

Natural Compounds for Preventing Age-Related Diseases and Cancers


Abstract
1. Introduction
2. Circadian Rhythm and Implication for Chronic Diseases
3. Aging-Related Disease
3.1. Cardiovascular Diseases

{kind=link}
| Rank | Global | USA | Republic of Korea |
|---|---|---|---|
| 1 | Ischemia heart disease (115.7) | Ischemia heart disease (153.4) | Ischemia heart disease (54.8) |
| 2 | Stroke (80.7) | Alzheimer’s disease and other dementias (87.3) | Lower respiratory infection (52.0) |
| 3 | Chronic obstructive pulmonary disease (42.0) | Chronic obstructive pulmonary disease (59.4) | Stroke (50.0) |
| 4 | Lower respiratory infection (33.8) | Stroke (48.2) | Trachea, bronchus, and lung cancers (39.6) |
| 5 | Neonatal condition (26.5) | Trachea, bronchus, and lung cancers (47.3) | Self-harm (28.6) |
| 6 | Trachea, bronchus, and lung cancers (23.2) | Kidney diseases (26.1) | Alzheimer’s disease and other dementias (23.7) |
| 7 | Alzheimer’s disease and other dementias (21.4) | Drug use disorders (22.6) | Liver cancer (22.6) |
| 8 | Diarrheal diseases (19.8) | Hypertensive heart disease (20.0) | Colon and rectum cancers (19.9) |
| 9 | Diabetes mellitus (19.5) | Colon and rectum cancers (18.8) | Kidney diseases (19.7) |
| 10 | Kidney diseases (17.4) | Diabetes mellitus (18.7) | Stomach cancer (16.9) |
| Rank | Global | USA | Republic of Korea |
|---|---|---|---|
| 1 | Ischemia heart disease (455) | Ischemia heart disease (422) | Ischemia heart disease (136) |
| 2 | Stroke (320) | Alzheimer’s disease and other dementias (247) | Lower respiratory infection (133) |
| 3 | Chronic obstructive pulmonary disease (174) | Chronic obstructive pulmonary disease (167) | Stroke (124) |
| 4 | Trachea, bronchus, and lung cancers (93) | Stroke (132) | Trachea, bronchus, and lung cancers (100) |
| 5 | Lower respiratory infection (90) | Trachea, bronchus, and lung cancers (131) | Alzheimer’s disease and other dementias (61) |
| 6 | Alzheimer’s disease and other dementias (90) | Kidney diseases (71) | Liver cancer (55) |
| 7 | Diabetes mellitus (76) | Hypertensive heart disease (53) | Kidney diseases (50) |
| 8 | Kidney diseases (63) | Colon and rectum cancers (50) | Colon and rectum cancers (49) |
| 9 | Hypertensive heart disease (60) | Lower respiratory infection (49) | Chronic obstructive pulmonary disease (43) |
| 10 | Cirrhosis of the liver (53) | Diabetes mellitus (49) | Self-harm (42) |
3.1.1. Coronary Heart Disease (CHD)
3.1.2. Hypertension
3.1.3. Heart Failure
3.1.4. Stroke
3.2. Degenerative Brain Disorders
3.2.1. Alzheimer’s Disease
3.2.2. Parkinson Disease
3.3. Diabetes Mellitus
3.4. Musculoskeletal Disorders
3.4.1. Arthritis
3.4.2. Osteoporosis and Fracture
3.4.3. Muscle Wasting
3.5. Ophthalmology Disorders
3.6. Skin Aging
4. Cancer
| 1 ASR Cancer Incidence per 100,000 Population | ASR Cancer Mortality per 100,000 Population | |||
|---|---|---|---|---|
| Cancer Type/Site | Global | Republic of Korea | Global | Republic of Korea |
| Bladder | 5.58 | 4.1 | 1.82 | 1.1 |
| Brain, CNS | 3.47 | 3.0 | 2.59 | 1.5 |
| Breast | 46.82 | 33.1 | 12.65 | 2.9 |
| Cervix uteri | 14.12 | 3.7 | 7.08 | 0.7 |
| Colorectum | 18.35 | 26.4 | 8.05 | 6.6 |
| Corpus uteri | 8.37 | 4.2 | 1.72 | 0.4 |
| Esophagus | 4.97 | 2.4 | 4.26 | 1.0 |
| Gallbladder | 1.15 | 6.6 | 0.83 | 3.9 |
| Kidney | 4.42 | 7.4 | 1.46 | 0.9 |
| Leukemia | 5.26 | 5.7 | 3.093 | 1.9 |
| Lip and oral cavity | 4.00 | 4.3 | 1.92 | 1.1 |
| 2 Liver | 8.57 | 14.3 | 7.37 | 7.7 |
| 3 Lung | 23.62 | 27.6 | 16.76 | 13.4 |
| Multiple myeloma | 1.79 | 1.8 | 1.11 | 0.7 |
| Non-Hodgkin’s lymphoma | 5.57 | 6.7 | 2.38 | 1.8 |
| Ovary | 6.65 | 3.7 | 3.97 | 1.3 |
| Pancreas | 4.69 | 7.7 | 4.21 | 5.6 |
| Prostate | 29.42 | 18.4 | 7.27 | 1.4 |
| Stomach | 9.18 | 24.0 | 6.09 | 4.1 |
| Thyroid | 9.12 | 47.6 | 0.44 | 0.2 |
4.1. Breast Cancer
4.2. Prostate Cancer
4.3. Lung Cancer
4.4. Colorectal Cancer
4.5. Liver Cancer
4.6. Gastric Cancer
4.7. Pancreatic Cancer
5. Natural Compounds in Aging and Cancer Therapy and Prevention
5.1. Natural Compounds in Age-Related Diseases
5.1.1. Cardiovascular Diseases (CVDs)
5.1.2. Degenerative Brain Disorders
5.1.3. Diabetes Mellitus
5.1.4. Musculoskeletal Disorders
5.1.5. Ophthalmology Disorder
5.1.6. Skin Aging
5.2. Natural Compounds in Cancer Treatment
5.2.1. Breast Cancer
5.2.2. Prostate Cancer
5.2.3. Lung Cancer
5.2.4. Colorectal Cancer
5.2.5. Liver Cancer
5.2.6. Gastric Cancer
5.2.7. Pancreatic Cancer
6. Advantages and Limitations of Natural Compound Use and Future Prospects
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vitiello, R.; Bellieni, A.; Oliva, M.S.; Di Capua, B.; Fusco, D.; Careri, S.; Colloca, G.F.; Perisano, C.; Maccauro, G.; Lillo, M. The importance of geriatric and surgical co-management of elderly in muscoloskeletal oncology: A literature review. Orthop. Rev. 2020, 12, 8662. [Google Scholar] [CrossRef] [PubMed]
- Berben, L.; Floris, G.; Wildiers, H.; Hatse, S. Cancer and Aging: Two Tightly Interconnected Biological Processes. Cancers 2021, 13, 1400. [Google Scholar] [CrossRef] [PubMed]
- The importance of aging in cancer research. Nat. Aging 2022, 2, 365–366. [CrossRef] [PubMed]
- Chatsirisupachai, K.; Lagger, C.; de Magalhães, J.P. Age-associated differences in the cancer molecular landscape. Trends Cancer 2022, 8, 962–971. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Blasco, M.A. Cancer and ageing: Convergent and divergent mechanisms. Nat. Rev. Mol. Cell Biol. 2007, 8, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Aunan, J.R.; Cho, W.C.; Soreide, K. The Biology of Aging and Cancer: A Brief Overview of Shared and Divergent Molecular Hallmarks. Aging Dis. 2017, 8, 628–642. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Pietrocola, F.; Roiz-Valle, D.; Galluzzi, L.; Kroemer, G. Meta-hallmarks of aging and cancer. Cell Metab. 2023, 35, 12–35. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Pickett, H.A. Targeting telomeres: Advances in telomere maintenance mechanism-specific cancer therapies. Nat. Rev. Cancer 2022, 22, 515–532. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, J.; Shibata, H.; Kabata, M.; Kato, M.; Fukuda, K.; Tanaka, A.; Ohta, S.; Ukai, T.; Mitsunaga, K.; Yamada, Y.; et al. DMRT1-mediated reprogramming drives development of cancer resembling human germ cell tumors with features of totipotency. Nat. Commun. 2021, 12, 5041. [Google Scholar] [CrossRef]
- Cerella, C.; Grandjenette, C.; Dicato, M.; Diederich, M. Roles of Apoptosis and Cellular Senescence in Cancer and Aging. Curr. Drug Targets 2016, 17, 405–415. [Google Scholar] [CrossRef]
- Sitnikova, S.I.; Walker, J.A.; Prickett, L.B.; Morrow, M.; Valge-Archer, V.E.; Robinson, M.J.; Wilkinson, R.W.; Dovedi, S.J. Age-induced changes in anti-tumor immunity alter the tumor immune infiltrate and impact response to immuno-oncology treatments. Front. Immunol. 2023, 14, 1258291. [Google Scholar] [CrossRef]
- Drapela, S.; Ilter, D.; Gomes, A.P. Metabolic reprogramming: A bridge between aging and tumorigenesis. Mol. Oncol. 2022, 16, 3295–3318. [Google Scholar] [CrossRef]
- Aman, Y.; Schmauck-Medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N.; et al. Autophagy in healthy aging and disease. Nat. Aging 2021, 1, 634–650. [Google Scholar] [CrossRef] [PubMed]
- Fagiani, F.; Di Marino, D.; Romagnoli, A.; Travelli, C.; Voltan, D.; Di Cesare Mannelli, L.; Racchi, M.; Govoni, S.; Lanni, C. Molecular regulations of circadian rhythm and implications for physiology and diseases. Signal Transduct. Target. Ther. 2022, 7, 41. [Google Scholar] [CrossRef]
- Juste, Y.R.; Kaushik, S.; Bourdenx, M.; Aflakpui, R.; Bandyopadhyay, S.; Garcia, F.; Diaz, A.; Lindenau, K.; Tu, V.; Krause, G.J.; et al. Reciprocal regulation of chaperone-mediated autophagy and the circadian clock. Nat. Cell Biol. 2021, 23, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Li, X. Editorial: Aging and chronic disease: Public health challenge and education reform. Front. Public Health 2023, 11, 1175898. [Google Scholar] [CrossRef] [PubMed]
- Eliasen, B.; Teare, H.; Vrihenhoek, T. Shifting to Preventive Care in a New Health System. Available online: https://www.himss.org/resources/shifting-preventive-care-new-health-system (accessed on 18 April 2024).
