
Effects of Nitrosyl Iron Complexes with Thiol, Phosphate, and Thiosulfate Ligands on Hemoglobin
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
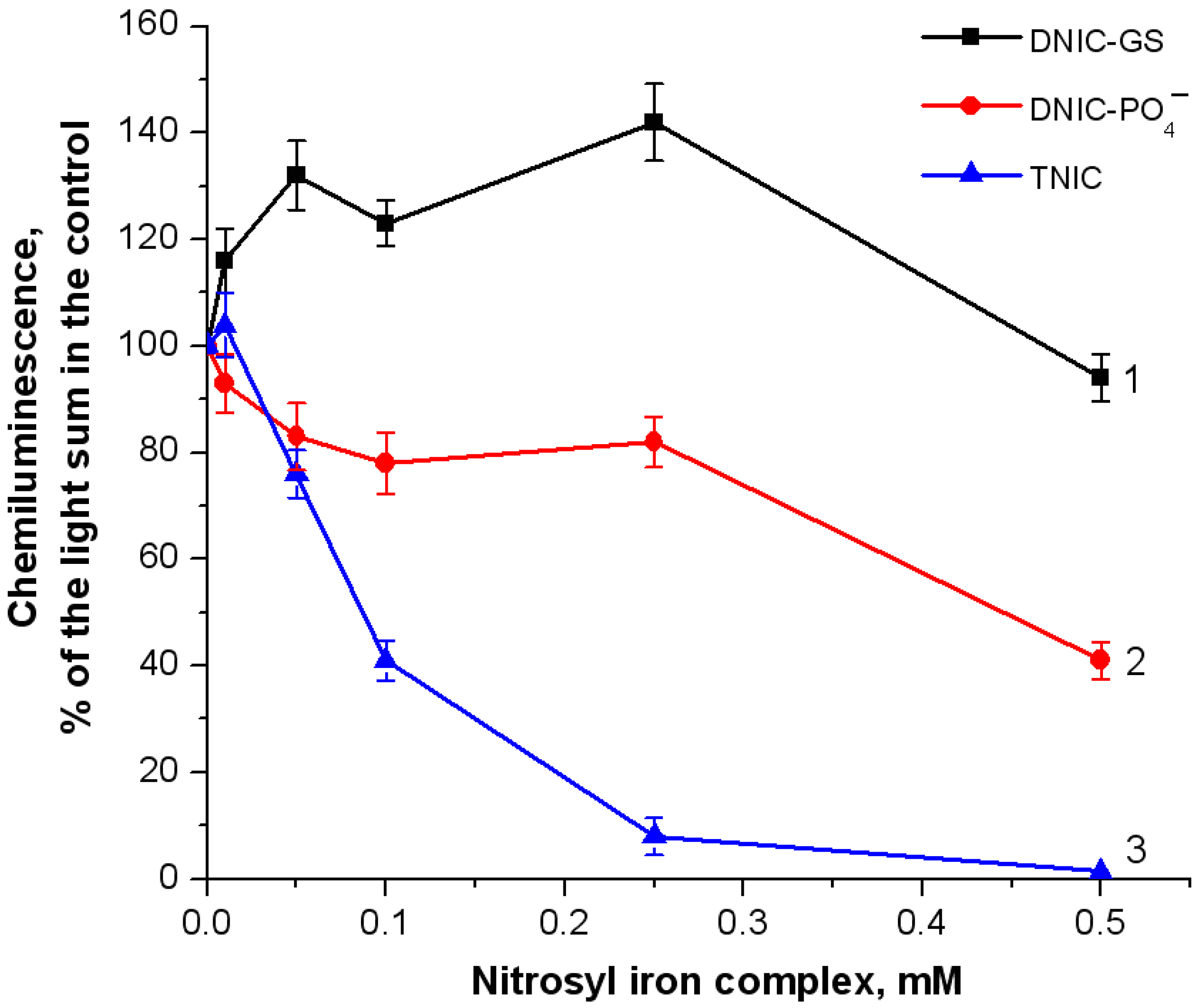
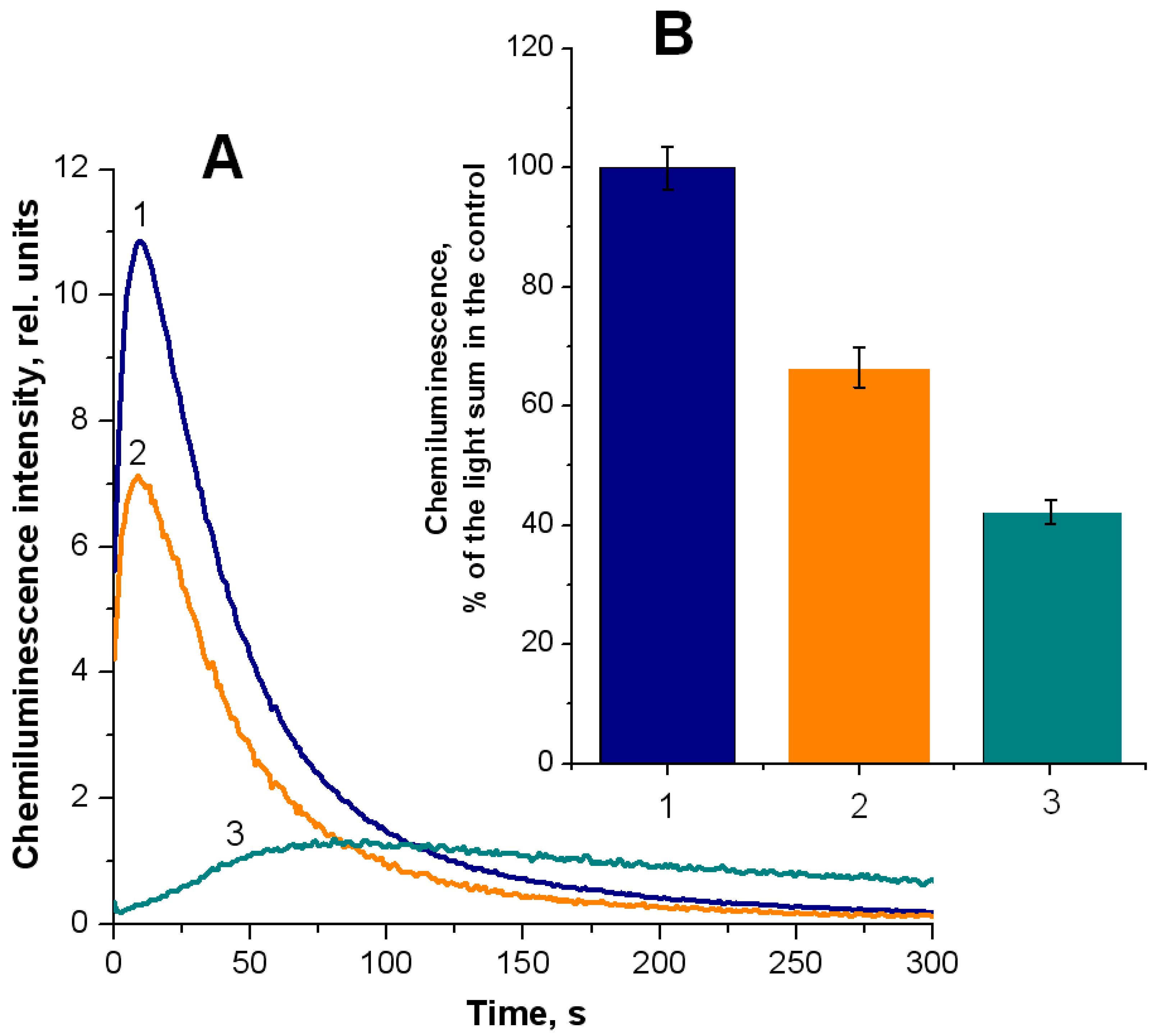
2.1. Formation of Free Radical Products during Hemoglobin Oxidation with Tert-Butyl Hydroperoxide
2.2. Oxidative Modifications of Hemoglobin
Hb-Fe3+ + 2NO• → Hb-Fe2+-NO + NO2− + H+
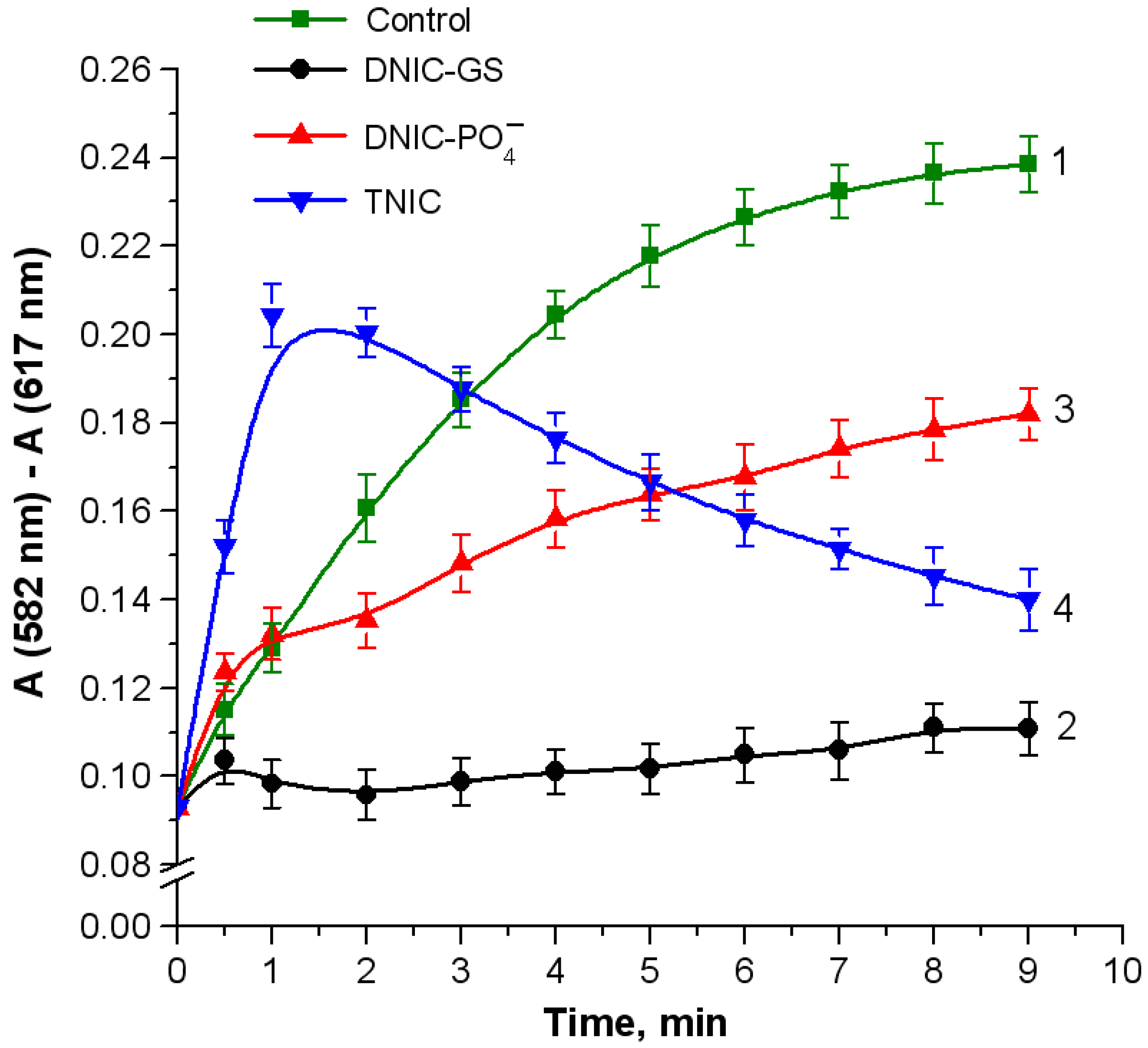
2.3. Oxidation of Linoleic Acid in Hemoglobin/Tert-Butyl Hydroperoxide System
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Measurement of Luminol-Dependent Chemiluminescence
4.3. Obtaining and Registering OxoferrylHb
4.4. Formation of the Products of Hemoglobin Oxidation in the Reaction with TMPD
4.5. Antioxidant Activity Measurement with ABTS
4.6. Measurement of Linoleic Acid Diene Conjugates
4.7. Determination of the Content of the Heme Group
4.8. SDS-PAGE Electrophoresis
4.9. Spectrophotometric Studies
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lewandowska, H.; Kalinowska, M.; Brzoska, K.; Wojciuk, K.; Wojciuk, G.; Kruszewski, M. Nitrosyl iron complexes—Synthesis, structure and biology. Dalton Trans. 2011, 40, 8273–8289. [Google Scholar] [CrossRef] [PubMed]
- Lundberg, J.O.; Weitzberg, E. Nitric oxide signaling in health and disease. Cell 2022, 185, 2853–2878. [Google Scholar] [CrossRef] [PubMed]
- Ferrer-Sueta, G.; Campolo, N.; Trujillo, M.; Bartesaghi, S.; Carballal, S.; Romero, N.; Alvarez, B.; Radi, R. Biochemistry of peroxynitrite and protein tyrosine nitration. Chem. Rev. 2018, 118, 1338–1408. [Google Scholar] [CrossRef] [PubMed]
- Raffetto, J.D.; Calanni, F.; Mattana, P.; Khalil, R.A. Sulodexide promotes arterial relaxation via endothelium-dependent nitric oxide-mediated pathway. Biochem. Pharmacol. 2019, 166, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, I. Nitric oxide mediated redox regulation of protein homeostasis. Cell. Signal. 2019, 53, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Dey, S.K.; Kundu, S. Functional implications of vascular endothelium in regulation of endothelial nitric oxide synthesis to control blood pressure and cardiac functions. Life Sci. 2020, 259, 118377. