Abstract
Breast cancer represents the most prevalent form of cancer and the leading cause of cancer-related mortality among females worldwide. It has been reported that several risk factors contribute to the appearance and progression of this disease. Despite the advancements in breast cancer treatment, a significant portion of patients with distant metastases still experiences no cure. The extracellular matrix represents a potential target for enhanced serum biomarkers in breast cancer. Furthermore, extracellular matrix degradation and epithelial–mesenchymal transition constitute the primary stages of local invasion during tumorigenesis. Additionally, the microbiome has a potential influence on diverse physiological processes. It is emerging that microbial dysbiosis is a significant element in the development and progression of various cancers, including breast cancer. Thus, a better understanding of extracellular matrix and microbiome interactions could provide novel alternatives to breast cancer treatment and management. In this review, we summarize the current evidence regarding the intricate relationship between breast cancer with the extracellular matrix and the microbiome. We discuss the arising associations and future perspectives in this field.
1. Introduction
Breast cancer (BC) is a severe, life-threatening condition widely recognized as a prevalent contributor to cancer mortality, being the most prevalent malignancy afflicting women [1]. Despite significant advancements in its diagnosis and treatment, BC metastasis continues to be a leading cause of mortality among women, causing an imperative need for a deeper understanding of the molecular and cellular mechanisms that empower cancer cells to not only survive but also expand [2]. Several risk factors have been described in the appearance and progression of this disease, including genetic factors (such as inherited mutations in genes), reproductive factors (e.g., age at menarche or age at first live birth), high breast density, dietary patterns, alcohol consumption, sedentary lifestyle, or obesity [3,4,5]. In this complex scenario, preclinical and clinical data have accumulated evidence indicating that some of these risk factors may worsen the incidence, severity, and mortality of many types of cancer, including BC, due to its relationship with adipose tissue-related chronic inflammation, immune cell dysregulation, angiogenesis, extracellular matrix (ECM) stiffness, and genomic instability [6].
Although conventional treatments such as chemotherapy, surgery, hormone therapy, or radiotherapy are commonly used, they often entail significant side effects. Therefore, there is a critical need to investigate more cost-effective and efficient treatment modalities for BC [7] like personalized medicine [8] based on genetic, phenotypic, and environmental factors, minimally invasive procedures [9] such as laparoscopic and robotic surgery, improved screening and early detection [10], cost-effective [11] drug development, combination therapies [12] using combinations of drugs that work synergistically, and telemedicine and remote monitoring [13].
A novel factor that has emerged in recent years and that could be linked with BC is the microbiome, which has been identified as a fundamental contributor to human health and overall wellbeing [14]. It can exert influence on a multitude of human biological, hormonal, and metabolic pathways. This influence can potentially stimulate the initiation, proliferation, and genetic instability of cancer within host cells or, alternatively, induce apoptosis [15]. The interplay between tumor cells and their microenvironment has been recognized as one of the key determinants of cancer development; data suggest that both the ECM and the microbiome represent microenvironments that contribute to the onset and progression of tumors [16]. ECM turnover represents a potential target for novel and enhanced serum biomarkers due to ECM homeostasis being meticulously regulated, as this process is disrupted in cancer, with a notable alteration in protein composition and elevated secretion of proteases [17]. The tumor microenvironment (which consists of cells, ECM, soluble factors, signaling molecules, and mechanical cues) supports tumor growth and invasion, providing protection from host immunity and niches for dormant metastases to thrive as well as fostering therapeutic resistance [18]. The ECM, as one constituent of the multifaceted tumor microenvironment, undergoes significant alterations during the progression of breast tumorigenesis since cancer cells can dysregulate ECM remodeling, facilitating irreversible proteolysis and crosslinking and influencing cell signaling, angiogenesis, and tissue biomechanics and hence exerting profound effects on disease advancement [19,20].
The present review summarizes the current evidence regarding the intricate relationship between BC, ECM, and the microbiome. It discusses the emergence of this association and its future prospects.
2. Breast Cancer
BC arises from the uncontrolled proliferation of breast epithelial cells triggered by diverse carcinogenic factors. In advanced stages, cancer cells can metastasize to distant sites, forming multi-organ lesions that directly jeopardize affected individuals’ lives [21]. BC represents the most prevalent form of cancer, surpassing lung cancer [22], and is the leading cause of cancer-related mortality among females in most countries except Australia/New Zealand, Northern Europe, Northern America, and China, where lung cancer deaths prevail, posing a public health challenge [23,24]. BC is characterized by diverse clinical manifestations among women and has become as a prevalent malignancy [25], constituting 31% of all cases and 15% of projected fatalities attributed to women’s cancer in 2023 [26]. Annually, 2.3 million new BC cases are reported [27] across all genders and age cohorts [28]. Although BC is commonly associated with women, men can also be affected, albeit with a lower frequency. Male BC comprises approximately 1% of all BC cases [29]. Regarding BC survival, significantly discrepant rates are evident on a global scale. These rates range from approximately 80% for developed nations to below 40% for developing countries. These data highlight substantial disparities in healthcare access and resources [30]. Early diagnosis of BC can cure 70–80% of patients. Nevertheless, the prognosis for metastatic BC is dismal, constituting over 90% of BC-related mortality [31]. Despite advancements in treatment modalities for primary BCs (including surgery, chemotherapy, and radiotherapy), a significant portion of patients with distant metastases still experiences a lack of cure [32]. Despite the majority of patients being diagnosed with early-stage BC, up to 30% of these cases experience relapse within the initial 10 years of follow-up [33].
Most breast malignancies are adenocarcinomas. In general, breast carcinoma is divided into ductal carcinoma in situ (DCIS; a noninvasive potentially malignant intraductal proliferation of epithelial cells, confined to the ducts and lobules) and invasive ductal carcinoma (IDC; the most common form of invasive BC) [34]. Breast tumors are categorized by histology and immunochemistry [2]. The transition from phenotypic to intrinsic molecular BC subtypes has made a paradigm shift in BC treatment. This shift highlights the importance of individualizing therapy according to tumor biomarker status in the evolving regimens [35]. Based on American Society of Clinical Oncology guidelines [36], estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) are the recommended tumor marker tests for the prevention, screening, and treatment of BC. Gene expression profiling of breast carcinoma also allows to primarily classify in at least four distinct “intrinsic” subtypes, including two ER-positive (luminal A and luminal B) and two ER-negative (ERBB2 and basal subtypes) groups, with different expression patterns and characteristics among them [37]. Furthermore, the study of markers of elevated proliferation, which generally indicate a poor outcome, is of significant interest. In this line, Ki-67 antigen is used to evaluate BC proliferative activity and is also a prognostic biomarker of the risk of recurrence [38].
Tumorigenesis disrupts the mammary gland equilibrium and its environment, reprograming fibroblasts into a pro-invasive phenotype, which promotes ECM remodeling and the collective invasion of cancer cells [39]. In primary BC, tumor cells that resist T-cell attack are quiescent, forming clusters with reduced immune infiltration that display superior tumorigenic capacity. Thus, they constitute immunotherapy-resistant reservoirs by orchestrating a local hypoxic immune-suppressive milieu that blocks T-cell function [40]. Furthermore, during tumorigenesis, vascular networks are formed (a process known as angiogenesis) to nourish and supply oxygen to BC cells. Elevated levels of angiogenic factors are symptomatic of the aggressive nature of the respective tumor cells and correlate with a relatively poor prognosis [41]. During BC development, metastasis may occur, and the local disease can spread to lymph nodes or distant organs. These metastatic BC cells acquire aggressive characteristics from the tumor microenvironment through several mechanisms, including epithelial–mesenchymal transition (i.e., epithelial cells losing their cell polarity and adhesion properties and transforming into mesenchymal stem cells with enhanced migratory and invasive capabilities [42]) and epigenetic regulation [43].
3. Extracellular Matrix
3.1. Structure, Composition, and Molecular Aspects
The ECM is an intricate network composed of a matrix of multidomain macromolecules organized in a specific manner to form a structurally stable composite. This contributes to tissues’ mechanical properties [44,45]. The content and proportion of ECM components (i.e., fibrillar collagens, fibronectin, specific laminins, proteoglycans, and matricellular proteins) dictate its compliance, stiffness, porosity, viscoelasticity, and biochemical properties [45,46]. Among ECM proteins, there are large numbers of ECM-modifying enzymes, ECM-binding growth factors, and other ECM-associated proteins (collagen subunits, proteoglycans, and glycoproteins) [47]. The collagen superfamily of proteins plays a dominant role in maintaining the tissues’ integrity. Most collagens form polymeric assemblies such as fibrils, networks, and filaments. Some of them, such as endostatin, inhibit angiogenesis and tumor growth [48]. Moreover, basement membranes are layered cell-adherent ECM components that form part of the tissue architecture, serving as an extension of the plasma membrane, protecting tissues from disruptive physical stresses, and providing an interactive interface between cell and the surrounding environment [49].
3.2. ECM in the Cancer and Metastasis Context
The ECM is a highly dynamic entity that plays an active role in tumor progression due to its constituent components and mechanical characteristics [50]. Notably, the collagen-rich and laminin-rich basement membrane, along with the basement membrane stroma, serve as the boundary for tumor cells that metastasize only upon successful penetration through this basement membrane barrier [46]. Thus, the ECM serves as a regulator of the stem cell niche, with its stiffness playing a pivotal role in modulating cell growth, survival, tissue-specific differentiation, and tissue homeostasis, as tumors typically exhibit increased fibrotic and stiff ECM environments [51]. As a result of this shift in ECM homeostasis, interstitial fibrillar type I and III collagen as well as basement membrane protein type IV collagen are particularly relevant [17].
ECM remodeling, known as desmoplasia, characterized by the formation of excessive connective tissue around invasive carcinoma primarily but not exclusively of the breast [52], is orchestrated by myofibroblasts, also known as “activated fibroblasts”, which generate an ECM abundant in fibronectin and collagen, resulting in a fibrotic and stiff ECM milieu [53]. ECM degradation and epithelial–mesenchymal transition constitute the primary stages of local invasion, facilitated by the up-regulation of various proteinases, with a complex interplay of different signaling pathways [54,55]. Furthermore, endothelial cells are assembled into a laminin-rich basement membrane matrix, which is degraded during angiogenesis. As angiogenesis proceeds, the ECM serves essential functions in supporting key signaling events promoting endothelial cells’ migration, invasion, proliferation, and survival [56]. Hence, cancer cells can degrade the ECM and can proliferate and infiltrate surrounding tissues, a pivotal step in the dissemination of the primary tumor and the formation of metastases [57].