- Baker, M.T.; Lu, P.; Parrella, J.A.; Leggette, H.R. Consumer Acceptance toward Functional Foods: A Scoping Review. Int. J. Environ. Res. Public Health 2022, 19, 1217. [Google Scholar] [CrossRef]
- Tsartsou, E.; Proutsos, N.; Papadopoulos, I.; Tzouvelekas, V.; Castanas, E.; Kampa, M. Consumers’ attitude toward dietary supplements and functional food: A prospective survey in a Greek population sample. Hormones 2021, 20, 177–188. [Google Scholar] [CrossRef]
- Tachie, C.; Nwachukwu, I.D.; Aryee, A.N.A. Trends and innovations in the formulation of plant-based foods. Food Prod. Process Nutr. 2023, 5, 16. [Google Scholar] [CrossRef]
- Najmi, A.; Javed, S.A.; Al Bratty, M.; Alhazmi, H.A. Modern Approaches in the Discovery and Development of Plant-Based Natural Products and Their Analogues as Potential Therapeutic Agents. Molecules 2022, 27, 349. [Google Scholar] [CrossRef]
- Salm, S.; Rutz, J.; van den Akker, M.; Blaheta, R.A.; Bachmeier, B.E. Current state of research on the clinical benefits of herbal medicines for non-life-threatening ailments. Front. Pharmacol. 2023, 14, 1234701. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.Y.; Lin, Q.; Hu, X.L.; Yang, Y.M.; Zheng, B.L.; Li, T.; Zhong, W.Q.; Wang, H.Y.; Zhang, Z.F.; Luo, B.J.; et al. Efficacy and safety of combined Chinese and Western medicine in the treatment of knee osteoarthritis: A prospective, multicenter cohort study. Front. Pharmacol. 2023, 14, 1176980. [Google Scholar] [CrossRef]
- Alarabei, A.A.; Abd Aziz, N.A.L.; Ab Razak, N.I.; Abas, R.; Bahari, H.; Abdullah, M.A.; Hussain, M.K.; Abdul Majid, A.M.S.; Basir, R. Immunomodulating Phytochemicals: An Insight Into Their Potential Use in Cytokine Storm Situations. Adv. Pharm. Bull. 2024, 14, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Adusei, S.; Otchere, J.K.; Oteng, P.; Mensah, R.Q.; Tei-Mensah, E. Phytochemical analysis, antioxidant and metal chelating capacity of Tetrapleura tetraptera. Heliyon 2019, 5, e02762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Gan, R.-Y.; Li, S.; Zhou, Y.; Li, A.-N.; Xu, D.-P.; Li, H.-B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Muscolo, A.; Mariateresa, O.; Giulio, T.; Mariateresa, R. Oxidative Stress: The Role of Antioxidant Phytochemicals in the Prevention and Treatment of Diseases. Int. J. Mol. Sci. 2024, 25, 3264. [Google Scholar] [CrossRef]
- Pyo, I.S.; Yun, S.; Yoon, Y.E.; Choi, J.-W.; Lee, S.-J. Mechanisms of Aging and the Preventive Effects of Resveratrol on Age-Related Diseases. Molecules 2020, 25, 4649. [Google Scholar] [CrossRef]
- Zhao, R.; Wang, L.; Liu, Y.; Shao, M.; Yang, W.; Fu, Y.; Gao, Q.; Feng, J.; Xing, Y.; Xiang, X. The influence of adjunctive traditional Chinese medicine therapy on survival in primary liver cancer: A real-world study based on electronic medical records. Front. Pharmacol. 2023, 14, 1231933. [Google Scholar] [CrossRef]
- Jang, J.-H.; Lee, T.-J. Mechanisms of Phytochemicals in Anti-Inflammatory and Anti-Cancer. Int. J. Mol. Sci. 2023, 24, 7863. [Google Scholar] [CrossRef]
- Sadiq, I.Z. Lifestyle medicine as a modality for prevention and management of chronic diseases. J. Taibah Univ. Med. Sci. 2023, 18, 1115–1117. [Google Scholar] [CrossRef]
- Akpınar, Ş.; Tek, N.A. Age-Related Changes in Circadian Rhythm and Association with Nutrition. Curr. Nutr. Rep. 2023, 12, 376–382. [Google Scholar] [CrossRef]
- Bae, S.A.; Fang, M.Z.; Rustgi, V.; Zarbl, H.; Androulakis, I.P. At the Interface of Lifestyle, Behavior, and Circadian Rhythms: Metabolic Implications. Front. Nutr. 2019, 6, 132. [Google Scholar] [CrossRef]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef]
- Juliana, N.; Azmi, L.; Effendy, N.M.; Mohd Fahmi Teng, N.I.; Abu, I.F.; Abu Bakar, N.N.; Azmani, S.; Yazit, N.A.A.; Kadiman, S.; Das, S. Effect of Circadian Rhythm Disturbance on the Human Musculoskeletal System and the Importance of Nutritional Strategies. Nutrients 2023, 15, 734. [Google Scholar] [CrossRef]
- Hastings, M.H.; Maywood, E.S.; Brancaccio, M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat. Rev. Neurosci. 2018, 19, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.S. Molecular components of the circadian clock in mammals. Diabetes Obes. Metab. 2015, 17 (Suppl. S1), 6–11. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y. Roles of circadian clocks in cancer pathogenesis and treatment. Exp. Mol. Med. 2021, 53, 1529–1538. [Google Scholar] [CrossRef]
- Solt, L.A.; Kojetin, D.J.; Burris, T.P. The REV-ERBs and RORs: Molecular links between circadian rhythms and lipid homeostasis. Future Med. Chem. 2011, 3, 623–638. [Google Scholar] [CrossRef] [PubMed]
- Guillaumond, F.; Dardente, H.; Giguere, V.; Cermakian, N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J. Biol. Rhythm. 2005, 20, 391–403. [Google Scholar] [CrossRef]
- Walker, W.H.; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar] [CrossRef]
- Kondratov, R.V.; Antoch, M.P. Circadian proteins in the regulation of cell cycle and genotoxic stress responses. Trends Cell Biol. 2007, 17, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Shafi, A.A.; McNair, C.M.; McCann, J.J.; Alshalalfa, M.; Shostak, A.; Severson, T.M.; Zhu, Y.; Bergman, A.; Gordon, N.; Mandigo, A.C.; et al. The circadian cryptochrome, CRY1, is a pro-tumorigenic factor that rhythmically modulates DNA repair. Nat. Commun. 2021, 12, 401. [Google Scholar] [CrossRef] [PubMed]
- Repouskou, A.; Prombona, A. c-MYC targets the central oscillator gene Per1 and is regulated by the circadian clock at the post-transcriptional level. Biochim. Biophys. Acta 2016, 1859, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, T.; Vila-Caballer, M.; Liu, J.; Schiffhauer, S.; Finkielstein, C.V. Association of the circadian factor Period 2 to p53 influences p53’s function in DNA-damage signaling. Mol. Biol. Cell 2015, 26, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.J.; Sellix, M.T.; Daniel, J.; Yamazaki, S.; Menaker, M.; Block, G.D. Chronic jet-lag increases mortality in aged mice. Curr. Biol. 2006, 16, R914–R916. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.J.; Purvis, T.E.; Hu, K.; Scheer, F.A. Circadian misalignment increases cardiovascular disease risk factors in humans. Proc. Natl. Acad. Sci. USA 2016, 113, E1402–E1411. [Google Scholar] [CrossRef] [PubMed]
- Kinouchi, K.; Sassone-Corsi, P. Metabolic rivalry: Circadian homeostasis and tumorigenesis. Nat. Rev. Cancer 2020, 20, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Rodriguez, V.A.; Rijo-Ferreira, F.; Green, C.B.; Takahashi, J.S. Importance of circadian timing for aging and longevity. Nat. Commun. 2021, 12, 2862. [Google Scholar] [CrossRef]
- Pariollaud, M.; Lamia, K.A. Cancer in the Fourth Dimension: What Is the Impact of Circadian Disruption? Cancer Discov. 2020, 10, 1455–1464. [Google Scholar] [CrossRef]
- Ward, E.M.; Germolec, D.; Kogevinas, M.; McCormick, D.; Vermeulen, R.; Anisimov, V.N.; Aronson, K.J.; Bhatti, P.; Cocco, P.; Costa, G.; et al. Carcinogenicity of night shift work. Lancet Oncol. 2019, 20, 1058–1059. [Google Scholar] [CrossRef]
- Seki, A.; Fishbein, M.C. Age-related cardiovascular changes and diseases. In Cardiovascular Pathology; Academic Press: Cambridge, MA, USA, 2022; pp. 85–121. [Google Scholar]
- Lakatta, E.G.; Levy, D. Arterial and Cardiac Aging: Major Shareholders in Cardiovascular Disease Enterprises. Circulation 2003, 107, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, N.S.; Amit, U.; Reibel, J.B.; Berlin, E.; Howell, K.; Ky, B. Cardiovascular disease and cancer: Shared risk factors and mechanisms. Nat. Rev. Cardiol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Hameau, R.; Gabrielli, L.; Garrido, M.; Guzman, A.M.; Retamal, I.; Vacarezza, M.J.; Greig, D.; Ocqueteau, M.; Sanchez, C.; Pizarro, M.; et al. Cardiotoxicity of anticancer therapies. Towards the implementation of cardio-oncology units. Rev. Med. Chile 2018, 146, 68–77. [Google Scholar] [CrossRef] [PubMed]
- WHO. The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 19 April 2024).
- WHO. Cause-Specific Mortality, 2000–2019. Available online: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates/ghe-leading-causes-of-death (accessed on 19 April 2024).
- Institute of Medicine (US). Committee on Social Security Cardiovascular Disability Criteria. In Cardiovascular Disability: Updating the Social Security Listings; National Academies Press (US): Washington, DC, USA, 2010. [Google Scholar] [CrossRef]
- Kannel, W.B. Coronary Heart Disease Risk Factors in the Elderly. Am. J. Geriatr. Cardiol. 2007, 11, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Poulter, N. Coronary heart disease is a multifactorial disease. Am. J. Hypertens. 1999, 12, 92–95. [Google Scholar] [CrossRef]
- Salmon, T.; Hatherley, J.; Sankaranarayanan, R. Hypertension: A cardiovascular risk factor. In Hypertension (Oxford Cardiology Library) 3E; Nadar, S.K., Lip, G.Y.H., Nadar, S., Lip, G., Eds.; Oxford University Press: Oxford, UK, 2022; pp. 41–48. [Google Scholar]
- Fisher, N.D.L.; Curfman, G. Hypertension—A Public Health Challenge of Global Proportions. JAMA 2018, 320, 1757–1759. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, M.H.; Liu, P.; Roth, G.A.; Ng, M.; Biryukov, S.; Marczak, L.; Alexander, L.; Estep, K.; Hassen Abate, K.; Akinyemiju, T.F.; et al. Global Burden of Hypertension and Systolic Blood Pressure of at Least 110 to 115 mm Hg, 1990–2015. JAMA 2017, 317, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.T.; Stefanescu, A.; He, J. The global epidemiology of hypertension. Nat. Rev. Nephrol. 2020, 16, 223–237. [Google Scholar] [CrossRef]
- Poznyak, A.V.; Sadykhov, N.K.; Kartuesov, A.G.; Borisov, E.E.; Melnichenko, A.A.; Grechko, A.V.; Orekhov, A.N. Hypertension as a risk factor for atherosclerosis: Cardiovascular risk assessment. Front. Cardiovasc. Med. 2022, 9, 959285. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Han, X.; Sun, J.; Li, C.; Adhikari, B.K.; Zhang, J.; Miao, X.; Chen, Z. Cardio-Oncology: A Myriad of Relationships Between Cardiovascular Disease and Cancer. Front. Cardiovasc. Med. 2022, 9, 727487. [Google Scholar] [CrossRef]
- Grossman, E.; Messerli, F.H.; Boyko, V.; Goldbourt, U. Is there an association between hypertension and cancer mortality? Am. J. Med. 2002, 112, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Schwinger, R.H.G. Pathophysiology of heart failure. Cardiovasc. Diagn. Ther. 2021, 11, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Katsagoni, C.N.; Kokkinos, P.; Sidossis, L.S. Heart Failure. In Prevention and Management of Cardiovascular and Metabolic Disease; John Wiley & Sons: Hoboken, NJ, USA, 2023; pp. 169–189. [Google Scholar] [CrossRef]
- Meijers, W.C.; Maglione, M.; Bakker, S.J.L.; Oberhuber, R.; Kieneker, L.M.; de Jong, S.; Haubner, B.J.; Nagengast, W.B.; Lyon, A.R.; van der Vegt, B.; et al. Heart Failure Stimulates Tumor Growth by Circulating Factors. Circulation 2018, 138, 678–691. [Google Scholar] [CrossRef] [PubMed]
- OHSU. Understanding Stroke. Available online: https://www.ohsu.edu/brain-institute/understanding-stroke (accessed on 12 April 2024).