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Jin, A.; Yang, Z.; Huang, W. Advanced nitric oxide generating nanomedicine for therapeutic applications. ACS Nano 2023, 17, 8935–8965. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-y.; Ye, Z.-x.; Wang, X.-f.; Chang, J.; Yang, M.-w.; Zhong, H.-h.; Hong, F.-f.; Yang, S.-l. Nitric oxide bioavailability dysfunction involves in atherosclerosis. Biomed. Pharmacother. 2018, 97, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Maharana, K.C.; Meenakshi, S.; Singh, S. Endothelial dysfunction and its relation in different disorders: Recent update. Health Sci. Rev. 2023, 7, 100084. [Google Scholar] [CrossRef]
- Xia, Z.; Vanhoutte, P.M. Nitric oxide and protection against cardiac ischemia. Curr. Pharm. Des. 2011, 17, 1774–1782. [Google Scholar] [CrossRef] [PubMed]
- Pisarenko, O.; Studneva, I.; Timoshin, A.; Veselova, O. Protective efficacy of dinitrosyl iron complexes with reduced glutathione in cardioplegia and reperfusion. Pflug. Arch. 2019, 471, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Sanina, N.A.; Kozub, G.I.; Kondrat’eva, T.A.; Korchagin, D.V.; Shilov, G.V.; Morgunov, R.B.; Ovanesyan, N.S.; Kulikov, A.V.; Stupina, T.S.; Terent’ev, A.A.; et al. Anionic dinitrosyl iron complexes—New nitric oxide donors with selective toxicity to human glioblastoma cells. J. Mol. Struct. 2022, 1266, 133506. [Google Scholar] [CrossRef]
- Amozova, V.I.; Balakina, A.A.; Mishchenko, D.V.; Sanina, N.A. Effect of the dinitrosyl iron complex with N-ethylthiourea on ROS and NO intracellular levels and caspase activity in HeLa tumor cells. Russ. Chem. Bull. 2023, 72, 1066–1074. [Google Scholar] [CrossRef]
- Wu, M.; Lu, Z.; Wu, K.; Nam, C.; Zhang, L.; Guo, J. Recent advances in the development of nitric oxide-releasing biomaterials and their application potentials in chronic wound healing. J. Mater. Chem. B 2021, 9, 7063–7075. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, Z.; Li, L.L. Advanced nitric oxide donors: Chemical structure of NO drugs, NO nanomedicines and biomedical applications. Nanoscale 2021, 13, 444–459. [Google Scholar] [CrossRef] [PubMed]