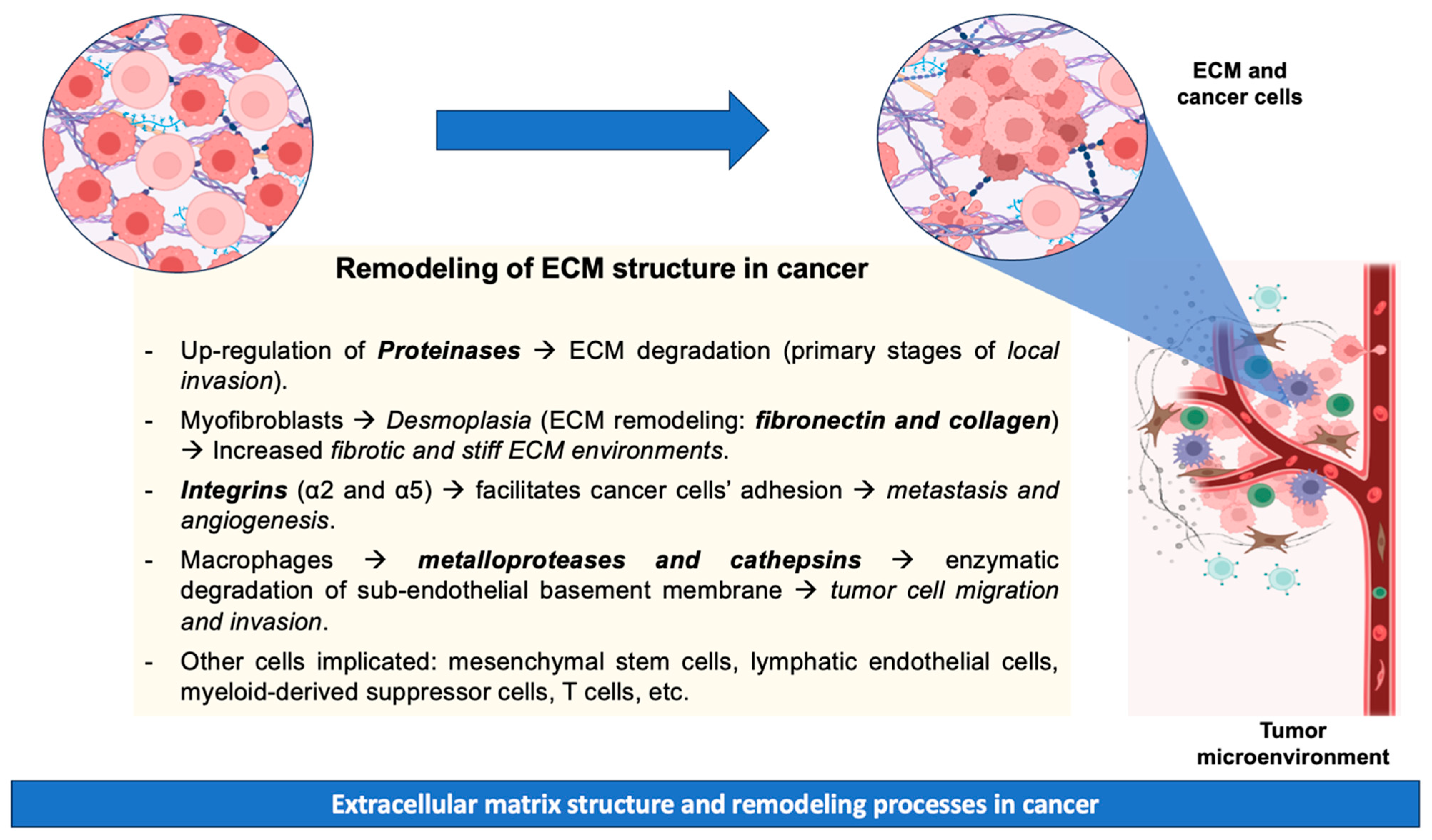
Metastatic disease results from the dissemination of metastatic tumor cells from the site of primary origin to distant organs. The tumor microenvironment is crucial to cancer progression, with various cell types participating in this process. These cell types include mesenchymal stem cells, lymphatic endothelial cells, cancer-associated fibroblasts, myeloid-derived suppressor cells, T cells, and tumor-associated macrophages [58]. The progression of metastatic carcinoma necessitates the initial penetration of the basement membrane, as neoplastic cells are compelled to enzymatically degrade the vascular sub-endothelial basement membrane [59]. During tissue remodeling, macrophages are recognized for their production of proteases (matrix metalloproteases and cathepsins) that facilitate the ECM breakdown, tumor cell migration, and invasion [60]. Additionally, cell adhesion to the ECM plays a pivotal role in regulating cellular processes (e.g., growth, differentiation, and apoptosis). However, cell–ECM adhesion mediated by integrins (specifically integrins α2 and α5) has been identified as a critical factor facilitating cancer cells’ adhesion, promoting metastasis and angiogenesis [61]. However, precise mechanisms related to cancer invasion and metastasis remain unknown due to their complexity [62]. The above-mentioned remodeling processes of ECM during cancer are summarized in Figure 1.
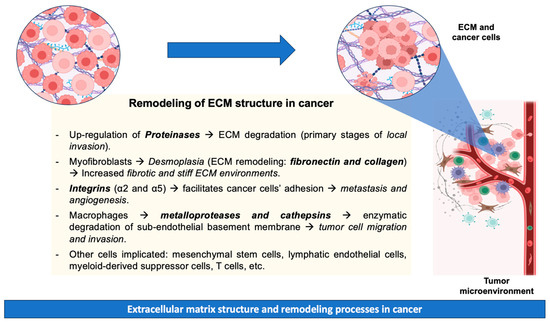
Figure 1.
Remodeling processes of ECM during cancer.
3.3. ECM-Targeting Therapies
ECM-targeting therapies have shown promise in various clinical applications, particularly in treating diseases where ECM dysregulation plays a significant role. For example, monoclonal antibodies [63] targeting ECM proteins (designed to interfere with the interactions between cancer cells and the ECM), small-molecule inhibitors of ECM-modifying enzymes [64] (target enzymes involved in the modification and turnover of ECM), matrix metalloproteinase (MMP) inhibitors [65] (which block their critical roles in the degradation and remodeling of the ECM), biological therapies targeting ECM-interacting proteins [66] (which involve using proteins, peptides, or antibodies to modulate interactions between cells and the ECM), and ECM-disrupting peptides [67] (short chains of amino acids designed to interfere with specific interactions between cells and the ECM).
3.4. Evidence Regarding ECM and BC
The ECM also plays a crucial role in normal breast function, such as mammary gland remodeling before and after lactation; its composition varies notably between metastatic and non-metastatic breast tumor stages [68]. Breast tumors are defined as epithelial neoplastic lesions distinguished by their dense structure, which predominantly comprises stromal cells and abundant ECM molecules. The reciprocal interactions between these components and epithelial cells are decisive in cancer progression [69]. In fact, BC progression frequently coincides with alterations in ECM stiffness and cellular adhesion capability, both intricately linked to cellular mechanotransduction [70]. Analyses of human BC samples have shown that tumor stiffness correlates with a more aggressive disease phenotype, as the progression is accompanied by substantial collagen deposition, linearization, and bundling [71]. Furthermore, the ECM plays a pivotal role in gene expression, cellular differentiation, and tissue organization during homeostasis. Hence, these factors have an essential role in the cellular invasion, metastasis, and drug responsiveness of BC cells [72].
During BC development, the ECM governs numerous pathways within cancer cells, including Wnt [73] (proteins that regulate cell-to-cell interactions as cell fate determination, cell proliferation, cell migration, and cell polarity), PI3K/AKT [74] (a critical intracellular signaling pathway that regulates various cellular processes, including metabolism, growth, proliferation, survival, and angiogenesis), ERK [75] (a series of phosphorylation events that culminate in the activation of ERK, whose dysregulation may lead to cancer, as it is a crucial intracellular signaling cascade), JNK [76] (involved in regulating inflammation, apoptosis, and stress responses that may be affected by stress stimuli, leading to the phosphorylation of target proteins and cancer), Src-FAK [77] (mediates cellular responses to extracellular signals, particularly those involving integrin-mediated adhesion and migration; Src kinase phosphorylates FAK at specific tyrosine residues, leading to the activation of downstream signaling pathways that regulate cytoskeletal dynamics, cell adhesion turnover, and cell motility), and Rho-GTPases [78] (essential for cellular functions such as the formation of stress fibers, regulation of cell polarity, and modulation of cell motility through their effects on actin cytoskeleton dynamics). Furthermore, the heightened deposition and crosslinking of collagens linked to tumorigenesis contributes to increased tissue stiffness [79]. Additionally, matrix metalloproteinases, which can enzymatically degrade diverse constituents of the ECM, and their functional genetic polymorphisms have been posited to potentially correlate with the BC risk [80]. Likewise, dysregulation in lysyl oxidase expression has been associated with the onset and progression of BC. This is due to the determinant role of this enzyme in the formation, maintenance, and functional characteristics of the ECM [81]. Another prominent ECM component within the stroma is hyaluronan (which belongs to the glycosaminoglycan family of polysaccharides), which play a significant role in the promotion of inflammation induced by BC [82].
Of significant importance, different inhibitors targeting various ECM components (e.g., integrins, proteoglycans, metalloproteinases, collagens, and the C1-peptidase protein families) have shown promising pre-clinical results in BC [68]. For instance, the inhibition of cathepsin B (namely the identified mutated CTSB gene in BC mice models) has been associated with reduced bone metastasis [83]. Likewise, the transforming growth factor β induced (TGFBI), which is released into the ECM (functioning mainly in cell adhesion, migration, proliferation, apoptosis, and angiogenesis), plays a suppressive role in the development of mesothelioma and BC cells, possibly through inhibitions of cell proliferation and inducing senescence [84]. The extracellular matrix protein 1 has also been proposed as a potential therapeutic target for overcoming tumor dissemination in BC metastasis. This is because of its apparent role in remodeling the actin cytoskeleton and in invadopodia (actin-based cortical protrusions of tumor cells required for stromal invasion and metastasis)[85].