- Dewangan, K.K.; Junaid, M.; Dixit, R.; Kamble, N.; Patil, A.R. Association of modifiable risk factors among stroke patients attending a tertiary care hospital at Durg district of Chhattisgarh: A case-control study. Int. J. Community Med. Public Health 2023, 10, 2180–2184. [Google Scholar] [CrossRef]
- Varkey, B.P.; Joseph, J.; Varghese, A.; Sharma, S.K.; Mathews, E.; Dhandapani, M.; Narasimha, V.L.; Kuttan, R.; Shah, S.; Dabla, S.; et al. The Distribution of Lifestyle Risk Factors Among Patients with Stroke in the Indian Setting: Systematic Review and Meta-Analysis. Ann. Neurosci. 2022, 30, 40–53. [Google Scholar] [CrossRef]
- Boehme, A.K.; Esenwa, C.; Elkind, M.S. Stroke Risk Factors, Genetics, and Prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, A.M.; Stowie, A.; Hill, A.; Ellis, I.; Rhodes, M.K.; Pollock, D.M.; Davidson, A.J. Long-term circadian disruption shortens life span and dampens blood pressure diurnal rhythms in stroke-prone spontaneously hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2023, 325, H106–H112. [Google Scholar] [CrossRef]
- Penaloza-Martinez, E.; Moreno, G.; Aroca-Crevillen, A.; Huertas, S.; Vicent, L.; Rosillo, N.; Hidalgo, A.; Bueno, H. Circadian rhythms in thrombosis and atherothrombotic events. Front. Biosci. 2022, 27, 51. [Google Scholar] [CrossRef] [PubMed]
- Mehu, M.; Narasimhulu, C.A.; Singla, D.K. Inflammatory Cells in Atherosclerosis. Antioxidants 2022, 11, 233. [Google Scholar] [CrossRef]
- Wang, C.; Yang, X.; Guo, Z.; Zhu, G.; Fan, L. Circadian gene CLOCK accelerates atherosclerosis by promoting endothelial autophagy. Biotechnol. Genet. Eng. Rev. 2023, 1–16. [Google Scholar] [CrossRef]
- Lu, H.; Wang, Y.; Fan, H.; Wang, Y.; Fan, S.; Hu, S.; Shen, H.; Li, H.; Xue, Q.; Ni, J.; et al. GluA1 Degradation by Autophagy Contributes to Circadian Rhythm Effects on Cerebral Ischemia Injury. J. Neurosci. 2023, 43, 2381–2397. [Google Scholar] [CrossRef]
- Rabinovich-Nikitin, I.; Lieberman, B.; Martino, T.A.; Kirshenbaum, L.A. Circadian-Regulated Cell Death in Cardiovascular Diseases. Circulation 2019, 139, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Wyse, C.; O’Malley, G.; Coogan, A.N.; McConkey, S.; Smith, D.J. Seasonal and daytime variation in multiple immune parameters in humans: Evidence from 329,261 participants of the UK Biobank cohort. iScience 2021, 24, 102255. [Google Scholar] [CrossRef] [PubMed]
- Dierickx, P.; Zhu, K.; Carpenter, B.J.; Jiang, C.; Vermunt, M.W.; Xiao, Y.; Luongo, T.S.; Yamamoto, T.; Martí-Pàmies, Í.; Mia, S.; et al. Circadian REV-ERBs repress E4bp4 to activate NAMPT-dependent NAD+ biosynthesis and sustain cardiac function. Nat. Cardiovasc. Res. 2021, 1, 45–58. [Google Scholar] [CrossRef]
- Park, B.H.; Lee, B.K.; Ahn, J.; Kim, N.S.; Park, J.; Kim, Y. Association of Participation in Health Check-ups with Risk Factors for Cardiovascular Diseases. J. Korean Med. Sci. 2021, 36, e19. [Google Scholar] [CrossRef] [PubMed]
- Rippe, J.M. Lifestyle Medicine: The Health Promoting Power of Daily Habits and Practices. Am. J. Lifestyle Med. 2018, 12, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Blinkouskaya, Y.; Caçoilo, A.; Gollamudi, T.; Jalalian, S.; Weickenmeier, J. Brain aging mechanisms with mechanical manifestations. Mech. Ageing Dev. 2021, 200, 111575. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, L.; Wåhlin, A. The many facets of brain aging. eLife 2020, 9, e56640. [Google Scholar] [CrossRef] [PubMed]
- John, O.O.; Amarachi, I.S.; Chinazom, A.P.; Adaeze, E.; Kale, M.B.; Umare, M.D.; Upaganlawar, A.B. Phytotherapy: A promising approach for the treatment of Alzheimer’s disease. Pharmacol. Res.-Mod. Chin. Med. 2022, 2, 100030. [Google Scholar] [CrossRef]
- Nixon, R.A. The role of autophagy in neurodegenerative disease. Nat. Med. 2013, 19, 983–997. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, W.; Zhao, M.; Ma, L.; Jiang, X.; Pei, H.; Cao, Y.; Li, H. Interaction between Aβ and Tau in the Pathogenesis of Alzheimer’s Disease. Int. J. Biol. Sci. 2021, 17, 2181–2192. [Google Scholar] [CrossRef]
- Yap, L.E.; Hunt, J.E.; Turner, R.S. Aging as a target for the prevention and treatment of Alzheimer’s disease. Front. Neurol. 2024, 15, 1376104. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Shin, S.J.; Nam, Y.; Park, Y.H.; Kim, B.-H.; Park, H.H.; Kumar, V.; Yoo, D.-H.; Lee, Y.Y.; Hoe, H.-S.; et al. Korean red ginseng polysaccharide as a potential therapeutic agent targeting tau pathology in Alzheimer’s disease. Int. J. Biol. Macromol. 2024, 263, 130516. [Google Scholar] [CrossRef] [PubMed]
- Marvi, F.; Chen, Y.H.; Sawan, M. Alzheimer’s Disease Diagnosis in the Preclinical Stage: Normal Aging or Dementia. IEEE Rev. Biomed. Eng. 2024. [Google Scholar] [CrossRef] [PubMed]
- Macedo, A.C.; Therriault, J.; Tissot, C.; Fernandez-Arias, J.; Ferreira, P.C.L.; Vitali, P.; Servaes, S.; Rahmouni, N.; Vermeiren, M.; Bezgin, G.; et al. Predicting functional decline in aging and Alzheimer’s disease with PET-based Braak staging. Brain Commun. 2024, 6, fcae043. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Bi, F.; Wang, C.; Huang, Y.; Xu, Y.; Liang, S.; Cai, T.; Xu, X.; Dong, L.; Li, T.; et al. The circadian rhythm: A new target of natural products that can protect against diseases of the metabolic system, cardiovascular system, and nervous system. J. Adv. Res. 2024, in press. [Google Scholar] [CrossRef] [PubMed]
- Opara, J.; Małecki, A.; Małecka, E.; Socha, T. Motor assessment in Parkinson`s disease. Ann. Agric. Environ. Med. 2017, 24, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Thein, J.; Linnhoff, S.; Voges, J.; Galazky, I.; Zaehle, T. Enhancing Attentional Performance in Parkinson’s Disease: The Impact of Combined Deep Brain Stimulation of the Substantia Nigra Pars Reticulata and the Subthalamic Nucleus. Am. J. Med. 2024, 137, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Tekriwal, A.; Felsen, G.; Ojemann, S.G.; Abosch, A.; Thompson, J.A. Motor context modulates substantia nigra pars reticulata spike activity in patients with Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2022, 93, 386–394. [Google Scholar] [CrossRef]
- Gregory, J.; Vengalasetti, Y.V.; Bredesen, D.E.; Rao, R.V. Neuroprotective Herbs for the Management of Alzheimer’s Disease. Biomolecules 2021, 11, 543. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, Z.; Cui, W. Marine-Derived Natural Compounds for the Treatment of Parkinson’s Disease. Mar. Drugs 2019, 17, 221. [Google Scholar] [CrossRef] [PubMed]
- Drobny, A.; Boros, F.A.; Balta, D.; Prieto Huarcaya, S.; Caylioglu, D.; Qazi, N.; Vandrey, J.; Schneider, Y.; Dobert, J.P.; Pitcairn, C.; et al. Reciprocal effects of alpha-synuclein aggregation and lysosomal homeostasis in synucleinopathy models. Transl. Neurodegener. 2023, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Lv, Q.-K.; Xie, W.-Y.; Gong, S.-Y.; Zhuang, S.; Liu, J.-Y.; Mao, C.-J.; Liu, C.-F. Circadian disruption and sleep disorders in neurodegeneration. Transl. Neurodegener. 2023, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Fedosov, I.; Penzel, T.; Li, D.; Yu, T.; Telnova, V.; Kaybeleva, E.; Saranceva, E.; Terskov, A.; Khorovodov, A.; et al. Brain Waste Removal System and Sleep: Photobiomodulation as an Innovative Strategy for Night Therapy of Brain Diseases. Int. J. Mol. Sci. 2023, 24, 3221. [Google Scholar] [CrossRef] [PubMed]
- Budkowska, M.; Cecerska-Heryć, E.; Marcinowska, Z.; Siennicka, A.; Dołęgowska, B. The Influence of Circadian Rhythm on the Activity of Oxidative Stress Enzymes. Int. J. Mol. Sci. 2022, 23, 14275. [Google Scholar] [CrossRef] [PubMed]
- Vallee, A.; Lecarpentier, Y.; Guillevin, R.; Vallee, J.N. Circadian rhythms, Neuroinflammation and Oxidative Stress in the Story of Parkinson’s Disease. Cells 2020, 9, 314. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Akioma, M.; Yuan, Z. Relationship between circadian rhythm and brain cognitive functions. Front. Optoelectron. 2021, 14, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Logan, R.W.; McClung, C.A. Rhythms of life: Circadian disruption and brain disorders across the lifespan. Nat. Rev. Neurosci. 2019, 20, 49–65. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.M.M.; Chua, Z.J.Y.; Tan, J.C.; Yang, Y.; Liao, Z.; Zhao, Y. From Pre-Diabetes to Diabetes: Diagnosis, Treatments and Translational Research. Medicina 2019, 55, 546. [Google Scholar] [CrossRef]
- Egan, A.M.; Dinneen, S.F. What is diabetes? Medicine 2019, 47, 1–4. [Google Scholar] [CrossRef]