- Sanina, N.A.; Aldoshin, S.M.; Shmatko, N.Y.; Korchagin, D.V.; Shilov, G.V.; Knyazkina, E.V.; Ovanesyan, N.S.; Kulikov, A.V. Nitrosyl iron complexes with enhanced NO donating ability: Synthesis, structure and properties of a new type of salt with the DNIC cations [Fe(SC(NH2)2)2(NO)2]+. New J. Chem. 2015, 39, 1022–1030. [Google Scholar] [CrossRef]
- Ahmed, R.; Augustine, R.; Chaudhry, M.; Akhtar, U.A.; Zahid, A.A.; Tariq, M.; Falahati, M.; Ahmad, I.S.; Hasan, A. Nitric oxide-releasing biomaterials for promoting wound healing in impaired diabetic wounds: State of the art and recent trends. Biomed. Pharmacother. 2022, 149, 112707. [Google Scholar] [CrossRef] [PubMed]
- Vanin, A.F.; Mokh, V.P.; Serezhenkov, V.A.; Chazov, E.I. Vasorelaxing activity of stable powder preparations of dinitrosyl iron complexes with cysteine or glutathione ligands. Nitric Oxide 2007, 16, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Vanin, A.F. Positive (regulatory) and negative (cytotoxic) effects of dinitrosyl iron complexes on living organisms. Biochemistry (Moscow) 2022, 87, 1367–1386. [Google Scholar] [CrossRef] [PubMed]
- Bouché, M.; Hognon, C.; Grandemange, S.; Gros, P.C. Recent advances in iron-complexes as drug candidates for cancer therapy: Reactivity, mechanism of action and metabolites. Dalton Trans. 2020, 49, 11451–11466. [Google Scholar] [CrossRef]
- Lu, T.-T.; Liaw, W.-F. Insight into the electronic structure of biomimetic dinitrosyliron complexes (DNICs): Toward the syntheses of amido-bridging dinuclear DNICs. Inorg. Chem. 2021, 60, 15846–15873. [Google Scholar] [CrossRef]
- Chuang, W.-H.; Chou, Y.-T.; Chen, Y.-H.; Kuo, T.-H.; Liaw, W.-F.; Lu, T.-T.; Kao, C.-F.; Wang, Y.-M. Neuroprotective effect of NO-delivery dinitrosyl iron complexes (DNICs) on amyloid pathology in the Alzheimer’s disease cell model. ACS Chem. Neurosci. 2023, 14, 2922–2934. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-C.; Lee, G.-H.; Chien, S.-Y.; Lee, C.-M. Light-induced NO release from iron-nitrosyl-thiolato complex: The role of noncovalent thiol/thioether. J. Chin. Chem. Soc. 2023, 70, 1125. [Google Scholar] [CrossRef]