4. Microbiome
4.1. The Human Microbiome
The human microbiome represents a complex and intricate ecological network, characterized by a diverse array of microorganisms (i.e., bacteria, viruses, fungi, and protozoa) that have established colonization at various anatomical sites throughout human bodies, such as the skin, oral cavity, vagina, or the gastrointestinal tract [86]. The microbiome inhabits and interacts with the human body. These interactions may be commensalistic, mutualistic, or pathogenic [87,88,89]. Furthermore, in recent years, the presence of microorganisms in organs or tissues has traditionally been considered as “sterile” [90]. These microbes have garnered increasing attention for their potential influence on diverse physiological processes (e.g., immune modulation, metabolism, and inflammation), and their role is emerging as a promising avenue for reshaping cancer development and therapy [91]. In this line, microbial dysbiosis has emerged as a crucial element in the development and progression of various cancers, including BC. A dysbiosis is defined as a derangement in bacterial abundance as a result of a decline in the richness and diversity of bacteria.
4.2. Microbiome and BC
Research evidence indicates a close relationship between microbial dysbiosis and BC [92,93]. BC manifests a notably diverse microbiome compared to normal breast samples and other forms of cancer. This diversity is marked by discernible variations across racial demographics, cancer stages, and BC subtypes [94,95]. These differences further exhibit variations based on clinicopathologic characteristics such as ER and PR status, levels of Ki-67, or HER2 status [96]. The characterization of the tumor microbiome remains challenging due to low biomass. Whether the presence of bacteria is advantageous to the tumors or to the bacteria themselves is unknown. For example, some BC subtypes characterized by increased oxidative stress are also enriched in bacteria that produce mycothiol, which detoxifies reactive oxygen species [97]. Thus, a thorough understanding of the breast microbiome and the impact of host genetics, lifestyle, and socioeconomic factors could be of significant interest in BC. The presence of some species has been found to be consistent across various studies, such as Prevotella and Micrococcus in normal breast tissue and Lactobacillus and Fusobacterium in malignant tumors of the breast [98,99,100,101]. In this line, it has been reported that bacterial composition differs from BC (where Ralstonia is abundant) to normal-breast individuals (highlighting the presence of Acetobacter aceti, Lactobacillus vini, Lactobacillus paracasei, and Xanthomonas spp.) [102]. Additionally, sequencing using breast tumor tissue and paired normal adjacent tissue from the same patient has shown that the bacterium Methylobacterium radiotolerans was relatively enriched in tumor tissue, while Sphingomonas yanoikuyae was enriched in the paired normal tissue, observing lower levels of antibacterial-response gene expression in tumor tissue [99]. Various processes, including the modulation of the immune response in BC, may be affected by the microbiome in breast tumors.
4.3. Oral Microbiome and BC
Recent research has underscored the significance of the oral microbiome, recognized as the most diverse microbiome within the human body, in the pathogenesis of various malignancies [103]. The current body of literature indicates a potential association between the oral microbiome and BC. Specifically, studies have demonstrated differences in oral microbial communities between BC patients and healthy women. BC risk is increased in women with periodontal disease. This is caused by bacteria such as Porphyromonas gingivalis, Tannerella forsythia, and Treponema denticola in the red complex. A number of other bacteria (Prevotella intermedia, Fusobacterium nucleatum, Prevotella nigrescens, Streptococcus constellatus, Peptostreptococcus micros, Campylobacter showae, Eubacterium nodatum, Campylobacter gracilis, and Campylobacter rectus) associated with periodontal disease have been identified in the orange complex [101,104,105]. BC cases showed lower relative abundances of Porphyromonas and Fusobacterium than controls. There is a strong correlation between alpha diversity and presence/relative abundance of specific genes from the oral and fecal microbiome among BC cases but not among controls. Among patients with BC, the relative abundance of oral Porphyromonas was inversely correlated with the relative abundance of fecal Bacteroides [106].
Moreover, one study identified a potential link between the oral microbiome, particularly that associated with menopausal and menstrual status, and BC risk [107]. This emerging association carries significant implications for cancer diagnostics, therapeutics, and prevention strategies. Consequently, comprehending the intricate interplay between the oral microbiome and carcinogenic pathways has emerged as a frontier in oncological research [108].
4.4. Gut Microbiome and BC
The importance of the gut microbiome to human health is increasingly recognized [109]. Gut dysbiosis has the potential to undermine gut barrier integrity, precipitating bacterial translocation and the subsequent establishment of a chronic inflammatory milieu within the gut. However, it remains unclear whether cancer pathology induces alterations in the gut microbiome or if dysbiosis itself serves as a carcinogenic factor [110]. Moreover, it is well established that chronic inflammation correlates with cancer growth and metastatic dissemination to distant organs [111]. In this regard, intracellular bacteria have been observed to accompany cancer cells within the circulatory system. This exerts a pivotal influence on tumor metastasis and colonization processes [112]. Obesity and a high-fat diet, particularly in cases of sporadic BC, may also be associated with the microbial communities residing within the host, potentially serving as a factor in breast carcinogenesis [113].
According to a cohort of 31 women with early BC, the abundance of Blautia species progressively increased with tumor grade, from 1.25 (1.01–1.43) in grade I to 2.95 (2.00–3.60) in grade III [114]. In patients with advanced clinical stages, Bacteroidota, Clostridium coccoides, and Clostridium leptum clusters were identified as well as Faecalibacterium prausnitzii and Blautia species [114].
Around 50% of breast carcinomas are associated with dietary factors. It is hypothesized that dietary fiber, by altering specific enzymatic activities such as β-glucuronidase, might influence the composition of the gut microbiome and estradiol metabolism, particularly among postmenopausal BC patients [115]. Furthermore, it has been reported that a reduction in gut bacterial diversity may contribute to estrogen release, ultimately resulting in an elevated BC risk [116]. Short-chain fatty acids such as butyrate can be produced in the gut microbiome through food digestion. These compounds possess enzyme-silencing properties that could potentially contribute to a decrease in BC development [117]. Additionally, low diversity in the gut microbiome has been correlated with diminished lymphocyte levels and elevated neutrophil counts, along with decreased survival rates among BC patients [118].
Given the gut microbiome’s capacity to influence estrogen metabolism and chronic inflammation linked to obesity, the composition of both the mammary and gut microbiome affects the risk of BC [119]. It has also been observed that restoring gut microbial homeostasis represents an emerging therapeutic strategy in BC treatment. Certain probiotics have demonstrated potential for preventing or treating BC by modulating gastrointestinal bacteria and the systemic immune system [120]. Another crucial element to consider is the role of phytoestrogens, which are metabolized by the gut microbiome and exert their effects through interaction with ERs, particularly ERβ. This interaction modulates target tissue responses. Although phytoestrogens typically exhibit lower receptor binding affinity than endogenous estrogen, their biological effects are contingent upon circulating estrogen levels, leading to context-dependent estrogenic or antiestrogenic actions [121]. Moreover, the gut microbiome influences chemotherapy efficacy and toxicity, thereby affecting treatment outcomes [122].
It must be noted that the gut microbiome is not exclusively shaped by dietary intake but also influenced by medications; certain studies indicate correlations between the microbiome composition and the response to chemotherapy, radiotherapy, and immunotherapy. Hence, interventions targeting microbiome manipulation could potentially be developed to prevent or treat BCs, optimizing therapeutic outcomes and mitigating adverse effects [123].
5. Relationship between BC, ECM, and the Microbiome
As part of its role in health maintenance, the ECM controls the diffusion of infections and inflammations, detects and adapts to external stimuli from the environment, and, in the case of cancer, interacts with cancer cells and controls tumor invasion and drug resistance [124]. Around BC masses, an accumulation of type I collagen fibrils, i.e., desmoplasia, occurs in order to resist cancer invasion mechanically [124]. Later, these same fibrils bundle to form fibers that run parallel to the surface of the tumor and prevent cancer cells from invading and spreading [124]. Cancer niche formation, tumor progression, and drug resistance are associated with positive interactions between tumor cells and the microbiome. In addition, several enzymes have been identified that are capable of degrading the host ECM (e.g., collagenase, hyaluronidase (HAase) and elastase) [125,126].
Hyaluronic acid is a major matrix molecule expressed in human malignancies [127]. There is a significant overexpression of it in BC (approximately 56%) [128]. This results in an increased biophysical barrier that significantly compresses blood vessels, increases interstitial fluid pressure, and hinders drug delivery [129,130]. Inflammation and cancer progression are facilitated by the binding of hyaluronic acid to the transmembrane receptor CD44, which is necessary for cell migration, proliferation, and invasion [131,132]. In clinical applications, especially oncology, hyaluronidase facilitates the dispersion and absorption of interstitial fluids. HAase protein derived from Staphylococcus aureus ATCC 29213 exhibits higher enzyme activity than that derived from other strains [133]. By degrading HA in the ECM, HAase reduces the interstitial fluid pressure in the tumor ECM. This enhances chemotherapy by activating interstitial space dispersion [134].
As previously mentioned, activated fibroblasts or myofibroblasts are responsible for the synthesis and remodeling of the ECM, which provides a path for tumor cells to exit the primary tissue and disseminate throughout the body [135]. Commensal dysbiosis increases the frequency of mast cells in the mammary tissue as well as their profibrogenicity, a phenotypic change that persists after tumor implantation [136]. The activation of fibroblasts and remodeling of tissue are associated with enhanced breast tumor metastasis. Through pharmacological and adoptive transfer approaches, Feng et al. were able to demonstrate that mast cells from dysbiotic animals increase the dissemination of hormone receptor-positive tumor cells in mammary tissue. Mammary collagen levels were correlated with mast cell abundance in hormone receptor-positive BC patients, suggesting that mast cell-mediated fibroblast activation may have clinical relevance. In conjunction, these data indicate that fibroblast activation and early tumor dissemination are mediated by a gut–mast cell axis [136].
Invasion of BC requires the stiffening and self-assembly of the matrix made up of collagen I and ECM modifiers [137]. A pathologic ECM promotes BC cell proliferation by activating the FAK and ERK pathways [138]. In BC, the balance between intracellular and cell–matrix adhesions determines the ECM dynamics and the invasion of cancerous cells [139]. Inflammation and remodeling of ECM have been reported in association with Anaerococcus [140]. A key role may be played by the interaction between the ECM, the microbiome, and inflammation in the onset, progression, and relapse of BC [141]. In general, BC patients showed an increase in bacterial richness [142]. According to Zeng et al. [143], patients with low alpha diversity (variation of microbes in a single sample) have a significantly longer recurrence-free survival than those with high alpha diversity [143].