- Banday, M.Z.; Sameer, A.S.; Nissar, S. Pathophysiology of diabetes: An overview. Avicenna J. Med. 2020, 10, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Hussain, M.E. Obesity and diabetes: An update. Diabetes Metab. Syndr. 2017, 11, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Rowley, W.R.; Bezold, C.; Arikan, Y.; Byrne, E.; Krohe, S. Diabetes 2030: Insights from Yesterday, Today, and Future Trends. Popul. Health Manag. 2017, 20, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- Kalyani, R.R.; Golden, S.H.; Cefalu, W.T. Diabetes and Aging: Unique Considerations and Goals of Care. Diabetes Care 2017, 40, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Halim, M.; Halim, A. The effects of inflammation, aging and oxidative stress on the pathogenesis of diabetes mellitus (type 2 diabetes). Diabetes Metab. Syndr. 2019, 13, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E. The relative contributions of insulin resistance and beta-cell dysfunction to the pathophysiology of Type 2 diabetes. Diabetologia 2003, 46, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, E.J.; LeRoith, D. The proliferating role of insulin and insulin-like growth factors in cancer. Trends Endocrinol. Metab. 2010, 21, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Jang, J.-Y.; Kim, D.-L.; Rhyu, Y.A.; Lee, S.E.; Ko, S.-H.; Han, K.; Song, K.-H. Site-specific cancer risk in patients with type 2 diabetes: A nationwide population-based cohort study in Korea. Korean J. Intern. Med. 2020, 35, 641. [Google Scholar] [CrossRef]
- Spampinato, S.F.; Caruso, G.I.; De Pasquale, R.; Sortino, M.A.; Merlo, S. The treatment of impaired wound healing in diabetes: Looking among old drugs. Pharmaceuticals 2020, 13, 60. [Google Scholar] [CrossRef]
- Wang, S.Y.; Liu, J.F.; Huang, Y.P.; Chang, Y.Y. The Diabetic Foot Amputation Decision-Making Process. Adv. Skin. Wound Care 2018, 31, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, S.; Ahmad, I.; Arif, H.; Ammara, U.E.; Majeed, A. Artificial pancreas control strategies used for type 1 diabetes control and treatment: A comprehensive analysis. Appl. Syst. Innov. 2020, 3, 31. [Google Scholar] [CrossRef]
- Quoc Lam, B.; Shrivastava, S.K.; Shrivastava, A.; Shankar, S.; Srivastava, R.K. The Impact of obesity and diabetes mellitus on pancreatic cancer: Molecular mechanisms and clinical perspectives. J. Cell. Mol. Med. 2020, 24, 7706–7716. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Scragg, R.; Petrov, M.S. Postpancreatitis diabetes confers higher risk for pancreatic cancer than type 2 diabetes: Results from a nationwide cancer registry. Diabetes Care 2020, 43, 2106–2112. [Google Scholar] [CrossRef]
- ElSayed, N.A.; Aleppo, G.; Aroda, V.R.; Bannuru, R.R.; Brown, F.M.; Bruemmer, D.; Collins, B.S.; Hilliard, M.E.; Isaacs, D.; Johnson, E.L.; et al. 8. Obesity and Weight Management for the Prevention and Treatment of Type 2 Diabetes: Standards of Care in Diabetes-2023. Diabetes Care 2023, 46, S128–S139. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.; Bain, S.; Kanamarlapudi, V. A Review of Current Trends with Type 2 Diabetes Epidemiology, Aetiology, Pathogenesis, Treatments and Future Perspectives. Diabetes Metab. Syndr. Obes. 2021, 14, 3567–3602. [Google Scholar] [CrossRef]
- Verma, P.; Dubey, R. Circadian Rhythm Disruption: Special Reference to Type 2 Diabetes Mellitus. In Pharmacological and Molecular Perspectives on Diabetes; Bentham Science: Sharjah, United Arab Emirates, 2022; pp. 134–145. [Google Scholar]
- Gabriel, B.M.; Altintas, A.; Smith, J.A.B.; Sardon-Puig, L.; Zhang, X.; Basse, A.L.; Laker, R.C.; Gao, H.; Liu, Z.; Dollet, L.; et al. Disrupted circadian oscillations in type 2 diabetes are linked to altered rhythmic mitochondrial metabolism in skeletal muscle. Sci. Adv. 2021, 7, eabi9654. [Google Scholar] [CrossRef] [PubMed]
- Zitting, K.M.; Vetrivelan, R.; Yuan, R.K.; Vujovic, N.; Wang, W.; Bandaru, S.S.; Quan, S.F.; Klerman, E.B.; Scheer, F.; Buxton, O.M.; et al. Chronic circadian disruption on a high-fat diet impairs glucose tolerance. Metabolism 2022, 130, 155158. [Google Scholar] [CrossRef]
- Hariri, A.; Mirian, M.; Zarrabi, A.; Kohandel, M.; Amini-Pozveh, M.; Aref, A.R.; Tabatabaee, A.; Prabhakar, P.K.; Sivakumar, P.M. The circadian rhythm: An influential soundtrack in the diabetes story. Front. Endocrinol. 2023, 14, 1156757. [Google Scholar] [CrossRef]
- Li, B.; Chen, D. Degenerative musculoskeletal diseases: Pathology and treatments. J. Orthop. Transl. 2019, 17, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Karanfil, Y.; Eşme, M.; Korkusuz, F. Musculoskeletal physiological changes in aging. In Beauty, Aging, and AntiAging; Academic Press: Cambridge, MA, USA, 2023; pp. 83–108. [Google Scholar]
- Cento, A.S.; Leigheb, M.; Caretti, G.; Penna, F. Exercise and Exercise Mimetics for the Treatment of Musculoskeletal Disorders. Curr. Osteoporos. Rep. 2022, 20, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Bornhoft, L.; Arvidsson, D.; Bergenheim, A.; Borjesson, M.; Fridolfsson, J.; Hellgren, M.; Nordeman, L.; Larsson, M.E. Development and feasibility of a function-based preventive intervention for lifestyle-related disorders. BMC Public Health 2024, 24, 681. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wei, J.; Xiao, W.; Xie, W.; Ru, Q.; Chen, L.; Wu, Y.; Mobasheri, A.; Li, Y. Insights into the Notch signaling pathway in degenerative musculoskeletal disorders: Mechanisms and perspectives. Biomed. Pharmacother. 2023, 169, 115884. [Google Scholar] [CrossRef] [PubMed]
- Jordan, K.P.; Croft, P. Mortality and cancer in patients with new musculoskeletal episodes: A cohort study. Br. J. Gen. Pract. 2010, 60, e105–e111. [Google Scholar] [CrossRef]
- WHO. Osteoarthritis. Available online: https://www.who.int/news-room/fact-sheets/detail/osteoarthritis (accessed on 21 April 2024).
- Kong, H.; Han, J.-J.; Dmitrii, G.; Zhang, X.-a. Phytochemicals against Osteoarthritis by Inhibiting Apoptosis. Molecules 2024, 29, 1487. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Wang, X.-Q.; Zhang, X.-A. Exercise for Osteoarthritis: A Literature Review of Pathology and Mechanism. Front. Aging Neurosci. 2022, 14, 854026. [Google Scholar] [CrossRef] [PubMed]
- Prasadam, I.; Crawford, R.; Fan, X.; Sun, A.; Sonar, Y.; Wu, X.; Wakale, S. How are Aging and Osteoarthritis Related? Aging Dis. 2023, 14, 592–604. [Google Scholar] [CrossRef]
- Mohammed, A.; Alshamarri, T.; Adeyeye, T.; Lazariu, V.; McNutt, L.A.; Carpenter, D.O. A comparison of risk factors for osteo- and rheumatoid arthritis using NHANES data. Prev. Med. Rep. 2020, 20, 101242. [Google Scholar] [CrossRef]
- Cho, M.H.; Cho, J.H.; Eun, Y.; Han, K.; Jung, J.; Cho, I.Y.; Yoo, J.E.; Lee, H.; Kim, H.; Park, S.Y.; et al. Rheumatoid Arthritis and Risk of Lung Cancer: A Nationwide Cohort Study. J. Thorac. Oncol. 2024, 19, 216–226. [Google Scholar] [CrossRef]
- McBeth, J.; Symmons, D.P.; Silman, A.J.; Allison, T.; Webb, R.; Brammah, T.; Macfarlane, G.J. Musculoskeletal pain is associated with a long-term increased risk of cancer and cardiovascular-related mortality. Rheumatology 2009, 48, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Pferdehirt, L.; Damato, A.R.; Dudek, M.; Meng, Q.J.; Herzog, E.D.; Guilak, F. Synthetic gene circuits for preventing disruption of the circadian clock due to interleukin-1-induced inflammation. Sci. Adv. 2022, 8, eabj8892. [Google Scholar] [CrossRef]
- Song, X.; Ma, T.; Hu, H.; Zhao, M.; Bai, H.; Wang, X.; Liu, L.; Li, T.; Sheng, X.; Xu, X.; et al. Chronic Circadian Rhythm Disturbance Accelerates Knee Cartilage Degeneration in Rats Accompanied by the Activation of the Canonical Wnt/beta-Catenin Signaling Pathway. Front. Pharmacol. 2021, 12, 760988. [Google Scholar] [CrossRef]
- Hand, L.E.; Dickson, S.H.; Freemont, A.J.; Ray, D.W.; Gibbs, J.E. The circadian regulator Bmal1 in joint mesenchymal cells regulates both joint development and inflammatory arthritis. Arthritis Res. Ther. 2019, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Kanis, J.A.; McCloskey, E.V.; Johansson, H.; Oden, A.; Melton, L.J.; Khaltaev, N. A reference standard for the description of osteoporosis. Bone 2008, 42, 467–475. [Google Scholar] [CrossRef]
- Ibrahim, N.A.; Nabil, N.; Ghaleb, S. Pathophysiology of the Risk Factors Associated with Osteoporosis and their Correlation to the T-score Value in Patients with Osteopenia and Osteoporosis in the United Arab Emirates. J. Pharm. Bioallied Sci. 2019, 11, 364–372. [Google Scholar] [CrossRef]
- Al-Toma, A.; Herman, A.; Lems, W.F.; Mulder, C.J.J. The Dietary and Non-Dietary Management of Osteoporosis in Adult-Onset Celiac Disease: Current Status and Practical Guidance. Nutrients 2022, 14, 4554. [Google Scholar] [CrossRef] [PubMed]
- Ilias, I.; Milionis, C.; Zoumakis, E. An Overview of Glucocorticoid-Induced Osteoporosis. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; South Dartmouth, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK278968/ (accessed on 17 April 2024).