- Pokidova, O.V.; Emel’yanova, N.S.; Psikha, B.L.; Kulikov, A.V. Decomposition of the binuclear nitrosyl iron complex with thiosulfato ligands in aqueous solutions: Experimental and theoretical study. Inorganica Chim. Acta 2020, 502, 119369. [Google Scholar] [CrossRef]
- Kanner, J.; Harel, S.; Granit, R. Nitric oxide as an antioxidant. Arch. Biochem. Biophys. 1991, 289, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Wink, D.A.; Mitchell, J.B. Chemical biology of nitric oxide: Insights into regulatory, cytotoxic, and cytoprotective mechanisms of nitric oxide. Free Radic. Biol. Med. 1998, 25, 434–456. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Osipov, A.N.; Day, B.W.; Zayas-Rivera, B.; Kagan, V.E.; Elsayed, N.M. Reduction of ferrylmyoglobin and ferrylhemoglobin by nitric oxide: A protective mechanism against ferryl hemoprotein-induced oxidations. Biochemistry 1995, 34, 6689–6699. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Tyurina, Y.Y.; Salama, G.; Day, B.W.; Claycamp, H.G.; Argyros, G.; Elsayed, N.M.; Kagan, V.E. Nitric oxide protects cardiomyocytes against tert-butyl hydroperoxide-induced formation of alkoxyl and peroxyl radicals and peroxidation of phosphatidylserine. Biochem. Biophys. Res. Commun. 1998, 244, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Hogg, N.; Kalyanaraman, B. Nitric oxide and lipid peroxidation. Biochim. Biophys. Acta (BBA)-Bioenerg. 1999, 1411, 378–384. [Google Scholar] [CrossRef]
- Shumaev, K.B.; Petrova, N.E.; Zabbarova, I.V.; Vanin, A.F.; Topunov, A.F.; Lankin, V.Z.; Ruuge, E.K. Interaction of oxoferrylmyoglobin and dinitrosyl-iron complexes. Biochemistry (Moscow) 2004, 69, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Shumaev, K.B.; Kosmachevskaya, O.V.; Timoshin, A.A.; Vanin, A.F.; Topunov, A.F. Dinitrosyl iron complexes bind with hemoglobin as markers of oxidative stress. Methods Enzymol. 2008, 436, 445–461. [Google Scholar] [CrossRef] [PubMed]
- Shumaev, K.B.; Gorudko, I.V.; Kosmachevskaya, O.V.; Grigorieva, D.V.; Panasenko, O.M.; Vanin, A.F.; Topunov, A.F.; Terekhova, M.S.; Sokolov, A.V.; Cherenkevich, S.N.; et al. Protective effect of dinitrosyl iron complexes with glutathione in red blood cell lysis induced by hypochlorous acid. Oxid. Med. Cell. Longev. 2019, 2019, 2798154. [Google Scholar] [CrossRef] [PubMed]