6. Further Perspectives
BC molecular subtyping is crucial for predictive and prognostic purposes due to diverse clinical behaviors observed across types. Recently, different alternatives and novel biomarkers of BC identification have emerged that may improve clinical outcomes and provide tailored treatments.
In this context, radiogenomics has the potential to significantly improve early detection, prognosis, and diagnosis of BC since it allows for visual features and genetic marker linkage that promise to eliminate the need for biopsy and sequencing [144]. Similarly, liquid biopsy using ctDNA, which has been reported to be less invasive and effective for comprehensive genetic analysis of heterogeneous solid tumors, could be useful across a wide range of cancer types, including BC. Its clinical applications are expected to expand further through ongoing research [145]. Likewise, the application of circRNAs in clinical practice for diagnosing, treating, and monitoring BC could be another promising alternative. Nevertheless, the interaction and influence of multiple pathways must be investigated, requiring more in-depth studies to improve the understanding of the molecular mechanism of the circRNA network [146]. Additionally, it has been highlighted that the potential diagnostic function of lncRNA H-19 and miR-200a in BC as well as the association of IL-6/SIRT-1 with lncRNA H-19/miR-200a expression could be a promising opportunity for clinical outcomes and tailored treatments [147]. Lastly, the levels of advanced oxidation protein products appear to have a significant role in predicting cancer-related events and may potentially serve as a simple prognostic marker in clinical practice [148].
Several treatment options for BC are available today, including surgery, chemotherapy, radiotherapy and immunotherapy, the last being a highly specific therapy involving adaptive immune responses and immunological memory. In this field, research has proposed the creation of cancer vaccines and immunotherapies for BC malignancies, targeting proteins such as the GRP78 protein [149]. Immunotherapy appears to have widely different results, with some patients showing a pronounced clinical response, while others experience minimal or no clinical benefit from the same treatment. Hence, understanding the complexity and diversity of the immune context of the tumor microenvironment could predict and guide the immunotherapeutic response [150]. On the other hand, bacterial therapeutics for tumor treatment and immune modulation are another promising alternative. This is due to the significance of the microbiome in mitigating BC via its anti-tumor activities [151]. Microbiome composition categorizes BC patients into subgroups, thereby facilitating tailored treatment approaches with enhanced efficacy. The integration of cell lines and circulating tumor cells with advanced analytical techniques unveils intricate mechanisms [152]. However, bacteriotherapy, aimed at reversing gut dysbiosis and enhancing diagnostic outcomes, is not without challenges [151].
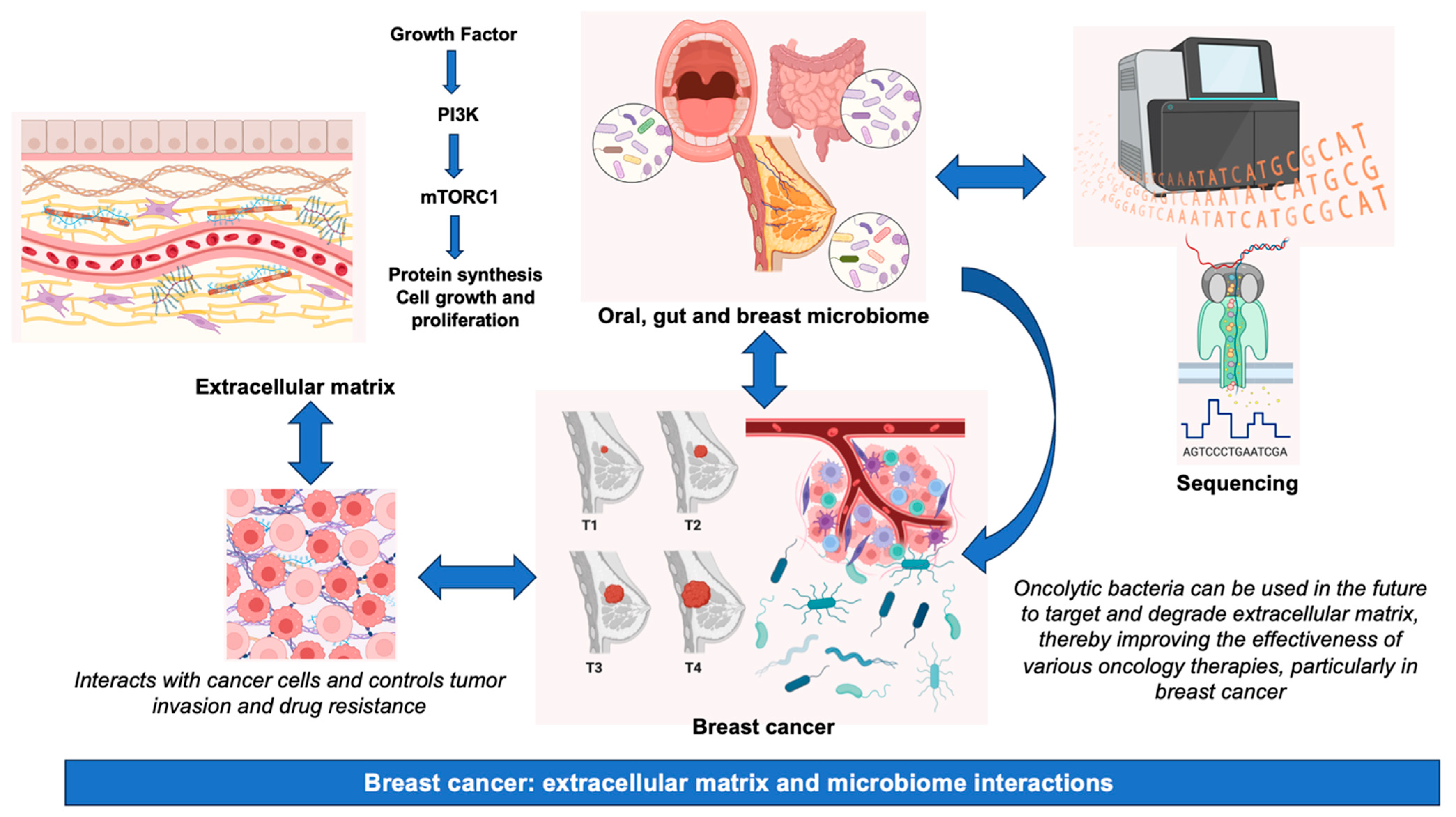
The results of studies in tumor mouse models of BC (4T1) showed that HAase-mediated ECM degradation reduces interstitial fluid pressure in the tumor microenvironment, suggesting that this strategy may overcome the limitations of chemotherapy. By using oncolytic bacteria to target and degrade ECM, drug penetration management will be facilitated in the future, thereby improving the outcomes of various oncology therapies, particularly in BC [134].
In summary, although the significance of the microbiome and other therapeutic approaches in BC pathogenesis is garnering escalating attention, it is imperative to acknowledge the inherent challenges, which encompass, among others, disparities in sample collection techniques, variations in DNA extraction methodologies, potential issues of contamination, and the necessity for rigorous bioinformatics and statistical analyses to interpret intricate datasets [153]. A summary of the main information presented in this review is graphically represented in Figure 2.
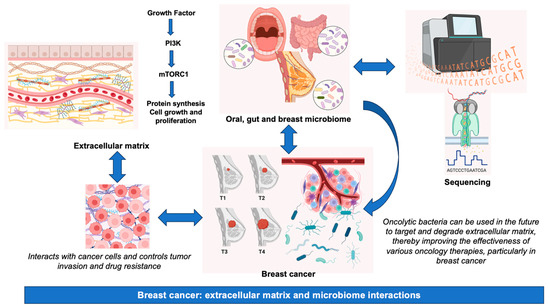
Figure 2.
Breast cancer: extracellular matrix and microbiome interactions. Abbreviations, mTORC1, mammalian target of rapamycin complex 1; PI3K, phosphatidylinositol-3 kinase.