- Emmanuelle, N.E.; Marie-Cecile, V.; Florence, T.; Jean-Francois, A.; Francoise, L.; Coralie, F.; Alexia, V. Critical Role of Estrogens on Bone Homeostasis in Both Male and Female: From Physiology to Medical Implications. Int. J. Mol. Sci. 2021, 22, 1568. [Google Scholar] [CrossRef]
- Rani, J.; Swati, S.; Meeta, M.; Singh, S.H.; Tanvir, T.; Madan, A. Postmenopausal Osteoporosis: Menopause Hormone Therapy and Selective Estrogen Receptor Modulators. Indian. J. Orthop. 2023, 57, 105–114. [Google Scholar] [CrossRef]
- Genazzani, A.R.; Monteleone, P.; Giannini, A.; Simoncini, T. Hormone therapy in the postmenopausal years: Considering benefits and risks in clinical practice. Hum. Reprod. Update 2021, 27, 1115–1150. [Google Scholar] [CrossRef]
- Esen, E.; Lee, S.-Y.; Wice, B.M.; Long, F. PTH Promotes Bone Anabolism by Stimulating Aerobic Glycolysis via IGF Signaling. J. Bone Miner. Res. 2015, 30, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Appelman-Dijkstra, N.M.; Papapoulos, S.E. Clinical advantages and disadvantages of anabolic bone therapies targeting the WNT pathway. Nat. Rev. Endocrinol. 2018, 14, 605–623. [Google Scholar] [CrossRef]
- Clarke, J. Ageing stem cells hold the key to age-related bone degeneration. Nat. Rev. Rheumatol. 2021, 17, 580. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, T.H.; Marecic, O.; McArdle, A.; Sinha, R.; Gulati, G.S.; Tong, X.; Wang, Y.; Steininger, H.M.; Hoover, M.Y.; Koepke, L.S.; et al. Aged skeletal stem cells generate an inflammatory degenerative niche. Nature 2021, 597, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Ming, J. The role of circadian rhythm in osteoporosis; a review. Front. Cell Dev. Biol. 2022, 10, 960456. [Google Scholar] [CrossRef] [PubMed]
- Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef] [PubMed]
- Abe, Y.O.; Yoshitane, H.; Kim, D.W.; Kawakami, S.; Koebis, M.; Nakao, K.; Aiba, A.; Kim, J.K.; Fukada, Y. Rhythmic transcription of Bmal1 stabilizes the circadian timekeeping system in mammals. Nat. Commun. 2022, 13, 4652. [Google Scholar] [CrossRef] [PubMed]
- Jinteng, L.; Peitao, X.; Wenhui, Y.; Guiwen, Y.; Feng, Y.; Xiaojun, X.; Zepeng, S.; Jiajie, L.; Yunshu, C.; Zhaoqiang, Z.; et al. BMAL1-TTK-H2Bub1 loop deficiency contributes to impaired BM-MSC-mediated bone formation in senile osteoporosis. Mol. Ther. Nucleic Acids 2023, 31, 568–585. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.F.C.; Ortega, D.Y.G. Perioperative Care of the Surgical Patient: Bone and Soft Tissue Tumors. In Perioperative Care of the Cancer Patient; Elsevier: Amsterdam, The Netherlands, 2023; pp. 351–355. [Google Scholar]
- Macedo, F.; Ladeira, K.; Pinho, F.; Saraiva, N.; Bonito, N.; Pinto, L.; Goncalves, F. Bone Metastases: An Overview. Oncol. Rev. 2017, 11, 321. [Google Scholar] [CrossRef]
- Chen, K.; Gao, P.; Li, Z.; Dai, A.; Yang, M.; Chen, S.; Su, J.; Deng, Z.; Li, L. Forkhead Box O Signaling Pathway in Skeletal Muscle Atrophy. Am. J. Pathol. 2022, 192, 1648–1657. [Google Scholar] [CrossRef]
- Bodine, S.C.; Sinha, I.; Sweeney, H.L. Mechanisms of Skeletal Muscle Atrophy and Molecular Circuitry of Stem Cell Fate in Skeletal Muscle Regeneration and Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2023, 78, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Deng, J.; Qiu, Y.; Gao, J.; Li, J.; Guan, L.; Lee, H.; Zhou, Q.; Xiao, J. Non-coding RNA basis of muscle atrophy. Mol. Ther. Nucleic Acids 2021, 26, 1066–1078. [Google Scholar] [CrossRef]
- Li, C.W.; Yu, K.; Shyh-Chang, N.; Jiang, Z.; Liu, T.; Ma, S.; Luo, L.; Guang, L.; Liang, K.; Ma, W.; et al. Pathogenesis of sarcopenia and the relationship with fat mass: Descriptive review. J. Cachexia Sarcopenia Muscle 2022, 13, 781–794. [Google Scholar] [CrossRef] [PubMed]
- Prado, C.M.; Batsis, J.A.; Donini, L.M.; Gonzalez, M.C.; Siervo, M. Sarcopenic obesity in older adults: A clinical overview. Nat. Rev. Endocrinol. 2024, 20, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Chun, S.; Shin, D.W.; Han, K.; Jung, J.H.; Kim, B.; Jung, H.-W.; Son, K.Y.; Lee, S.-P.; Lee, S.C. The Timed Up and Go test and the ageing heart: Findings from a national health screening of 1,084,875 community-dwelling older adults. Eur. J. Prev. Cardiol. 2021, 28, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Zhang, L. Cancer Cachexia: Definition, Staging, and Emerging Treatments. Cancer Manag. Res. 2020, 12, 5597–5605. [Google Scholar] [CrossRef]
- Malhan, D.; Yalcin, M.; Schoenrock, B.; Blottner, D.; Relogio, A. Skeletal muscle gene expression dysregulation in long-term spaceflights and aging is clock-dependent. NPJ Microgravity 2023, 9, 30. [Google Scholar] [CrossRef]
- Luo, X.; Yang, X.; Yang, Y.; Li, H.; Cui, H.; Cao, X. The interrelationship between inflammatory cytokines and skeletal muscle decay from the viewpoint of circadian rhythms. Arch. Physiol. Biochem. 2022, 128, 1559–1565. [Google Scholar] [CrossRef]
- Choi, Y.; Cho, J.; No, M.H.; Heo, J.W.; Cho, E.J.; Chang, E.; Park, D.H.; Kang, J.H.; Kwak, H.B. Re-Setting the Circadian Clock Using Exercise against Sarcopenia. Int. J. Mol. Sci. 2020, 21, 3106. [Google Scholar] [CrossRef]
- Gao, H.; Xiong, X.; Lin, Y.; Chatterjee, S.; Ma, K. The clock regulator Bmal1 protects against muscular dystrophy. Exp. Cell Res. 2020, 397, 112348. [Google Scholar] [CrossRef]
- Coleman-Belin, J.; Harris, A.; Chen, B.; Zhou, J.; Ciulla, T.; Verticchio, A.; Antman, G.; Chang, M.; Siesky, B. Aging Effects on Optic Nerve Neurodegeneration. Int. J. Mol. Sci. 2023, 24, 2573. [Google Scholar] [CrossRef]
- Parekh, N.; Voland, R.P.; Moeller, S.M.; Blodi, B.A.; Ritenbaugh, C.; Chappell, R.J.; Wallace, R.B.; Mares, J.A.; Group, C.R.S. Association between dietary fat intake and age-related macular degeneration in the Carotenoids in Age-Related Eye Disease Study (CAREDS): An ancillary study of the Women’s Health Initiative. Arch. Ophthalmol. 2009, 127, 1483–1493. [Google Scholar] [CrossRef] [PubMed]
- Shiels, A.; Hejtmancik, J.F. Biology of Inherited Cataracts and Opportunities for Treatment. Annu. Rev. Vis. Sci. 2019, 5, 123–149. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Wykoff, C.C.; Khurana, R.N.; Nguyen, Q.D.; Kelly, S.P.; Lum, F.; Hall, R.; Abbass, I.M.; Abolian, A.M.; Stoilov, I.; To, T.M.; et al. Risk of Blindness Among Patients With Diabetes and Newly Diagnosed Diabetic Retinopathy. Diabetes Care 2021, 44, 748–756. [Google Scholar] [CrossRef]
- Khan, J.; Shaw, S. Risk of cataract and glaucoma among older persons with diabetes in India: A cross-sectional study based on LASI, Wave-1. Sci. Rep. 2023, 13, 11973. [Google Scholar] [CrossRef] [PubMed]
- Mathew, D.; Luo, Q.; Bhatwadekar, A.D. Circadian rhythm disruption results in visual dysfunction. FASEB Bioadv. 2022, 4, 364–378. [Google Scholar] [CrossRef] [PubMed]
- Tosini, G. Blue-light-blocking Lenses in Eyeglasses: A Question of Timing. Optom. Vis. Sci. 2022, 99, 228–229. [Google Scholar] [CrossRef]
- Andrews, C.D.; Foster, R.G.; Alexander, I.; Vasudevan, S.; Downes, S.M.; Heneghan, C.; Pluddemann, A. Sleep-Wake Disturbance Related to Ocular Disease: A Systematic Review of Phase-Shifting Pharmaceutical Therapies. Transl. Vis. Sci. Technol. 2019, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Turkiewicz, S.; Karuga, F.F.; Sochal, M.; Strzelecki, D.; Bialasiewicz, P. Disruption of Circadian Rhythm Genes in Obstructive Sleep Apnea Patients-Possible Mechanisms Involved and Clinical Implication. Int. J. Mol. Sci. 2022, 23, 709. [Google Scholar] [CrossRef]
- D’Souza, H.; Kapoor, K.G. Retinal vascular manifestations of obstructive sleep apnea. Curr. Opin. Ophthalmol. 2020, 31, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Mitter, S.K.; Yan, Y.; Busik, J.V.; Grant, M.B.; Boulton, M.E. Diurnal Rhythmicity of Autophagy Is Impaired in the Diabetic Retina. Cells 2020, 9, 905. [Google Scholar] [CrossRef] [PubMed]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin Wound Healing Process and New Emerging Technologies for Skin Wound Care and Regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef] [PubMed]
- Dąbrowska, A.; Spano, F.; Derler, S.; Adlhart, C.; Spencer, N.D.; Rossi, R.M. The relationship between skin function, barrier properties, and body-dependent factors. Skin. Res. Technol. 2018, 24, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Russell-Goldman, E.; Murphy, G.F. The pathobiology of skin aging: New insights into an old dilemma. Am. J. Pathol. 2020, 190, 1356–1369. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.H.; Chien, A.L. Photoaging: A review of current literature. Curr. Dermatol. Rep. 2020, 9, 22–29. [Google Scholar] [CrossRef]
- Pourang, A.; Tisack, A.; Ezekwe, N.; Torres, A.E.; Kohli, I.; Hamzavi, I.H.; Lim, H.W. Effects of visible light on mechanisms of skin photoaging. Photodermatol. Photoimmunol. Photomed. 2022, 38, 191–196. [Google Scholar] [CrossRef]
- Evans, M.; Lewis, E.D.; Zakaria, N.; Pelipyagina, T.; Guthrie, N. A randomized, triple-blind, placebo-controlled, parallel study to evaluate the efficacy of a freshwater marine collagen on skin wrinkles and elasticity. J. Cosmet. Dermatol. 2021, 20, 825–834. [Google Scholar] [CrossRef]
- White-Chu, E.F.; Reddy, M. Dry skin in the elderly: Complexities of a common problem. Clin. Dermatol. 2011, 29, 37–42. [Google Scholar] [CrossRef]
- Lee, A.-Y. Skin pigmentation abnormalities and their possible relationship with skin aging. Int. J. Mol. Sci. 2021, 22, 3727. [Google Scholar] [CrossRef]
- Guida, S.; Pellacani, G.; Ciardo, S.; Longo, C. Reflectance Confocal Microscopy of Aging Skin and Skin Cancer. Dermatol. Pract. Concept. 2021, 11, e2021068. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.J. Introduction to skin aging. J. Tissue Viability 2017, 26, 37–46. [Google Scholar] [CrossRef]
- Benjamin, C.L.; Ananthaswamy, H.N. p53 and the pathogenesis of skin cancer. Toxicol. Appl. Pharmacol. 2007, 224, 241–248. [Google Scholar] [CrossRef]
- Lo, J.A.; Fisher, D.E. The melanoma revolution: From UV carcinogenesis to a new era in therapeutics. Science 2014, 346, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Kirchner, S.J.; Lei, V.; Kim, P.T.; Patel, M.; Shannon, J.L.; Corcoran, D.; Hughes, D.; Waters, D.K.; Dzirasa, K.; Erdmann, D.; et al. An aging-susceptible circadian rhythm controls cutaneous antiviral immunity. JCI Insight 2023, 8, e171548. [Google Scholar] [CrossRef]
- GBD2015. Mortality and Causes of Death Collaborators. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef]
- Madia, F.; Worth, A.; Whelan, M.; Corvi, R. Carcinogenicity assessment: Addressing the challenges of cancer and chemicals in the environment. Environ. Int. 2019, 128, 417–429. [Google Scholar] [CrossRef]
- Brown, J.S.; Amend, S.R.; Austin, R.H.; Gatenby, R.A.; Hammarlund, E.U.; Pienta, K.J. Updating the Definition of Cancer. Mol. Cancer Res. 2023, 21, 1142–1147. [Google Scholar] [CrossRef]
- IARC. Age-Standardized Rate (World) per 100 000, Incidence and Mortality, Both Sexes, in 2022 World, in Cancer Today. Available online: https://gco.iarc.fr/today/en/dataviz/tables?types=0_1&mode=cancer&group_populations=1&sort_by=value1&multiple_populations=1&populations=900&sexes=0 (accessed on 7 June 2024).