- Hummel, S.G.; Fischer, A.J.; Martin, S.M.; Schafer, F.Q.; Buettner, G.R. Nitric oxide as a cellular antioxidant: A little goes a long way. Free Radic. Biol. Med. 2006, 40, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Hickok, J.R.; Thomas, D.D. Nitric oxide reduces oxidative stress in cancers cells by forming dinitrosyliron complexes. Nitric Oxide 2018, 76, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Kosmachevskaya, O.V.; Nasybullina, E.I.; Shumaev, K.B.; Novikova, N.N.; Topunov, A.F. Protective effect of dinitrosyl iron complexes bound with hemoglobin on oxidative modification by peroxynitrite. Int. J. Mol. Sci. 2021, 22, 13649. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, I.I. Peroxidase activity of human hemoproteins: Keeping the fire under control. Molecules 2018, 23, 2561. [Google Scholar] [CrossRef] [PubMed]
- Karoui, H.; Chalier, F.; Finet, J.P.; Tordo, P. DEPMPO: An efficient tool for the coupled ESR-spin trapping of alkylperoxyl radicals in water. Org. Biomol. Chem. 2011, 9, 2473–2480. [Google Scholar] [CrossRef] [PubMed]
- Karoui, H.; Hogg, N.; Fréjaville, C.; Tordo, P.; Kalyanaraman, B. Characterization of sulfur-centered radical intermediates formed during the oxidation of thiols and sulfite by peroxynitrite. ESR-spin trapping and oxygen uptake studies. J. Biol. Chem. 1996, 271, 6000–6009. [Google Scholar] [CrossRef]
- Reeder, B.J.; Svistunenko, D.A.; Cooper, C.E.; Wilson, M.T. The radical and redox chemistry of myoglobin and hemoglobin: From in vitro studies to human pathology. Antioxid. Redox Signal. 2004, 6, 954–966. [Google Scholar] [CrossRef]
- Kosmachevskaya, O.V.; Nasybullina, E.I.; Pugachenko, I.S.; Novikova, N.N.; Topunov, A.F. Antiglycation and antioxidant effect of nitroxyl towards hemoglobin. Antioxidants 2022, 11, 2007. [Google Scholar] [CrossRef]
- Kûka, M.; Èakste, I.; Kûka, P. Inhibition of formation of conjugated dienes in linseed oil. Proc. Latv. Acad. Sci. Sect. B 2018, 72, 80–84. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X.L. Glutathione: A key component of the cytoplasmic labile iron pool. Biometals 2011, 24, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Toledo, J.C., Jr.; Bosworth, C.A.; Hennon, S.W.; Mahtani, H.A.; Bergonia, H.A.; Lancaster, J.R., Jr. Nitric oxide-induced conversion of cellular chelatable iron into macromolecule-bound paramagnetic dinitrosyliron complexes. J. Biol. Chem. 2008, 283, 28926–28933. [Google Scholar] [CrossRef] [PubMed]