Author Contributions
Conceptualization, L.H.-Q. and J.P.-D.; methodology, L.H.-Q. and H.V.-L.; software, J.P.-D.; validation, J.P.-D.; writing—original draft preparation, L.H.-Q. and H.V.-L.; writing—review and editing, J.P.-D.; visualization, J.P.-D.; supervision J.P.-D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ranaee, M.; Torabi, H.; Azhganzad, N.; Shirini, K.; Hosseini, A.S.; Hajian, K. The Relationship Between Tumor Budding and Patient’s Survival in Breast Cancer. Clin. Pathol. 2024, 17, 2632010X241235543. [Google Scholar] [CrossRef]
- Grasset, E.M.; Barille-Nion, S.; Juin, P.P. Stress in the metastatic journey—The role of cell communication and clustering in breast cancer progression and treatment resistance. Dis. Models Mech. 2024, 17, dmm050542. [Google Scholar] [CrossRef]
- Pfeiffer, R.M.; Webb-Vargas, Y.; Wheeler, W.; Gail, M.H. Proportion of U.S. Trends in Breast Cancer Incidence Attributable to Long-term Changes in Risk Factor Distributions. Cancer Epidemiol. Biomark. Prev. 2018, 27, 1214–1222. [Google Scholar] [CrossRef]
- Zamzam, S.; Said, S.; Yaghi, J.; Faisal, F.S.; Hassan, D.; Abdul Majeed, S.; Al Rajabi, A.; Tayyem, R. Dietary Patterns Associated with Breast Cancer in the Middle East: A Scoping Review. Nutrients 2024, 16, 579. [Google Scholar] [CrossRef]
- Nguyen, M.R.; Ma, E.; Wyatt, D.; Knight, K.L.; Osipo, C. The effect of an exopolysaccharide probiotic molecule from Bacillus subtilis on breast cancer cells. Front. Oncol. 2023, 13, 1292635. [Google Scholar] [CrossRef]
- Reggiani, F.; Bertolini, F. Roles of obesity in the development and progression of breast cancer. Discov. Med. 2017, 24, 183–190. [Google Scholar]
- Mohan Ram Kumar, R.; Rajan, L.; Joghee, S. Breast cancer derived exosomes: Theragnostic perspectives and implications. Clin. Chim. Acta 2024, 557, 117875. [Google Scholar] [CrossRef]
- Ashley, E.A. The precision medicine initiative: A new national effort. JAMA 2015, 313, 2119–2120. [Google Scholar] [CrossRef]
- Krag, D.N.; Anderson, S.J.; Julian, T.B.; Brown, A.M.; Harlow, S.P.; Costantino, J.P.; Ashikaga, T.; Weaver, D.L.; Mamounas, E.P.; Jalovec, L.M.; et al. Sentinel-lymph-node resection compared with conventional axillary-lymph-node dissection in clinically node-negative patients with breast cancer: Overall survival findings from the NSABP B-32 randomised phase 3 trial. Lancet Oncol. 2010, 11, 927–933. [Google Scholar] [CrossRef]
- Tabar, L.; Yen, M.F.; Vitak, B.; Chen, H.H.; Smith, R.A.; Duffy, S.W. Mammography service screening and mortality in breast cancer patients: 20-year follow-up before and after introduction of screening. Lancet 2003, 361, 1405–1410. [Google Scholar] [CrossRef]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar] [CrossRef]
- Bardia, A.; Mayer, I.A.; Vahdat, L.T.; Tolaney, S.M.; Isakoff, S.J.; Diamond, J.R.; O’Shaughnessy, J.; Moroose, R.L.; Santin, A.D.; Abramson, V.G.; et al. Sacituzumab Govitecan-hziy in Refractory Metastatic Triple-Negative Breast Cancer. N. Engl. J. Med. 2019, 380, 741–751. [Google Scholar] [CrossRef]
- Vidal-Alaball, J.; Acosta-Roja, R.; Pastor Hernandez, N.; Sanchez Luque, U.; Morrison, D.; Narejos Perez, S.; Perez-Llano, J.; Salvador Verges, A.; Lopez Segui, F. Telemedicine in the face of the COVID-19 pandemic. Aten. Primaria 2020, 52, 418–422. [Google Scholar] [CrossRef]
- Luo, L.; Chen, Y.; Ma, Q.; Huang, Y.; Xu, L.; Shu, K.; Zhang, Z.; Liu, Z. Ginger volatile oil inhibits the growth of MDA-MB-231 in the bisphenol A environment by altering gut microbial diversity. Heliyon 2024, 10, e24388. [Google Scholar] [CrossRef]
- Roy, S.; Trinchieri, G. Microbiota: A key orchestrator of cancer therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef]
- Alfano, M.; Canducci, F.; Nebuloni, M.; Clementi, M.; Montorsi, F.; Salonia, A. The interplay of extracellular matrix and microbiome in urothelial bladder cancer. Nat. Rev. Urol. 2016, 13, 77–90. [Google Scholar] [CrossRef]
- Bager, C.L.; Willumsen, N.; Leeming, D.J.; Smith, V.; Karsdal, M.A.; Dornan, D.; Bay-Jensen, A.C. Collagen degradation products measured in serum can separate ovarian and breast cancer patients from healthy controls: A preliminary study. Cancer Biomark. 2015, 15, 783–788. [Google Scholar] [CrossRef]
- Swartz, M.A.; Iida, N.; Roberts, E.W.; Sangaletti, S.; Wong, M.H.; Yull, F.E.; Coussens, L.M.; DeClerck, Y.A. Tumor microenvironment complexity: Emerging roles in cancer therapy. Cancer Res. 2012, 72, 2473–2480. [Google Scholar] [CrossRef]
- Mohan, V.; Das, A.; Sagi, I. Emerging roles of ECM remodeling processes in cancer. Semin. Cancer Biol. 2020, 62, 192–200. [Google Scholar] [CrossRef]
- Lambert, A.W.; Ozturk, S.; Thiagalingam, S. Integrin signaling in mammary epithelial cells and breast cancer. ISRN Oncol. 2012, 2012, 493283. [Google Scholar] [CrossRef]
- Luan, B.; Ge, F.; Lu, X.; Li, Z.; Zhang, H.; Wu, J.; Yang, Q.; Chen, L.; Zhang, W.; Chen, W. Changes in the fecal microbiota of breast cancer patients based on 16S rRNA gene sequencing: A systematic review and meta-analysis. Clin. Transl. Oncol. 2024, 26, 1480–1496. [Google Scholar] [CrossRef]
- Li, H.; Dong, T.; Tao, M.; Zhao, H.; Lan, T.; Yan, S.; Gong, X.; Hou, Q.; Ma, X.; Song, Y. Fucoidan enhances the anti-tumor effect of anti-PD-1 immunotherapy by regulating gut microbiota. Food Funct. 2024, 15, 3463–3478. [Google Scholar] [CrossRef]
- Chen, S.; Navickas, A.; Goodarzi, H. Translational adaptation in breast cancer metastasis and emerging therapeutic opportunities. Trends Pharmacol. Sci. 2024, 45, 304–318. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Barzaman, K.; Karami, J.; Zarei, Z.; Hosseinzadeh, A.; Kazemi, M.H.; Moradi-Kalbolandi, S.; Safari, E.; Farahmand, L. Breast cancer: Biology, biomarkers, and treatments. Int. Immunopharmacol. 2020, 84, 106535. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Kim, M.C.; Cho, E.Y.; Park, S.Y.; Lee, H.J.; Lee, J.S.; Kim, J.Y.; Lee, H.C.; Yoo, J.Y.; Kim, H.S.; Kim, B.; et al. A Nationwide Study on HER2-low Breast Cancer in South Korea: Its Incidence of 2022 Real World Data and the Importance of Immunohistochemical Staining Protocols. Cancer Res. Treat. 2024. [Google Scholar] [CrossRef]
- Goh, S.P.; Ong, S.C.; Chan, J.E. Economic evaluation of germline genetic testing for breast cancer in low- and middle-income countries: A systematic review. BMC Cancer 2024, 24, 316. [Google Scholar] [CrossRef]
- Ray, S.K.; Mukherjee, S. Breast cancer stem cells as novel biomarkers. Clin. Chim. Acta 2024, 557, 117855. [Google Scholar] [CrossRef]
- Akram, M.; Iqbal, M.; Daniyal, M.; Khan, A.U. Awareness and current knowledge of breast cancer. Biol. Res. 2017, 50, 33. [Google Scholar] [CrossRef]
- Tsubaki, M.; Genno, S.; Takeda, T.; Matsuda, T.; Kimura, N.; Yamashita, Y.; Morii, Y.; Shimomura, K.; Nishida, S. Rhosin Suppressed Tumor Cell Metastasis through Inhibition of Rho/YAP Pathway and Expression of RHAMM and CXCR4 in Melanoma and Breast Cancer Cells. Biomedicines 2021, 9, 35. [Google Scholar] [CrossRef]
- Sun, H.F.; Yang, X.L.; Zhao, Y.; Tian, Q.; Chen, M.T.; Zhao, Y.Y.; Jin, W. Loss of TMEM126A promotes extracellular matrix remodeling, epithelial-to-mesenchymal transition, and breast cancer metastasis by regulating mitochondrial retrograde signaling. Cancer Lett. 2019, 440–441, 189–201. [Google Scholar] [CrossRef]
- Garcia-Torralba, E.; Perez Ramos, M.; Ivars Rubio, A.; Navarro Manzano, E.; Blaya Boluda, N.; Lloret Gil, M.; Aller, A.; de la Morena Barrio, P.; Garcia Garre, E.; Martinez Diaz, F.; et al. Deconstructing neutrophil to lymphocyte ratio (NLR) in early breast cancer: Lack of prognostic utility and biological correlates across tumor subtypes. Breast Cancer Res. Treat. 2024, 205, 475–485. [Google Scholar] [CrossRef]
- Makki, J. Diversity of Breast Carcinoma: Histological Subtypes and Clinical Relevance. Clin. Med. Insights Pathol. 2015, 8, 23–31. [Google Scholar] [CrossRef]
- Hashmi, A.A.; Hashmi, K.A.; Irfan, M.; Khan, S.M.; Edhi, M.M.; Ali, J.P.; Hashmi, S.K.; Asif, H.; Faridi, N.; Khan, A. Ki67 index in intrinsic breast cancer subtypes and its association with prognostic parameters. BMC Res. Notes 2019, 12, 605. [Google Scholar] [CrossRef]
- Harris, L.; Fritsche, H.; Mennel, R.; Norton, L.; Ravdin, P.; Taube, S.; Somerfield, M.R.; Hayes, D.F.; Bast, R.C., Jr.; American Society of Clinical, O. American Society of Clinical Oncology 2007 update of recommendations for the use of tumor markers in breast cancer. J. Clin. Oncol. 2007, 25, 5287–5312. [Google Scholar] [CrossRef]
- Bombonati, A.; Sgroi, D.C. The molecular pathology of breast cancer progression. J. Pathol. 2011, 223, 307–317. [Google Scholar] [CrossRef]
- de Azambuja, E.; Cardoso, F.; de Castro, G., Jr.; Colozza, M.; Mano, M.S.; Durbecq, V.; Sotiriou, C.; Larsimont, D.; Piccart-Gebhart, M.J.; Paesmans, M. Ki-67 as prognostic marker in early breast cancer: A meta-analysis of published studies involving 12,155 patients. Br. J. Cancer 2007, 96, 1504–1513. [Google Scholar] [CrossRef]
