insertion mutation disrupts genesis of miR-16 and causes increased body weight in domesticated chicken. Sci. Rep. 2016, 6, 36433. [Google Scholar] [CrossRef]
- Madeira, M.; Costa, P.; Alfaia, C.; Lopes, P.; Bessa, R.; Lemos, J.C.; Prates, J. The increased intramuscular fat promoted by dietary lysine restriction in lean but not in fatty pig genotypes improves pork sensory attributes. J. Anim. Sci. 2013, 91, 3177–3187. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Tabuchi, T.; Tamaki, Y.; Kosaka, K.; Takikawa, Y.; Satoh, T. Carnosic acid and carnosol inhibit adipocyte differentiation in mouse 3T3-L1 cells through induction of phase2 enzymes and activation of glutathione metabolism. Biochem. Biophys. Res. Commun. 2009, 382, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177. [Google Scholar] [PubMed]
- Wei, C.; Niu, Y.; Chen, B.; Qin, P.; Wang, Y.; Hou, D.; Li, T.; Li, R.; Wang, C.; Yin, H. Genetic effect of an InDel in the promoter region of the NUDT15 and its effect on myoblast proliferation in chickens. BMC Genom. 2022, 23, 138. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Zhang, Z.; Fu, R.; Yang, Y.; Li, W.; Liang, J.; Mo, G.; Luo, W.; Zhang, X. A 51 bp indel polymorphism within the PTH1R gene is significantly associated with chicken growth and carcass traits. Anim. Genet. 2020, 51, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBPα and PPARγ controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell 1999, 3, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Ren, T.; Li, W.; Liu, M.; He, D.; Liang, S.; Luo, W.; Zhang, X. Novel 61-bp indel of RIN2 is associated with fat and hatching weight traits in chickens. Front. Genet. 2021, 12, 672888. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, T.; Tahara, K.; Takano, S.; Inoue-Murayama, M.; Rose, M.T.; Minashima, T.; Aso, H.; Ito, S.i. Mechanism of plasma glutathione peroxidase production in bovine adipocytes. Cell Tissue Res. 2006, 326, 139–147. [Google Scholar] [CrossRef]
- Pu, L.; Luo, Y.; Wen, Z.; Dai, Y.; Zheng, C.; Zhu, X.; Qin, L.; Zhang, C.; Liang, H.; Zhang, J. GPX2 Gene Affects Feed Efficiency of Pigs by Inhibiting Fat Deposition and Promoting Muscle Development. Animals 2022, 12, 3528. [Google Scholar] [CrossRef]
- Urs, S.; Smith, C.; Campbell, B.; Saxton, A.M.; Taylor, J.; Zhang, B.; Snoddy, J.; Jones Voy, B.; Moustaid-Moussa, N. Gene expression profiling in human preadipocytes and adipocytes by microarray analysis. J. Nutr. 2004, 134, 762–770. [Google Scholar] [CrossRef]
- Menon, D.; Innes, A.; Oakley, A.J.; Dahlstrom, J.E.; Jensen, L.M.; Brüstle, A.; Tummala, P.; Rooke, M.; Casarotto, M.G.; Baell, J.B. GSTO1-1 plays a pro-inflammatory role in models of inflammation, colitis and obesity. Sci. Rep. 2017, 7, 17832. [Google Scholar] [CrossRef]
- Cartoni Mancinelli, A.; Di Veroli, A.; Mattioli, S.; Cruciani, G.; Dal Bosco, A.; Castellini, C. Lipid metabolism analysis in liver of different chicken genotypes and impact on nutritionally relevant polyunsaturated fatty acids of meat. Sci. Rep. 2022, 12, 1888. [Google Scholar] [CrossRef]
- Zhang, J.; Cai, B.; Ma, M.; Luo, W.; Zhang, Z.; Zhang, X.; Nie, Q. ALDH1A1 inhibits chicken preadipocytes’ proliferation and differentiation via the PPARγ pathway in vitro and in vivo. Int. J. Mol. Sci. 2020, 21, 3150. [Google Scholar] [CrossRef]
- Yasmeen, R.; Reichert, B.; Deiuliis, J.; Yang, F.; Lynch, A.; Meyers, J.; Sharlach, M.; Shin, S.; Volz, K.S.; Green, K.B. Autocrine function of aldehyde dehydrogenase 1 as a determinant of diet-and sex-specific differences in visceral adiposity. Diabetes 2013, 62, 124–136. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Koutnikova, H.; Auwerx, J. Regulation of adipocyte differentiation. Ann. Med. 2001, 33, 556–561. [Google Scholar] [CrossRef]