- Bosworth, C.; Toledo, J., Jr.; Zmijewski, J.; Li, Q.; Lancaster, J. Dinitrosyliron complexes and the mechanism(s) of cellular protein nitrosothiol formation from nitric oxide. Proc. Natl. Acad. Sci. USA 2009, 106, 4671–4676. [Google Scholar] [CrossRef] [PubMed]
- Schneppensieper, T.; Wanat, A.; Stochel, G.; van Eldik, R. Mechanistic information on the reversible binding of NO to selected iron(II) chelates from activation parameters. Inorg. Chem. 2002, 41, 2565–2573. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, C.; Mahtani, H.K.; Du, J.; Patel, A.R.; Lancaster, J.R., Jr. Nitrosothiol formation and protection against Fenton chemistry by nitric oxide-induced dinitrosyliron complex formation from anoxia-initiated cellular chelatable iron increase. J. Biol. Chem. 2014, 289, 19917–19927. [Google Scholar] [CrossRef]
- Hsiao, H.-Y.; Chung, C.-W.; Santos, J.; Baluran, O. Villaflores Fe in biosynthesis, translocation, and signal transduction of NO: Toward bioinorganic engineering of dinitrosyl iron complexes into NO-delivery scaffolds for tissue engineering. Dalton Trans. 2019, 48, 9431–9453. [Google Scholar] [CrossRef] [PubMed]
- Vanin, A.F. Dinitrosyl Iron Complexes as a “Working Form” of Nitric Oxide in Living Organisms; Cambridge Scholars Publishing: Cambridge, UK, 2019; 276p. [Google Scholar]
- Cortese-Krott, M.M.; Kelm, M. Endothelial nitric oxide synthase in red blood cells: Key to a new erythrocrine function? Redox Biol. 2014, 2, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Gladwin, M.T.; Grubina, R.; Doyle, M.P. The new chemical biology of nitrite reactions with hemoglobin: R-state catalysis, oxidative denitrosylation, and nitrite reductase/anhydrase. Acc. Chem. Res. 2009, 42, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Kleschyov, A.L.; Alencar, J.L.; Vanin, A.F.; Stoclet, J.C. Nitric oxide transport and storage in the cardiovascular system. Ann. N.-Y. Acad. Sci. 2002, 962, 131–139. [Google Scholar] [CrossRef]