- Albrengues, J.; Bertero, T.; Grasset, E.; Bonan, S.; Maiel, M.; Bourget, I.; Philippe, C.; Herraiz Serrano, C.; Benamar, S.; Croce, O.; et al. Epigenetic switch drives the conversion of fibroblasts into proinvasive cancer-associated fibroblasts. Nat. Commun. 2015, 6, 10204. [Google Scholar] [CrossRef]
- Baldominos, P.; Barbera-Mourelle, A.; Barreiro, O.; Huang, Y.; Wight, A.; Cho, J.W.; Zhao, X.; Estivill, G.; Adam, I.; Sanchez, X.; et al. Quiescent cancer cells resist T cell attack by forming an immunosuppressive niche. Cell 2022, 185, 1694–1708.e1619. [Google Scholar] [CrossRef]
- Madu, C.O.; Wang, S.; Madu, C.O.; Lu, Y. Angiogenesis in Breast Cancer Progression, Diagnosis, and Treatment. J. Cancer 2020, 11, 4474–4494. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Park, M.; Kim, D.; Ko, S.; Kim, A.; Mo, K.; Yoon, H. Breast Cancer Metastasis: Mechanisms and Therapeutic Implications. Int. J. Mol. Sci. 2022, 23, 6806. [Google Scholar] [CrossRef]
- Yue, B. Biology of the extracellular matrix: An overview. J. Glaucoma 2014, 23, S20–S23. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Mendez-Gutierrez, A.; Aguilera, C.M.; Plaza-Diaz, J. Extracellular Matrix Remodeling of Adipose Tissue in Obesity and Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef]
- Wu, C.; Dong, S.; Huang, R.; Chen, X. Cancer-Associated Adipocytes and Breast Cancer: Intertwining in the Tumor Microenvironment and Challenges for Cancer Therapy. Cancers 2023, 15, 726. [Google Scholar] [CrossRef]
- Hynes, R.O.; Naba, A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef]
- Myllyharju, J.; Kivirikko, K.I. Collagens and collagen-related diseases. Ann. Med. 2001, 33, 7–21. [Google Scholar] [CrossRef]
- Yurchenco, P.D. Basement membranes: Cell scaffoldings and signaling platforms. Cold Spring Harb. Perspect. Biol. 2011, 3, a004911. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, Z.; Pan, D.; Li, Y.; Zhou, J.; Chen, H.; Li, Z.; Zhu, M.; Li, C.; Qin, L.; et al. Dendritic Polymer-Based Nanomedicines Remodel the Tumor Stroma: Improve Drug Penetration and Enhance Antitumor Immune Response. Adv. Mater. 2024, 36, 2401304. [Google Scholar] [CrossRef]
- Northey, J.J.; Hayward, M.K.; Yui, Y.; Stashko, C.; Kai, F.; Mouw, J.K.; Thakar, D.; Lakins, J.N.; Ironside, A.J.; Samson, S.; et al. Mechanosensitive hormone signaling promotes mammary progenitor expansion and breast cancer risk. Cell Stem Cell 2024, 31, 106–126.e113. [Google Scholar] [CrossRef] [PubMed]
- DeClerck, Y.A. Desmoplasia: A response or a niche? Cancer Discov. 2012, 2, 772–774. [Google Scholar] [CrossRef]
- Holm, J.B.; Rosendahl, A.H.; Borgquist, S. Local Biomarkers Involved in the Interplay between Obesity and Breast Cancer. Cancers 2021, 13, 6286. [Google Scholar] [CrossRef]
- Zandi, Z.; Kashani, B.; Poursani, E.M.; Bashash, D.; Kabuli, M.; Momeny, M.; Mousavi-Pak, S.H.; Sheikhsaran, F.; Alimoghaddam, K.; Mousavi, S.A.; et al. TLR4 blockade using TAK-242 suppresses ovarian and breast cancer cells invasion through the inhibition of extracellular matrix degradation and epithelial-mesenchymal transition. Eur. J. Pharmacol. 2019, 853, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Anguita-Ruiz, A.; Bustos-Aibar, M.; Plaza-Diaz, J.; Mendez-Gutierrez, A.; Alcala-Fdez, J.; Aguilera, C.M.; Ruiz-Ojeda, F.J. Omics Approaches in Adipose Tissue and Skeletal Muscle Addressing the Role of Extracellular Matrix in Obesity and Metabolic Dysfunction. Int. J. Mol. Sci. 2021, 22, 2756. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.E.; Senger, D.R. Endothelial extracellular matrix: Biosynthesis, remodeling, and functions during vascular morphogenesis and neovessel stabilization. Circ. Res. 2005, 97, 1093–1107. [Google Scholar] [CrossRef]
- Xue, Y.; Li, M.; Hu, J.; Song, Y.; Guo, W.; Miao, C.; Ge, D.; Hou, Y.; Wang, X.; Huang, X.; et al. Ca(v)2.2-NFAT2-USP43 axis promotes invadopodia formation and breast cancer metastasis through cortactin stabilization. Cell Death Dis. 2022, 13, 812. [Google Scholar] [CrossRef]
- Riaz, F.; Zhang, J.; Pan, F. Forces at play: Exploring factors affecting the cancer metastasis. Front. Immunol. 2024, 15, 1274474. [Google Scholar] [CrossRef]
- Mader, C.C.; Oser, M.; Magalhaes, M.A.; Bravo-Cordero, J.J.; Condeelis, J.; Koleske, A.J.; Gil-Henn, H. An EGFR-Src-Arg-cortactin pathway mediates functional maturation of invadopodia and breast cancer cell invasion. Cancer Res. 2011, 71, 1730–1741. [Google Scholar] [CrossRef]
- Elfstrum, A.K.; Bapat, A.S.; Schwertfeger, K.L. Defining and targeting macrophage heterogeneity in the mammary gland and breast cancer. Cancer Med. 2024, 13, e7053. [Google Scholar] [CrossRef]
- Kim, E.S.; Kim, S.Y.; Koh, M.; Lee, H.M.; Kim, K.; Jung, J.; Kim, H.S.; Moon, W.K.; Hwang, S.; Moon, A. C-reactive protein binds to integrin alpha2 and Fcgamma receptor I, leading to breast cell adhesion and breast cancer progression. Oncogene 2018, 37, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Song, B.; Hong, S.; Kim, S.H.; Lee, K. Biomimetic 3D Clusters Using Human Adipose Derived Mesenchymal Stem Cells and Breast Cancer Cells: A Study on Migration and Invasion of Breast Cancer Cells. Mol. Pharm. 2016, 13, 2204–2213. [Google Scholar] [CrossRef] [PubMed]
- Polman, C.H.; O’Connor, P.W.; Havrdova, E.; Hutchinson, M.; Kappos, L.; Miller, D.H.; Phillips, J.T.; Lublin, F.D.; Giovannoni, G.; Wajgt, A.; et al. A randomized, placebo-controlled trial of natalizumab for relapsing multiple sclerosis. N. Engl. J. Med. 2006, 354, 899–910. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, I.F.; Chernichovsky, T.; Hagin, D.; Ingbir, M.; Reshef, R.; Chernin, G.; Levo, Y.; Schwartz, D. Differential regulation of L-arginine transporters (cationic amino acid transporter-1 and -2) by peroxynitrite in rat mesangial cells. Nephrol. Dial. Transplant. 2006, 21, 3409–3414. [Google Scholar] [CrossRef] [PubMed]
- Granda, T.G.; D’Attino, R.M.; Filipski, E.; Vrignaud, P.; Garufi, C.; Terzoli, E.; Bissery, M.C.; Levi, F. Circadian optimisation of irinotecan and oxaliplatin efficacy in mice with Glasgow osteosarcoma. Br. J. Cancer 2002, 86, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R.V. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J. Cell Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Apuzzo, M.L. Ad astra per aspera: Audacity and reinvention. Neurosurgery 2001, 49, 239. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, M.; Drigo, S.A.; Tonhosolo, R.; Andrade, S.C.S.; Marchi, F.A.; Jurisica, I.; Kowalski, L.P.; Achatz, M.I.; Rogatto, S.R. HABP2 p.G534E variant in patients with family history of thyroid and breast cancer. Oncotarget 2017, 8, 40896–40905. [Google Scholar] [CrossRef]
- Mohammed, H.; Carroll, J.S. Approaches for assessing and discovering protein interactions in cancer. Mol. Cancer Res. 2013, 11, 1295–1302. [Google Scholar] [CrossRef]
- Liang, P.; Wang, B. An autophagy-independent role of ULK1/ULK2 in mechanotransduction and breast cancer cell migration. Autophagy 2024, 20, 1199–1200. [Google Scholar] [CrossRef]
- Ponce, I.; Garrido, N.; Tobar, N.; Melo, F.; Smith, P.C.; Martinez, J. Matrix Stiffness Modulates Metabolic Interaction between Human Stromal and Breast Cancer Cells to Stimulate Epithelial Motility. Metabolites 2021, 11, 432. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.M.; Callaway, M.K.; Dos Santos, C.O. Clinical applications of 3D normal and breast cancer organoids: A review of concepts and methods. Exp. Biol. Med. 2022, 247, 2176–2183. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Hoxhaj, G.; Manning, B.D. The PI3K-AKT network at the interface of oncogenic signalling and cancer metabolism. Nat. Rev. Cancer 2020, 20, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J. Signal transduction by the JNK group of MAP kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef]
- Schaller, M.D. Cellular functions of FAK kinases: Insight into molecular mechanisms and novel functions. J. Cell Sci. 2010, 123, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell., 5th ed.; Garland Science: New York, USA, 2007; pp. 1–1392. [Google Scholar]
- Zhu, J.; Xiong, G.; Trinkle, C.; Xu, R. Integrated extracellular matrix signaling in mammary gland development and breast cancer progression. Histol. Histopathol. 2014, 29, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Du, L.F.; Lv, G.Q.; Yu, X.M.; Gu, Y.L.; Li, J.P.; Zhang, C. Current evidence on the relationship between four polymorphisms in the matrix metalloproteinases (MMP) gene and breast cancer risk: A meta-analysis. Breast Cancer Res. Treat. 2011, 127, 813–818. [Google Scholar] [CrossRef]
- Ferreira, S.; Saraiva, N.; Rijo, P.; Fernandes, A.S. LOXL2 Inhibitors and Breast Cancer Progression. Antioxidants 2021, 10, 312. [Google Scholar] [CrossRef]
- Schwertfeger, K.L.; Cowman, M.K.; Telmer, P.G.; Turley, E.A.; McCarthy, J.B. Hyaluronan, Inflammation, and Breast Cancer Progression. Front. Immunol. 2015, 6, 236. [Google Scholar] [CrossRef]