- Jung, K.W.; Won, Y.J.; Kang, M.J.; Kong, H.J.; Im, J.S.; Seo, H.G. Prediction of Cancer Incidence and Mortality in Korea, 2022. Cancer Res. Treat. 2022, 54, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.N.; Dave, R.; Sanadya, J.; Sharma, P.; Sharma, K. Various types and management of breast cancer: An overview. J. Adv. Pharm. Technol. Res. 2010, 1, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.J.; Zhu, Z.Y.; Shi, W.; Jiang, J.; Yao, P.P.; Zhu, H.P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Panjarian, S.; Madzo, J.; Keith, K.; Slater, C.M.; Sapienza, C.; Jelinek, J.; Issa, J.J. Accelerated aging in normal breast tissue of women with breast cancer. Breast Cancer Res. 2021, 23, 58. [Google Scholar] [CrossRef]
- Benz, C.C. Impact of aging on the biology of breast cancer. Crit. Rev. Oncol. Hematol. 2008, 66, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Kimmick, G.G.; Balducci, L. Breast cancer and aging. Clinical interactions. Hematol. Oncol. Clin. N. Am. 2000, 14, 213–234. [Google Scholar] [CrossRef]
- Lukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanislawek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- D’Cunha, K.; Park, Y.; Protani, M.M.; Reeves, M.M. Circadian rhythm disrupting behaviours and cancer outcomes in breast cancer survivors: A systematic review. Breast Cancer Res. Treat. 2023, 198, 413–421. [Google Scholar] [CrossRef]
- Ybanez, W.S.; Bagamasbad, P.D. Kruppel-like factor 9 (KLF9) links hormone dysregulation and circadian disruption to breast cancer pathogenesis. Cancer Cell Int. 2023, 23, 33. [Google Scholar] [CrossRef]
- Hammarlund, J.A.; Li, S.Y.; Wu, G.; Lian, J.W.; Howell, S.J.; Clarke, R.; Adamson, A.; Goncalves, C.F.; Hogenesch, J.B.; Meng, Q.J.; et al. Subtype-specific circadian clock dysregulation modulates breast cancer biology, invasiveness, and prognosis. bioRxiv 2023, Preprint. [Google Scholar] [CrossRef]
- Burguin, A.; Diorio, C.; Durocher, F. Breast Cancer Treatments: Updates and New Challenges. J. Pers. Med. 2021, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Schnitt, S.J.; Moran, M.S.; Giuliano, A.E. Lumpectomy Margins for Invasive Breast Cancer and Ductal Carcinoma in Situ: Current Guideline Recommendations, Their Implications, and Impact. J. Clin. Oncol. 2020, 38, 2240–2245. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.J.; Nie, X.Y.; Ji, C.C.; Lin, X.X.; Liu, L.J.; Chen, X.M.; Yao, H.; Wu, S.H. Long-Term Cardiovascular Risk After Radiotherapy in Women With Breast Cancer. J. Am. Heart Assoc. 2017, 6, e005633. [Google Scholar] [CrossRef] [PubMed]
- Chughtai, B.; Forde, J.C.; Thomas, D.D.M.; Laor, L.; Hossack, T.; Woo, H.H.; Te, A.E.; Kaplan, S.A. Benign prostatic hyperplasia. Nat. Rev. Dis. Primers 2016, 2, 16031. [Google Scholar] [CrossRef]
- Langan, R.C. Benign Prostatic Hyperplasia. Prim. Care 2019, 46, 223–232. [Google Scholar] [CrossRef]
- Liu, D.; Shoag, J.E.; Poliak, D.; Goueli, R.S.; Ravikumar, V.; Redmond, D.; Vosoughi, A.; Fontugne, J.; Pan, H.; Lee, D.; et al. Integrative multiplatform molecular profiling of benign prostatic hyperplasia identifies distinct subtypes. Nat. Commun. 2020, 11, 1987. [Google Scholar] [CrossRef]
- Lloyd, G.L.; Marks, J.M.; Ricke, W.A. Benign Prostatic Hyperplasia and Lower Urinary Tract Symptoms: What Is the Role and Significance of Inflammation? Curr. Urol. Rep. 2019, 20, 54. [Google Scholar] [CrossRef] [PubMed]
- Manov, J.J.; Mohan, P.P.; Kava, B.; Bhatia, S. Benign Prostatic Hyperplasia: A Brief Overview of Pathogenesis, Diagnosis, and Current State of Therapy. Tech. Vasc. Interv. Radiol. 2020, 23, 100687. [Google Scholar] [CrossRef]
- Freeland, J.; Crowell, P.D.; Giafaglione, J.M.; Boutros, P.C.; Goldstein, A.S. Aging of the progenitor cells that initiate prostate cancer. Cancer Lett. 2021, 515, 28–35. [Google Scholar] [CrossRef]
- Cooper, C.S.; Eeles, R.; Wedge, D.C.; Van Loo, P.; Gundem, G.; Alexandrov, L.B.; Kremeyer, B.; Butler, A.; Lynch, A.G.; Camacho, N.; et al. Analysis of the genetic phylogeny of multifocal prostate cancer identifies multiple independent clonal expansions in neoplastic and morphologically normal prostate tissue. Nat. Genet. 2015, 47, 367–372. [Google Scholar] [CrossRef]
- Martincorena, I.; Fowler, J.C.; Wabik, A.; Lawson, A.R.J.; Abascal, F.; Hall, M.W.J.; Cagan, A.; Murai, K.; Mahbubani, K.; Stratton, M.R.; et al. Somatic mutant clones colonize the human esophagus with age. Science 2018, 362, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Lee-Six, H.; Olafsson, S.; Ellis, P.; Osborne, R.J.; Sanders, M.A.; Moore, L.; Georgakopoulos, N.; Torrente, F.; Noorani, A.; Goddard, M.; et al. The landscape of somatic mutation in normal colorectal epithelial cells. Nature 2019, 574, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Benayoun, B.A.; Pollina, E.A.; Singh, P.P.; Mahmoudi, S.; Harel, I.; Casey, K.M.; Dulken, B.W.; Kundaje, A.; Brunet, A. Remodeling of epigenome and transcriptome landscapes with aging in mice reveals widespread induction of inflammatory responses. Genome Res. 2019, 29, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Tajuddin, S.M.; Hernandez, D.G.; Chen, B.H.; Noren Hooten, N.; Mode, N.A.; Nalls, M.A.; Singleton, A.B.; Ejiogu, N.; Chitrala, K.N.; Zonderman, A.B.; et al. Novel age-associated DNA methylation changes and epigenetic age acceleration in middle-aged African Americans and whites. Clin. Epigenet. 2019, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.-Z.; He, Q.-Y.; Feng, D.-C.; Wei, Q.; Yang, L. Circadian rhythm in prostate cancer: Time to take notice of the clock. Asian J. Androl. 2023, 25, 184–191. [Google Scholar] [CrossRef]
- Yu, C.-C.; Chen, L.-C.; Chiou, C.-Y.; Chang, Y.-J.; Lin, V.C.; Huang, C.-Y.; Lin, I.L.; Chang, T.-Y.; Lu, T.-L.; Lee, C.-H.; et al. Genetic variants in the circadian rhythm pathway as indicators of prostate cancer progression. Cancer Cell Int. 2019, 19, 87. [Google Scholar] [CrossRef]
- Cao, Q.; Gery, S.; Dashti, A.; Yin, D.; Zhou, Y.; Gu, J.; Koeffler, H.P. A role for the clock gene per1 in prostate cancer. Cancer Res. 2009, 69, 7619–7625. [Google Scholar] [CrossRef]
- Zhu, Y.; Zheng, T.; Stevens, R.G.; Zhang, Y.; Boyle, P. Does “clock” matter in prostate cancer? Cancer Epidemiol. Biomark. Prev. 2006, 15, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Fontana, F.; Raimondi, M.; Marzagalli, M.; Di Domizio, A.; Limonta, P. Natural Compounds in Prostate Cancer Prevention and Treatment: Mechanisms of Action and Molecular Targets. Cells 2020, 9, 460. [Google Scholar] [CrossRef]
- McCaffery, K.; Nickel, B.; Pickles, K.; Moynihan, R.; Kramer, B.; Barratt, A.; Hersch, J. Resisting recommended treatment for prostate cancer: A qualitative analysis of the lived experience of possible overdiagnosis. BMJ Open 2019, 9, e026960. [Google Scholar] [CrossRef]
- Ehsani, M.; Bartsch, S.; Rasa, S.M.M.; Dittmann, J.; Pungsrinont, T.; Neubert, L.; Huettner, S.S.; Kotolloshi, R.; Schindler, K.; Ahmad, A.; et al. The natural compound atraric acid suppresses androgen-regulated neo-angiogenesis of castration-resistant prostate cancer through angiopoietin 2. Oncogene 2022, 41, 3263–3277. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.F.; Jabbarzadeh, E. The use of natural products to target cancer stem cells. Am. J. Cancer Res. 2017, 7, 1588–1605. [Google Scholar] [PubMed]
- Schabath, M.B.; Cote, M.L. Cancer Progress and Priorities: Lung Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Aluru, J.S.; Barsouk, A. Epidemiology of lung cancer. Contemp. Oncol. 2021, 25, 45–52. [Google Scholar] [CrossRef]
- Latimer, K.M. Lung Cancer: Clinical Presentation and Diagnosis. FP Essent. 2018, 464, 23–26. [Google Scholar] [PubMed]
- Lemjabbar-Alaoui, H.; Hassan, O.U.; Yang, Y.-W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta Rev. Cancer 2015, 1856, 189–210. [Google Scholar] [CrossRef]
- Araghi, M.; Mannani, R.; Heidarnejad Maleki, A.; Hamidi, A.; Rostami, S.; Safa, S.H.; Faramarzi, F.; Khorasani, S.; Alimohammadi, M.; Tahmasebi, S.; et al. Recent advances in non-small cell lung cancer targeted therapy; an update review. Cancer Cell Int. 2023, 23, 162. [Google Scholar] [CrossRef] [PubMed]
- Budisan, L.; Zanoaga, O.; Braicu, C.; Pirlog, R.; Covaliu, B.; Esanu, V.; Korban, S.S.; Berindan-Neagoe, I. Links between Infections, Lung Cancer, and the Immune System. Int. J. Mol. Sci. 2021, 22, 9394. [Google Scholar] [CrossRef] [PubMed]