- Włodek, L. Beneficial and harmful effects of thiols. Pol. J. Pharmacol. 2002, 54, 215–223. [Google Scholar] [PubMed]
- Tran, N.G.; Kalyvas, H.; Skodje, K.M.; Hayashi, T.; Moënne-Loccoz, P.; Callan, P.E.; Shearer, J.; Kirschenbaum, L.J.; Kim, E. Phenol nitration induced by an {Fe(NO)2}(10) dinitrosyl iron complex. J. Am. Chem. Soc. 2011, 133, 1184–1187. [Google Scholar] [CrossRef] [PubMed]
- Kalita, A.; Kumar, P.; Mondal, B. Reaction of a copper(II)–nitrosyl complex with hydrogen peroxide: Putative formation of a copper(I)–peroxynitrite intermediate. Chem. Commun. 2012, 48, 4636–4638. [Google Scholar] [CrossRef] [PubMed]
- Lobysheva, I.I.; Serezhenkov, V.A.; Vanin, A.F. Interaction of peroxynitrite and hydrogen peroxide with dinitrosyl iron complexes containing thiol ligands in vitro. Biochemistry (Moscow) 1999, 64, 153–158. [Google Scholar] [PubMed]
- Balagopalakrishna, C.; Abugo, O.O.; Horsky, J.; Manoharan, P.T.; Nagababu, E.; Rifkind, J.M. Superoxide produced in the heme pocket of the beta-chain of hemoglobin reacts with the beta-93 cysteine to produce a thiyl radical. Biochemistry 1998, 37, 13194–13202. [Google Scholar] [CrossRef] [PubMed]
- Rubino, F.M. The redox potential of the β-93-cysteine thiol group in human hemoglobin estimated from in vitro oxidant challenge experiments. Molecules 2021, 26, 2528. [Google Scholar] [CrossRef] [PubMed]
- Vitturi, D.A.; Sun, C.-W.; Harper, V.M.; Thrash-Williams, B.; Cantu-Medellin, N.; Chacko, B.K.; Peng, N.; Dai, Y.; Wyss, J.M.; Townes, T.; et al. Antioxidant functions for the hemoglobin β93 cysteine residue in erythrocytes and in the vascular compartment in vivo. Free Radic. Biol. Med. 2013, 55, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Kapelko, V.I.; Lakomkin, V.L.; Abramov, A.A.; Lukoshkova, E.V.; Undrovinas, N.A.; Khapchaev, A.Y.; Shirinsky, V.P. Protective effects of dinitrosyl iron complexes under oxidative stress in the heart. Oxid. Med. Cell. Longev. 2017, 2017, 9456163. [Google Scholar] [CrossRef] [PubMed]
- Dungel, P.; Perlinger, M.; Weidinger, A.; Redl, H.; Kozlov, A.V. The cytoprotective effect of nitrite is based on the formation of dinitrosyl iron complexes. Free Radic. Biol. Med. 2015, 89, 300–310. [Google Scholar] [CrossRef]