- Withana, N.P.; Blum, G.; Sameni, M.; Slaney, C.; Anbalagan, A.; Olive, M.B.; Bidwell, B.N.; Edgington, L.; Wang, L.; Moin, K.; et al. Cathepsin B inhibition limits bone metastasis in breast cancer. Cancer Res. 2012, 72, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wen, G.; Zhao, Y.; Tong, J.; Hei, T.K. The role of TGFBI in mesothelioma and breast cancer: Association with tumor suppression. BMC Cancer 2012, 12, 239. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, D.; Luo, Q.; Yang, Q.; Zhao, C.; Zhang, D.; Zeng, Y.; Huang, L.; Zhang, Z.; Qi, Z. Extracellular matrix protein 1 recruits moesin to facilitate invadopodia formation and breast cancer metastasis. Cancer Lett. 2018, 437, 44–55. [Google Scholar] [CrossRef]
- Feng, K.; Ren, F.; Xing, Z.; Zhao, Y.; Yang, C.; Liu, J.; Shang, Q.; Wang, X.; Wang, X. Microbiome and its implications in oncogenesis: A Mendelian randomization perspective. Am. J. Cancer Res. 2023, 13, 5785–5804. [Google Scholar] [PubMed]
- Ogunrinola, G.A.; Oyewale, J.O.; Oshamika, O.O.; Olasehinde, G.I. The Human Microbiome and Its Impacts on Health. Int. J. Microbiol. 2020, 2020, 8045646. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Alvarez-Mercado, A.I. The Interplay between Microbiota and Chemotherapy-Derived Metabolites in Breast Cancer. Metabolites 2023, 13, 703. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J. Nutrition, Microbiota and Noncommunicable Diseases. Nutrients 2020, 12, 1971. [Google Scholar] [CrossRef]
- Michan-Dona, A.; Vazquez-Borrego, M.C.; Michan, C. Are there any completely sterile organs or tissues in the human body? Is there any sacred place? Microb. Biotechnol. 2024, 17, e14442. [Google Scholar] [CrossRef]
- Liu, W.; Pi, Z.; Wang, X.; Shang, C.; Song, C.; Wang, R.; He, Z.; Zhang, X.; Wan, Y.; Mao, W. Microbiome and lung cancer: Carcinogenic mechanisms, early cancer diagnosis, and promising microbial therapies. Crit. Rev. Oncol. Hematol. 2024, 196, 104322. [Google Scholar] [CrossRef]
- Ye, C.; Liu, X.; Liu, Z.; Pan, C.; Zhang, X.; Zhao, Z.; Sun, H. Fusobacterium nucleatum in tumors: From tumorigenesis to tumor metastasis and tumor resistance. Cancer Biol. Ther. 2024, 25, 2306676. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Alvarez-Mercado, A.I.; Ruiz-Marin, C.M.; Reina-Perez, I.; Perez-Alonso, A.J.; Sanchez-Andujar, M.B.; Torne, P.; Gallart-Aragon, T.; Sanchez-Barron, M.T.; Reyes Lartategui, S.; et al. Association of breast and gut microbiota dysbiosis and the risk of breast cancer: A case-control clinical study. BMC Cancer 2019, 19, 495. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Cai, Y.; Yao, P.; Jia, Y.; Wei, X.; Du, C.; Zhang, S. Multi-omics analysis elucidates the relationship between intratumor microbiome and host immune heterogeneity in breast cancer. Microbiol. Spectr. 2024, 12, e0410423. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.F.; Reina-Perez, I.; Astorga, J.M.; Rodriguez-Carrillo, A.; Plaza-Diaz, J.; Fontana, L. Breast Cancer and Its Relationship with the Microbiota. Int. J. Environ. Res. Public Health 2018, 15, 1747. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.M.; Tran, H.T.T.; Long, J.; Shrubsole, M.J.; Cai, H.; Yang, Y.; Nguyen, L.M.; Nguyen, G.H.; Nguyen, C.V.; Ta, T.V.; et al. Gut Microbiome of Patients With Breast Cancer in Vietnam. JCO Glob. Oncol. 2024, 10, e2300234. [Google Scholar] [CrossRef] [PubMed]
- Nejman, D.; Livyatan, I.; Fuks, G.; Gavert, N.; Zwang, Y.; Geller, L.T.; Rotter-Maskowitz, A.; Weiser, R.; Mallel, G.; Gigi, E.; et al. The human tumor microbiome is composed of tumor type-specific intracellular bacteria. Science 2020, 368, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The microbiota of breast tissue and its association with breast cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef]
- Xuan, C.; Shamonki, J.M.; Chung, A.; DiNome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS ONE 2014, 9, e83744. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.A.; Bashir, M.; Rivas, M.N.; Duvall, K.; Sieling, P.A.; Pieber, T.R.; Vaishampayan, P.A.; Love, S.M.; Lee, D.J. Characterization of the microbiome of nipple aspirate fluid of breast cancer survivors. Sci. Rep. 2016, 6, 28061. [Google Scholar] [CrossRef]
- Wang, H.; Altemus, J.; Niazi, F.; Green, H.; Calhoun, B.C.; Sturgis, C.; Grobmyer, S.R.; Eng, C. Breast tissue, oral and urinary microbiomes in breast cancer. Oncotarget 2017, 8, 88122. [Google Scholar] [CrossRef]
- German, R.; Marino, N.; Hemmerich, C.; Podicheti, R.; Rusch, D.B.; Stiemsma, L.T.; Gao, H.; Xuei, X.; Rockey, P.; Storniolo, A.M. Exploring breast tissue microbial composition and the association with breast cancer risk factors. Breast Cancer Res. 2023, 25, 82. [Google Scholar] [CrossRef]
- Li, S.; Wang, T.; Ren, Y.; Liu, Z.; Gao, J.; Guo, Z. Prognostic impact of oral microbiome on survival of malignancies: A systematic review and meta-analysis. Syst. Rev. 2024, 13, 41. [Google Scholar] [CrossRef]
- Thompson, K.J.; Ingle, J.N.; Tang, X.; Chia, N.; Jeraldo, P.R.; Walther-Antonio, M.R.; Kandimalla, K.K.; Johnson, S.; Yao, J.Z.; Harrington, S.C. A comprehensive analysis of breast cancer microbiota and host gene expression. PLoS ONE 2017, 12, e0188873. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Chen, Y.-J.; Fan, T.-C.; Chang, N.-C.; Chen, Y.-J.; Midha, M.K.; Chen, T.-H.; Yang, H.-H.; Wang, Y.-T.; Yu, A.L. Analysis of microbial sequences in plasma cell-free DNA for early-onset breast cancer patients and healthy females. BMC Med. Genom. 2018, 11, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Byrd, D.A.; Wan, Y.; Ansong, D.; Clegg-Lamptey, J.N.; Wiafe-Addai, B.; Edusei, L.; Adjei, E.; Titiloye, N.; Dedey, F.; et al. The oral microbiome and breast cancer and nonmalignant breast disease, and its relationship with the fecal microbiome in the Ghana Breast Health Study. Int. J. Cancer 2022, 151, 1248–1260. [Google Scholar] [CrossRef]
- Feng, K.; Ren, F.; Wang, X. Relationships among breast, gut, and oral microbiota across diverse pathological types of breast cancer, a Chinese cohort study. Front. Mol. Biosci. 2023, 10, 1325552. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Ren, F.; Wang, X. Association between oral microbiome and seven types of cancers in East Asian population: A two-sample Mendelian randomization analysis. Front. Mol. Biosci. 2023, 10, 1327893. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.M.; Tran, H.T.T.; Long, J.; Shrubsole, M.J.; Cai, H.; Yang, Y.; Cai, Q.; Tran, T.V.; Zheng, W.; Shu, X.O. Gut microbiome in association with chemotherapy-induced toxicities among patients with breast cancer. Cancer 2024, 130, 2014–2030. [Google Scholar] [CrossRef]
- Ruiz-Saavedra, S.; Zapico, A.; Gonzalez, S.; Salazar, N.; de Los Reyes-Gavilan, C.G. Role of the intestinal microbiota and diet in the onset and progression of colorectal and breast cancers and the interconnection between both types of tumours. Microbiome Res. Rep. 2024, 3, 6. [Google Scholar] [CrossRef]
- Dutta, R.K.; Abu, Y.F.; Tao, J.; Chupikova, I.; Oleas, J.; Singh, P.K.; Vitari, N.A.; Qureshi, R.; Ramakrishnan, S.; Roy, S. Altered gut microbiome drives heightened pain sensitivity in a murine model of metastatic triple-negative breast cancer. Am. J. Cancer Res. 2024, 14, 274–299. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, S.; Yang, S.; Ma, F.; Gao, H. Recent progress in nanomaterials for bacteria-related tumor therapy. Biomater. Sci. 2024, 12, 1965–1980. [Google Scholar] [CrossRef] [PubMed]
- Mahno, N.E.; Tay, D.D.; Khalid, N.S.; Yassim, A.S.M.; Alias, N.S.; Termizi, S.A.; Kasian, J.; Mokhtar, N.M.; Ahmad, H.F. The Relationship Between Gut Microbiome Estrobolome and Breast Cancer: A Systematic Review of Current Evidences. Indian J. Microbiol. 2024, 64, 1–19. [Google Scholar] [CrossRef]
- Luu, T.H.; Michel, C.; Bard, J.-M.; Dravet, F.; Nazih, H.; Bobin-Dubigeon, C. Intestinal proportion of Blautia sp. is associated with clinical stage and histoprognostic grade in patients with early-stage breast cancer. Nutr. Cancer 2017, 69, 267–275. [Google Scholar] [CrossRef]
- Suzuki, R.; Rylander-Rudqvist, T.; Ye, W.; Saji, S.; Adlercreutz, H.; Wolk, A. Dietary fiber intake and risk of postmenopausal breast cancer defined by estrogen and progesterone receptor status--a prospective cohort study among Swedish women. Int. J. Cancer 2008, 122, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Shi, J.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Association of fecal microbial diversity and taxonomy with selected enzymatic functions. PLoS ONE 2012, 7, e39745. [Google Scholar] [CrossRef]
- Witt, B.L.; Tollefsbol, T.O. Molecular, Cellular, and Technical Aspects of Breast Cancer Cell Lines as a Foundational Tool in Cancer Research. Life 2023, 13, 2311. [Google Scholar] [CrossRef]