- Edelman, M.J.; Hodgson, L.; Wang, X.; Kratzke, R.A.; Vokes, E.E. Cyclooxygenase-2 (COX-2) as a predictive marker for the use of COX-2 inhibitors in advanced non-small-cell lung cancer. J. Clin. Oncol. 2012, 30, 2019–2020. [Google Scholar] [CrossRef]
- Ridker, P.M.; MacFadyen, J.G.; Thuren, T.; Everett, B.M.; Libby, P.; Glynn, R.J.; Group, C.T. Effect of interleukin-1beta inhibition with canakinumab on incident lung cancer in patients with atherosclerosis: Exploratory results from a randomised, double-blind, placebo-controlled trial. Lancet 2017, 390, 1833–1842. [Google Scholar] [CrossRef]
- Altorki, N.K.; Markowitz, G.J.; Gao, D.; Port, J.L.; Saxena, A.; Stiles, B.; McGraw, T.; Mittal, V. The lung microenvironment: An important regulator of tumour growth and metastasis. Nat. Rev. Cancer 2019, 19, 9–31. [Google Scholar] [CrossRef] [PubMed]
- Malkani, N.; Rashid, M.U. SARS-CoV-2 infection and lung tumor microenvironment. Mol. Biol. Rep. 2021, 48, 1925–1934. [Google Scholar] [CrossRef] [PubMed]
- Parris, B.A.; O’Farrell, H.E.; Fong, K.M.; Yang, I.A. Chronic obstructive pulmonary disease (COPD) and lung cancer: Common pathways for pathogenesis. J. Thorac. Dis. 2019, 11, S2155–S2172. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yuan, M.; Wang, C.; Chen, Y.; Zhang, Y.; Zhang, J. T lymphocyte cell: A pivotal player in lung cancer. Front. Immunol. 2023, 14, 1102778. [Google Scholar] [CrossRef]
- Au, P.C.; Lee, A.W.; Lee, V.H.; Wong, I.C.; Hui, R.Y.; Cheung, C.L. The trends in lung cancer prevalence, incidence, and survival in Hong Kong over the past two decades (2002–2021): A population-based study. Lancet Reg. Health West. Pac. 2024, 45, 101030. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Zhang, S.; Wang, S.; Chen, R.; Sun, K.; Zeng, H.; Li, L.; Wei, W.; He, J. Lung cancer incidence and mortality in China: Updated statistics and an overview of temporal trends from 2000 to 2016. J. Natl. Cancer Cent. 2022, 2, 139–147. [Google Scholar] [CrossRef]
- Chen, S.; Wu, S. Identifying Lung Cancer Risk Factors in the Elderly Using Deep Neural Networks: Quantitative Analysis of Web-Based Survey Data. J. Med. Internet Res. 2020, 22, e17695. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vilanova, A.; Olmo-Fontánez, A.M.; Moliva, J.I.; Allué-Guardia, A.; Singh, H.; Merritt, R.E.; Maselli, D.J.; Peters, J.I.; Restrepo, B.I.; Wang, Y.; et al. The Aging Human Lung Mucosa: A Proteomics Study. J. Gerontol. Ser. A 2022, 77, 1969–1974. [Google Scholar] [CrossRef]
- Xie, L.; Qian, Y.; Liu, Y.; Li, Y.; Jia, S.; Yu, H.; Wang, C.; Qian, B.; Bao, P. Distinctive lung cancer incidence trends among men and women attributable to the period effect in Shanghai: An analysis spanning 42 years. Cancer Med. 2020, 9, 2930–2939. [Google Scholar] [CrossRef]
- May, L.; Shows, K.; Nana-Sinkam, P.; Li, H.; Landry, J.W. Sex Differences in Lung Cancer. Cancers 2023, 15, 3111. [Google Scholar] [CrossRef]
- Novello, S.; Stabile, L.P.; Siegfried, J.M. Gender-Related Differences in Lung Cancer. In IASLC Thoracic Oncology, 2nd ed.; Pass, H.I., Ball, D., Scagliotti, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 30–45.e35. [Google Scholar]
- Pariollaud, M.; Ibrahim, L.H.; Irizarry, E.; Mello, R.M.; Chan, A.B.; Altman, B.J.; Shaw, R.J.; Bollong, M.J.; Wiseman, R.L.; Lamia, K.A. Circadian disruption enhances HSF1 signaling and tumorigenesis in Kras-driven lung cancer. Sci. Adv. 2022, 8, eabo1123. [Google Scholar] [CrossRef]
- Roberts, N.T.; MacDonald, C.R.; Mohammadpour, H.; Antoch, M.P.; Repasky, E.A. Circadian Rhythm Disruption Increases Tumor Growth Rate and Accumulation of Myeloid-Derived Suppressor Cells. Adv. Biol. 2022, 6, e2200031. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Lin, W.H.; Ku, S.C.; Shen, W.J.; Ta, H.D.K.; Anuraga, G.; Liu, F.W.; Shen, C.F.; Wang, S.H.; Yang, C.C.; et al. Circadian rhythm-related factors of PER and CRY family genes function as novel therapeutic targets and prognostic biomarkers in lung adenocarcinoma. Aging 2022, 14, 9056–9089. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; He, T.; Zhu, J.; Wang, X.; Tong, J.; Li, Z.; Dong, J. The Link between Circadian Clock Genes and Autophagy in Chronic Obstructive Pulmonary Disease. Mediat. Inflamm. 2021, 2021, 2689600. [Google Scholar] [CrossRef]
- Hossain, M.S.; Karuniawati, H.; Jairoun, A.A.; Urbi, Z.; Ooi, J.; John, A.; Lim, Y.C.; Kibria, K.M.K.; Mohiuddin, A.K.M.; Ming, L.C.; et al. Colorectal Cancer: A Review of Carcinogenesis, Global Epidemiology, Current Challenges, Risk Factors, Preventive and Treatment Strategies. Cancers 2022, 14, 1732. [Google Scholar] [CrossRef] [PubMed]
- Sabit, H.; Cevik, E.; Tombuloglu, H. Colorectal cancer: The epigenetic role of microbiome. World J. Clin. Cases 2019, 7, 3683–3697. [Google Scholar] [CrossRef]
- Khil, H.; Kim, S.M.; Hong, S.; Gil, H.M.; Cheon, E.; Lee, D.H.; Kim, Y.A.; Keum, N. Time trends of colorectal cancer incidence and associated lifestyle factors in Republic of Korea. Sci. Rep. 2021, 11, 2413. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.L.; Wang, X.B.; Chen, D.D.; Guo, X.J.; Yang, Q.J.; Dong, L.H.; Cheng, L. Germanicol induces selective growth inhibitory effects in human colon HCT-116 and HT29 cancer cells through induction of apoptosis, cell cycle arrest and inhibition of cell migration. J. BUON 2016, 21, 626–632. [Google Scholar]
- Pandurangan, A.K.; Divya, T.; Kumar, K.; Dineshbabu, V.; Velavan, B.; Sudhandiran, G. Colorectal carcinogenesis: Insights into the cell death and signal transduction pathways: A review. World J. Gastrointest. Oncol. 2018, 10, 244–259. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, P.; Sarma, D.K.; Kumawat, M.; Tiwari, R.; Verma, V.; Nagpal, R.; Kumar, M. Implication of Obesity and Gut Microbiome Dysbiosis in the Etiology of Colorectal Cancer. Cancers 2023, 15, 1913. [Google Scholar] [CrossRef]
- Sato, Y.; Tsujinaka, S.; Miura, T.; Kitamura, Y.; Suzuki, H.; Shibata, C. Inflammatory Bowel Disease and Colorectal Cancer: Epidemiology, Etiology, Surveillance, and Management. Cancers 2023, 15, 4154. [Google Scholar] [CrossRef]
- Dougherty, M.W.; Jobin, C. Intestinal bacteria and colorectal cancer: Etiology and treatment. Gut Microbes 2023, 15, 2185028. [Google Scholar] [CrossRef]
- Medici, B.; Ricco, B.; Caffari, E.; Zaniboni, S.; Salati, M.; Spallanzani, A.; Garajova, I.; Benatti, S.; Chiavelli, C.; Dominici, M.; et al. Early Onset Metastatic Colorectal Cancer: Current Insights and Clinical Management of a Rising Condition. Cancers 2023, 15, 3509. [Google Scholar] [CrossRef]
- Ciardiello, F.; Ciardiello, D.; Martini, G.; Napolitano, S.; Tabernero, J.; Cervantes, A. Clinical management of metastatic colorectal cancer in the era of precision medicine. CA Cancer J. Clin. 2022, 72, 372–401. [Google Scholar] [CrossRef]
- Thanikachalam, K.; Khan, G. Colorectal Cancer and Nutrition. Nutrients 2019, 11, 164. [Google Scholar] [CrossRef]
- Wang, Y.; Narasimamurthy, R.; Qu, M.; Shi, N.; Guo, H.; Xue, Y.; Barker, N. Circadian regulation of cancer stem cells and the tumor microenvironment during metastasis. Nat. Cancer 2024, 5, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Dalerba, P.; Dylla, S.J.; Park, I.K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef]
- Stokes, K.; Nunes, M.; Trombley, C.; Flôres, D.; Wu, G.; Taleb, Z.; Alkhateeb, A.; Banskota, S.; Harris, C.; Love, O.P.; et al. The Circadian Clock Gene, Bmal1, Regulates Intestinal Stem Cell Signaling and Represses Tumor Initiation. Cell Mol. Gastroenterol. Hepatol. 2021, 12, 1847–1872.e0. [Google Scholar] [CrossRef]
- Zhang, Y.; Devocelle, A.; Souza, L.; Foudi, A.; Tenreira Bento, S.; Desterke, C.; Sherrard, R.; Ballesta, A.; Adam, R.; Giron-Michel, J.; et al. BMAL1 knockdown triggers different colon carcinoma cell fates by altering the delicate equilibrium between AKT/mTOR and P53/P21 pathways. Aging 2020, 12, 8067–8083. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Wang, C.Y.; Cheng, T.Y.; Rixiati, Y.; Ji, C.; Deng, M.; Yao, S.; Yuan, L.H.; Zhao, Y.Y.; Shen, T.; et al. Circadian Clock Disruption Suppresses PDL1(+) Intraepithelial B Cells in Experimental Colitis and Colitis-Associated Colorectal Cancer. Cell Mol. Gastroenterol. Hepatol. 2021, 12, 251–276. [Google Scholar] [CrossRef] [PubMed]