- Lowell, B.B. An essential regulator of adipogenesis and modulator of fat cell function: PPARγ. Cell 1999, 99, 239–242. [Google Scholar] [CrossRef]
- Yeh, W.-C.; Cao, Z.; Classon, M.; McKnight, S.L. Cascade regulation of terminal adipocyte differentiation by three members of the C/EBP family of leucine zipper proteins. Genes Dev. 1995, 9, 168–181. [Google Scholar] [CrossRef]
- Mandrup, S.; Lane, M.D. Regulating adipogenesis. J. Biol. Chem. 1997, 272, 5367–5370. [Google Scholar] [CrossRef]
- Abdalla, B.A.; Chen, X.; Li, K.; Chen, J.; Yi, Z.; Zhang, X.; Li, Z.; Nie, Q. Control of preadipocyte proliferation, apoptosis and early adipogenesis by the forkhead transcription factor FoxO6. Life Sci. 2021, 265, 118858. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Huang, W.; Wang, Z.; Nie, Q. m6A demethylase FTO regulate CTNNB1 to promote adipogenesis of chicken preadipocyte. J. Anim. Sci. Biotechnol. 2022, 13, 147. [Google Scholar] [CrossRef]
- Cui, H.; Liu, L.; Liu, X.; Wang, Y.; Luo, N.; Tan, X.; Zhu, Y.; Liu, R.; Zhao, G.; Wen, J. A selected population study reveals the biochemical mechanism of intramuscular fat deposition in chicken meat. J. Anim. Sci. Biotechnol. 2022, 13, 54. [Google Scholar] [CrossRef]
- Lei, M.; Nie, Q.; Peng, X.; Zhang, D.; Zhang, X. Single nucleotide polymorphisms of the chicken insulin-like factor binding protein 2 gene associated with chicken growth and carcass traits. Poultry Sci. 2005, 84, 1191–1198. [Google Scholar] [CrossRef]
- Hou, Y.; Qi, F.; Bai, X.; Ren, T.; Shen, X.; Chu, Q.; Zhang, X.; Lu, X. Genome-wide analysis reveals molecular convergence underlying domestication in 7 bird and mammals. BMC Genom. 2020, 21, 204. [Google Scholar] [CrossRef]
- Yong, Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeds/n | Genotypic and Allelic Frequencies | Ho | He | Ne | PIC | p-Value (HWE) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| DD | ID | II | D | I | ||||||
| F2/359 | 0.00 | 0.18 | 0.82 | 0.09 | 0.91 | 0.18 | 0.17 | 1.20 | 0.15 | 0.062 |
| N409/428 | 0.11 | 0.35 | 0.54 | 0.29 | 0.71 | 0.35 | 0.41 | 1.69 | 0.32 | 0.004 |
| ND/71 | 0.00 | 0.04 | 0.96 | 0.02 | 0.98 | 0.04 | 0.04 | 1.04 | 0.04 | 0.856 |
| GS/50 | 0.00 | 0.26 | 0.74 | 0.13 | 0.87 | 0.26 | 0.23 | 1.29 | 0.20 | 1.00 |
| GX/69 | 0.19 | 0.32 | 0.49 | 0.35 | 0.65 | 0.32 | 0.45 | 1.83 | 0.35 | 0.014 |
| WC/46 | 0.00 | 0.17 | 0.83 | 0.09 | 0.91 | 0.17 | 0.16 | 1.19 | 0.15 | 0.518 |
| QY/60 | 0.05 | 0.28 | 0.67 | 0.19 | 0.81 | 0.28 | 0.31 | 1.45 | 0.26 | 0.507 |
| ISA/67 | 0.01 | 0.15 | 0.84 | 0.09 | 0.91 | 0.15 | 0.16 | 1.19 | 0.15 | 0.488 |
| RW/55 | 0.00 | 0.00 | 1.00 | 0.00 | 1.00 | 0.00 | 0.00 | 1.00 | 0.00 | NA |
| Traits | Mean ± SEM | p-Value | |
|---|---|---|---|
| II | ID | ||
| SFT (mm) | 4.183 ± 0.068 a | 3.814 ± 0.146 b | 0.022 |
| AFW (g) | 28.98 ± 0.983 a | 23.684 ± 2.151 b | 0.025 |
| LMCF (%) | 3.662 ± 0.221 a | 3.108 ± 0.265 b | 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Xu, M.; Zhang, Z.; Liang, J.; Fu, R.; Lin, W.; Luo, W.; Zhang, X.; Ren, T. Regulatory Effects of 198-bp Structural Variants in the GSTA2 Promoter Region on Adipogenesis in Chickens. Int. J. Mol. Sci. 2024, 25, 7155. https://doi.org/10.3390/ijms25137155
Li W, Xu M, Zhang Z, Liang J, Fu R, Lin W, Luo W, Zhang X, Ren T. Regulatory Effects of 198-bp Structural Variants in the GSTA2 Promoter Region on Adipogenesis in Chickens. International Journal of Molecular Sciences. 2024; 25(13):7155. https://doi.org/10.3390/ijms25137155
Chicago/Turabian StyleLi, Wangyu, Meng Xu, Zihao Zhang, Jiaying Liang, Rong Fu, Wujian Lin, Wen Luo, Xiquan Zhang, and Tuanhui Ren. 2024. "Regulatory Effects of 198-bp Structural Variants in the GSTA2 Promoter Region on Adipogenesis in Chickens" International Journal of Molecular Sciences 25, no. 13: 7155. https://doi.org/10.3390/ijms25137155