- Alvarez-Frutos, L.; Barriuso, D.; Duran, M.; Infante, M.; Kroemer, G.; Palacios-Ramirez, R.; Senovilla, L. Multiomics insights on the onset, progression, and metastatic evolution of breast cancer. Front. Oncol. 2023, 13, 1292046. [Google Scholar] [CrossRef] [PubMed]
- Zeber-Lubecka, N.; Kulecka, M.; Jagiello-Gruszfeld, A.; Dabrowska, M.; Kluska, A.; Piatkowska, M.; Baginska, K.; Glowienka, M.; Surynt, P.; Tenderenda, M.; et al. Breast cancer but not the menopausal status is associated with small changes of the gut microbiota. Front. Oncol. 2024, 14, 1279132. [Google Scholar] [CrossRef]
- He, K.; Meng, X.; Su, J.; Jiang, S.; Chu, M.; Huang, B. Oleanolic acid inhibits the tumor progression by regulating Lactobacillus through the cytokine-cytokine receptor interaction pathway in 4T1-induced mice breast cancer model. Heliyon 2024, 10, e27028. [Google Scholar] [CrossRef]
- Kumari, N.; Kumari, R.; Dua, A.; Singh, M.; Kumar, R.; Singh, P.; Duyar-Ayerdi, S.; Pradeep, S.; Ojesina, A.I.; Kumar, R. From Gut to Hormones: Unraveling the Role of Gut Microbiota in (Phyto)Estrogen Modulation in Health and Disease. Mol. Nutr. Food Res. 2024, 68, e2300688. [Google Scholar] [CrossRef]
- He, Z.; Xie, H.; Xu, H.; Wu, J.; Zeng, W.; He, Q.; Jobin, C.; Jin, S.; Lan, P. Chemotherapy-induced microbiota exacerbates the toxicity of chemotherapy through the suppression of interleukin-10 from macrophages. Gut Microbes 2024, 16, 2319511. [Google Scholar] [CrossRef]
- Heath, H.; Mogol, A.N.; Santaliz Casiano, A.; Zuo, Q.; Madak-Erdogan, Z. Targeting systemic and gut microbial metabolism in ER(+) breast cancer. Trends Endocrinol. Metab. 2024, 35, 321–330. [Google Scholar] [CrossRef]
- Franchi, M.; Piperigkou, Z.; Mastronikolis, N.S.; Karamanos, N. Extracellular matrix biomechanical roles and adaptation in health and disease. FEBS J. 2024, 291, 430–440. [Google Scholar] [CrossRef]
- McCarty, S.M.; Cochrane, C.A.; Clegg, P.D.; Percival, S.L. The role of endogenous and exogenous enzymes in chronic wounds: A focus on the implications of aberrant levels of both host and bacterial proteases in wound healing. Wound Repair Regen. 2012, 20, 125–136. [Google Scholar] [CrossRef]
- Wu, S.; Baum, M.M.; Kerwin, J.; Guerrero, D.; Webster, S.; Schaudinn, C.; VanderVelde, D.; Webster, P. Biofilm-specific extracellular matrix proteins of nontypeable Haemophilus influenzae. Pathog. Dis. 2014, 72, 143–160. [Google Scholar]
- Sironen, R.; Tammi, M.; Tammi, R.; Auvinen, P.; Anttila, M.; Kosma, V. Hyaluronan in human malignancies. Exp. Cell Res. 2011, 317, 383–391. [Google Scholar] [CrossRef]
- Kultti, A.; Li, X.; Jiang, P.; Thompson, C.B.; Frost, G.I.; Shepard, H.M. Therapeutic targeting of hyaluronan in the tumor stroma. Cancers 2012, 4, 873–903. [Google Scholar] [CrossRef]
- Erkan, M.; Hausmann, S.; Michalski, C.W.; Fingerle, A.A.; Dobritz, M.; Kleeff, J.; Friess, H. The role of stroma in pancreatic cancer: Diagnostic and therapeutic implications. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 454–467. [Google Scholar] [CrossRef]
- Jacobetz, M.A.; Chan, D.S.; Neesse, A.; Bapiro, T.E.; Cook, N.; Frese, K.K.; Feig, C.; Nakagawa, T.; Caldwell, M.E.; Zecchini, H.I. Hyaluronan impairs vascular function and drug delivery in a mouse model of pancreatic cancer. Gut 2013, 62, 112–120. [Google Scholar] [CrossRef]
- Turley, E.A.; Noble, P.W.; Bourguignon, L.Y. Signaling properties of hyaluronan receptors. J. Biol. Chem. 2002, 277, 4589–4592. [Google Scholar] [CrossRef]
- Chen, C.; Zhao, S.; Karnad, A.; Freeman, J.W. The biology and role of CD44 in cancer progression: Therapeutic implications. J. Hematol. Oncol. 2018, 11, 64. [Google Scholar] [CrossRef]
- Hart, M.E.; Hart, M.J.; Roop, A.J. Genotypic and phenotypic assessment of hyaluronidase among type strains of a select group of staphylococcal species. Int. J. Microbiol. 2009, 2009, 614371. [Google Scholar] [CrossRef]
- Kim, J.S.; Park, J.E.; Choi, S.H.; Kang, S.W.; Lee, J.H.; Lee, J.S.; Shin, M.; Park, S.H. ECM-targeting bacteria enhance chemotherapeutic drug efficacy by lowering IFP in tumor mouse models. J. Control. Release 2023, 355, 199–210. [Google Scholar] [CrossRef]
- Avagliano, A.; Fiume, G.; Ruocco, M.R.; Martucci, N.; Vecchio, E.; Insabato, L.; Russo, D.; Accurso, A.; Masone, S.; Montagnani, S. Influence of fibroblasts on mammary gland development, breast cancer microenvironment remodeling, and cancer cell dissemination. Cancers 2020, 12, 1697. [Google Scholar] [CrossRef]
- Feng, T.Y.; Azar, F.N.; Dreger, S.A.; Rosean, C.B.; McGinty, M.T.; Putelo, A.M.; Kolli, S.H.; Carey, M.A.; Greenfield, S.; Fowler, W.J.; et al. Reciprocal Interactions Between the Gut Microbiome and Mammary Tissue Mast Cells Promote Metastatic Dissemination of HR+ Breast Tumors. Cancer Immunol. Res. 2022, 10, 1309–1325. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Alam, K.; Roy, N.S.; Kaur, K.; Kaity, S.; Ravichandiran, V.; Roy, S. Exploring the interaction between extracellular matrix components in a 3D organoid disease model to replicate the pathophysiology of breast cancer. J. Exp. Clin. Cancer Res. 2023, 42, 343. [Google Scholar] [CrossRef]
- Provenzano, P.P.; Inman, D.R.; Eliceiri, K.W.; Keely, P.J. Matrix density-induced mechanoregulation of breast cell phenotype, signaling and gene expression through a FAK–ERK linkage. Oncogene 2009, 28, 4326–4343. [Google Scholar] [CrossRef]
- Pally, D.; Pramanik, D.; Bhat, R. An interplay between reaction-diffusion and cell-matrix adhesion regulates multiscale invasion in early breast carcinomatosis. Front. Physiol. 2019, 10, 458157. [Google Scholar] [CrossRef]
- Murphy, E.C.; Frick, I.-M. Gram-positive anaerobic cocci–commensals and opportunistic pathogens. FEMS Microbiol. Rev. 2013, 37, 520–553. [Google Scholar] [CrossRef]
- Bieri, U.; Scharl, M.; Sigg, S.; Szczerba, B.M.; Morsy, Y.; Ruschoff, J.H.; Schraml, P.H.; Krauthammer, M.; Hefermehl, L.J.; Eberli, D.; et al. Prospective observational study of the role of the microbiome in BCG responsiveness prediction (SILENT-EMPIRE): A study protocol. BMJ Open 2022, 12, e061421. [Google Scholar] [CrossRef]
- Wu, P.; Zhang, G.; Zhao, J.; Chen, J.; Chen, Y.; Huang, W.; Zhong, J.; Zeng, J. Profiling the urinary microbiota in male patients with bladder cancer in China. Front. Cell. Infect. Microbiol. 2018, 8, 167. [Google Scholar]
- Zeng, J.; Zhang, G.; Chen, C.; Li, K.; Wen, Y.; Zhao, J.; Wu, P. Alterations in urobiome in patients with bladder cancer and implications for clinical outcome: A single-institution study. Front. Cell. Infect. Microbiol. 2020, 10, 555508. [Google Scholar] [CrossRef]
- Demetriou, D.; Lockhat, Z.; Brzozowski, L.; Saini, K.S.; Dlamini, Z.; Hull, R. The Convergence of Radiology and Genomics: Advancing Breast Cancer Diagnosis with Radiogenomics. Cancers 2024, 16, 1076. [Google Scholar] [CrossRef]
- Ohmura, H.; Hanamura, F.; Okumura, Y.; Ando, Y.; Masuda, T.; Mimori, K.; Akashi, K.; Baba, E. Liquid biopsy for breast cancer and other solid tumors: A review of recent advances. Breast Cancer 2024. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, R.; Li, D.; Guo, R. Molecular mechanisms of circular RNA in breast cancer: A narrative review. Transl. Cancer Res. 2024, 13, 1139–1149. [Google Scholar] [CrossRef]
- Khalil, M.; Desouky, E.M.; Khaliefa, A.K.; Hozyen, W.G.; Mohamed, S.S.; Hasona, N.A. Insights into the Crosstalk Between miR-200a/lncRNA H-19 and IL-6/SIRT-1 Axis in Breast Cancer. J. Interferon Cytokine Res. 2024, 44, 191–197. [Google Scholar] [CrossRef]
- Napiorkowska-Mastalerz, M.; Wybranowski, T.; Bosek, M.; Kruszewski, S.; Rhone, P.; Ruszkowska-Ciastek, B. A Preliminary Evaluation of Advanced Oxidation Protein Products (AOPPs) as a Potential Approach to Evaluating Prognosis in Early-Stage Breast Cancer Patients and Its Implication in Tumour Angiogenesis: A 7-Year Single-Centre Study. Cancers 2024, 16, 1068. [Google Scholar] [CrossRef]
- Zare, H.; Bakherad, H.; Nasr Esfahani, A.; Norouzi, M.; Aghamollaei, H.; Mousavi Gargari, S.L.; Mahmoodi, F.; Aliomrani, M.; Ebrahimizadeh, W. Introduction of a new recombinant vaccine based on GRP78 for breast cancer immunotherapy and evaluation in a mouse model. Bioimpacts 2024, 14, 27829. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Filippou, C.; Themistocleous, S.C.; Marangos, G.; Panayiotou, Y.; Fyrilla, M.; Kousparou, C.A.; Pana, Z.D.; Tsioutis, C.; Johnson, E.O.; Yiallouris, A. Microbial Therapy and Breast Cancer Management: Exploring Mechanisms, Clinical Efficacy, and Integration within the One Health Approach. Int. J. Mol. Sci. 2024, 25, 1110. [Google Scholar] [CrossRef]
- Sharma, M.P.; Shukla, S.; Misra, G. Recent advances in breast cancer cell line research. Int. J. Cancer 2024, 154, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Avtanski, D.; Reddy, V.; Stojchevski, R.; Hadzi-Petrushev, N.; Mladenov, M. The Microbiome in the Obesity-Breast Cancer Axis: Diagnostic and Therapeutic Potential. Pathogens 2023, 12, 1402. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).