- Akentieva, N.P.; Sanina, N.A.; Gizatullin, A.R.; Shkondina, N.I.; Prikhodchenko, T.R.; Shram, S.I.; Zhelev, N.; Aldoshin, S.M. Cytoprotective effects of dinitrosyl iron complexes on viability of human fibroblasts and cardiomyocytes. Front. Pharmacol. 2019, 10, 1277. [Google Scholar] [CrossRef] [PubMed]
- Pokidova, O.V.; Emel’yanova, N.S.; Kormukhina, A.Y.; Novikova, V.O. Albumin as prospective carrier of nitrosyl iron complex with thiourea and thiosulfate ligands under aerobic conditions. Dalton Trans. 2022, 51, 6473–6485. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Zhou, S.; Gu, L.; Gell, D.A.; Mackay, J.P.; Weiss, M.J.; Gow, A.J.; Shi, Y. Structure of oxidized alpha-haemoglobin bound to AHSP reveals a protective mechanism for haem. Nature 2005, 435, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP decolorization assay of antioxidant capacity reaction pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [PubMed]
- Riggs, A. Preparation of blood hemoglobins of vertebrates. Methods Enzymol. 1981, 76, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Kosmachevskaya, O.V.; Novikova, N.N.; Topunov, A.F. Carbonyl stress in red blood cells and hemoglobin. Antioxidants 2021, 10, 253. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosmachevskaya, O.V.; Nasybullina, E.I.; Pokidova, O.V.; Sanina, N.A.; Topunov, A.F. Effects of Nitrosyl Iron Complexes with Thiol, Phosphate, and Thiosulfate Ligands on Hemoglobin. Int. J. Mol. Sci. 2024, 25, 7194. https://doi.org/10.3390/ijms25137194
Kosmachevskaya OV, Nasybullina EI, Pokidova OV, Sanina NA, Topunov AF. Effects of Nitrosyl Iron Complexes with Thiol, Phosphate, and Thiosulfate Ligands on Hemoglobin. International Journal of Molecular Sciences. 2024; 25(13):7194. https://doi.org/10.3390/ijms25137194
Chicago/Turabian StyleKosmachevskaya, Olga V., Elvira I. Nasybullina, Olesya V. Pokidova, Natalia A. Sanina, and Alexey F. Topunov. 2024. "Effects of Nitrosyl Iron Complexes with Thiol, Phosphate, and Thiosulfate Ligands on Hemoglobin" International Journal of Molecular Sciences 25, no. 13: 7194. https://doi.org/10.3390/ijms25137194