- Biller, L.H.; Schrag, D. A Review of the Diagnosis and Treatment of Metastatic Colorectal Cancer-Reply. JAMA 2021, 325, 2405. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, F.; Khaderi, S.; Singal, A.G.; Marrero, J.A.; Loo, N.; Asrani, S.K.; Amos, C.I.; Thrift, A.P.; Gu, X.; Luster, M.; et al. Risk factors for HCC in contemporary cohorts of patients with cirrhosis. Hepatology 2023, 77, 997–1005. [Google Scholar] [CrossRef]
- Jeng, K.S.; Chang, C.F.; Sheen, I.S.; Jeng, C.J.; Wang, C.H. Upper Gastrointestinal Cancer and Liver Cirrhosis. Cancers 2022, 14, 2269. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, N.S.; Kumar, N.; Duseja, A. Peroxisome Proliferator-Activated Receptors and Their Agonists in Nonalcoholic Fatty Liver Disease. J. Clin. Exp. Hepatol. 2019, 9, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, R.; Mitra, A. An overview of effective therapies and recent advances in biomarkers for chronic liver diseases and associated liver cancer. Int. Immunopharmacol. 2015, 24, 335–345. [Google Scholar] [CrossRef]
- Flores, J.E.; Thompson, A.J.; Ryan, M.; Howell, J. The Global Impact of Hepatitis B Vaccination on Hepatocellular Carcinoma. Vaccines 2022, 10, 793. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Singal, A.G.; El-Serag, H.B. Hepatocellular Carcinoma From Epidemiology to Prevention: Translating Knowledge into Practice. Clin. Gastroenterol. Hepatol. 2015, 13, 2140–2151. [Google Scholar] [CrossRef]
- Yang, L.N.; Xu, S.; Tang, M.; Zhou, X.; Liao, Y.; Nussler, A.K.; Liu, L.; Yang, W. The circadian rhythm gene Bmal1 ameliorates acute deoxynivalenol-induced liver damage. Arch. Toxicol. 2023, 97, 787–804. [Google Scholar] [CrossRef]
- Guan, D.; Xiong, Y.; Trinh, T.M.; Xiao, Y.; Hu, W.; Jiang, C.; Dierickx, P.; Jang, C.; Rabinowitz, J.D.; Lazar, M.A. The hepatocyte clock and feeding control chronophysiology of multiple liver cell types. Science 2020, 369, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Scheuermaier, K.; Chang, A.M.; Duffy, J.F. Sleep-independent circadian rhythm of aldosterone secretion in healthy young adults. Sleep. Health 2024, 10, S103–S107. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gan, L.; Luo, D.; Sun, C. Melatonin promotes circadian rhythm-induced proliferation through Clock/histone deacetylase 3/c-Myc interaction in mouse adipose tissue. J. Pineal Res. 2017, 62, e12383. [Google Scholar] [CrossRef]
- Yasmin, A.; Regan, D.P.; Schook, L.B.; Gaba, R.C.; Schachtschneider, K.M. Transcriptional regulation of alcohol induced liver fibrosis in a translational porcine hepatocellular carcinoma model. Biochimie 2021, 182, 73–84. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, W.; Yang, Y.; He, M.; Li, A.; Bai, L.; Yu, B.; Yu, Z. Cancer nanotechnology: Enhancing tumor cell response to chemotherapy for hepatocellular carcinoma therapy. Asian J. Pharm. Sci. 2019, 14, 581–594. [Google Scholar] [CrossRef]
- Anwanwan, D.; Singh, S.K.; Singh, S.; Saikam, V.; Singh, R. Challenges in liver cancer and possible treatment approaches. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188314. [Google Scholar] [CrossRef] [PubMed]
- Man, K.F.; Ma, S. Mechanisms of resistance to tyrosine kinase inhibitors in liver cancer stem cells and potential therapeutic approaches. Essays Biochem. 2022, 66, 371–386. [Google Scholar] [CrossRef]
- Yang, Z.; Qi, Y.; Lai, N.; Zhang, J.; Chen, Z.; Liu, M.; Zhang, W.; Luo, R.; Kang, S. Notch1 signaling in melanoma cells promoted tumor-induced immunosuppression via upregulation of TGF-β1. J. Exp. Clin. Cancer Res. 2018, 37, 1. [Google Scholar] [CrossRef]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- WCRF. Stomach Cancer Statistics. Available online: https://www.wcrf.org/cancer-trends/stomach-cancer-statistics/ (accessed on 15 April 2024).
- Islami, F.; DeSantis, C.E.; Jemal, A. Incidence Trends of Esophageal and Gastric Cancer Subtypes by Race, Ethnicity, and Age in the United States, 1997–2014. Clin. Gastroenterol. Hepatol. 2019, 17, 429–439. [Google Scholar] [CrossRef]
- Suarez, F.; Lecuit, M. Infection-associated non-Hodgkin lymphomas. Clin. Microbiol. Infect. 2015, 21, 991–997. [Google Scholar] [CrossRef]
- Lai, Y.; Shi, H.; Wang, Z.; Feng, Y.; Bao, Y.; Li, Y.; Li, J.; Wu, A. Incidence trends and disparities in Helicobacter pylori related malignancy among US adults, 2000–2019. Front. Public Health 2022, 10, 1056157. [Google Scholar] [CrossRef]
- Ning, F.L.; Zhang, N.N.; Zhao, Z.M.; Du, W.Y.; Zeng, Y.J.; Abe, M.; Pei, J.P.; Zhang, C.D. Global, Regional, and National Burdens with Temporal Trends of Early-, Intermediate-, and Later-Onset Gastric Cancer from 1990 to 2019 and Predictions up to 2035. Cancers 2022, 14, 5417. [Google Scholar] [CrossRef]
- Umesawa, M.; Iso, H.; Fujino, Y.; Kikuchi, S.; Tamakoshi, A.; Group, J.S. Salty Food Preference and Intake and Risk of Gastric Cancer: The JACC Study. J. Epidemiol. 2016, 26, 92–97. [Google Scholar] [CrossRef]
- Butt, J.; Varga, M.G.; Wang, T.; Tsugane, S.; Shimazu, T.; Zheng, W.; Abnet, C.C.; Yoo, K.Y.; Park, S.K.; Kim, J.; et al. Smoking, Helicobacter Pylori Serology, and Gastric Cancer Risk in Prospective Studies from China, Japan, and Korea. Cancer Prev. Res. 2019, 12, 667–674. [Google Scholar] [CrossRef]
- Na, H.K.; Lee, J.Y. Molecular Basis of Alcohol-Related Gastric and Colon Cancer. Int. J. Mol. Sci. 2017, 18, 1116. [Google Scholar] [CrossRef]
- Anderson, W.F.; Rabkin, C.S.; Turner, N.; Fraumeni, J.F., Jr.; Rosenberg, P.S.; Camargo, M.C. The Changing Face of Noncardia Gastric Cancer Incidence Among US Non-Hispanic Whites. J. Natl. Cancer Inst. 2018, 110, 608–615. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, N.; Kim, K.W.; Jo, H.H.; Park, J.; Yoon, H.; Shin, C.M.; Park, Y.S.; Lee, D.H. Gastric Cancer in Older Patients: A Retrospective Study and Literature Review. Ann. Geriatr. Med. Res. 2022, 26, 33–41. [Google Scholar] [CrossRef]
- Samartzis, E.; Noske, A.; Dedes, K.; Fink, D.; Imesch, P. ARID1A Mutations and PI3K/AKT Pathway Alterations in Endometriosis and Endometriosis-Associated Ovarian Carcinomas. Int. J. Mol. Sci. 2013, 14, 18824–18849. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Mao, Z.; Zhou, X.; Zou, Y.; Zhao, M. Construction and validation of a novel prognostic model using the cellular senescence-associated long non-coding RNA in gastric cancer: A biological analysis. J. Gastrointest. Oncol. 2022, 13, 1640–1655. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yin, X.; Fang, T.; Lou, S.; Han, B.; Gao, J.; Wang, Y.; Zhang, D.; Wang, X.; Lu, Z.; et al. Development and Validation of an Age-Related Gastric Cancer-Specific Immune Index. J. Inflamm. Res. 2022, 15, 6393–6407. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, W.; Zheng, S.; Wang, L.; Zhang, L.; Pei, X. Construction of an immune-related risk score signature for gastric cancer based on multi-omics data. Sci. Rep. 2024, 14, 1422. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.K.; Ma, H.; Chong, R.H.H.; Uchihara, T.; Lian, B.S.X.; Zhu, F.; Sheng, T.; Srivastava, S.; Tay, S.T.; Sundar, R.; et al. Spatiotemporal genomic profiling of intestinal metaplasia reveals clonal dynamics of gastric cancer progression. Cancer Cell 2023, 41, 2019–2037.e8. [Google Scholar] [CrossRef] [PubMed]
- Sugano, K.; Moss, S.F.; Kuipers, E.J. Gastric Intestinal Metaplasia: Real Culprit or Innocent Bystander as a Precancerous Condition for Gastric Cancer? Gastroenterology 2023, 165, 1352–1366.e1. [Google Scholar] [CrossRef] [PubMed]
- Ki, M.R.; Hwang, M.; Kim, A.Y.; Lee, E.M.; Lee, E.J.; Lee, M.M.; Sung, S.E.; Kim, S.H.; Lee, H.S.; Jeong, K.S. Role of vacuolating cytotoxin VacA and cytotoxin-associated antigen CagA of Helicobacter pylori in the progression of gastric cancer. Mol. Cell Biochem. 2014, 396, 23–32. [Google Scholar] [CrossRef]
- Cui, S.; Chen, Y.; Guo, Y.; Chen, D. Clock genes and gastric cancer. Biol. Rhythm. Res. 2022, 53, 1720–1734. [Google Scholar] [CrossRef]
- Zheng, Z.X.; Cai, X.; Bi, J.T.; Liu, Y.Q. The expression and prognostic significance of circadian gene NR1D1/2 in patients with gastric cancer. Res. Sq. 2023, preprint. [Google Scholar] [CrossRef]
- Noh, S.G.; Jung, H.J.; Kim, S.; Arulkumar, R.; Kim, D.H.; Park, D.; Chung, H.Y. Regulation of Circadian Genes Nr1d1 and Nr1d2 in Sex-Different Manners during Liver Aging. Int. J. Mol. Sci. 2022, 23, 10032. [Google Scholar] [CrossRef]
- Wang, S.; Khan, S.; Nabi, G.; Li, H.Y. Circadian rhythm as a key player in cancer progression as well as a therapeutic target in HER2-positive advanced gastric cancer treatment. Front. Oncol. 2023, 13, 1240676. [Google Scholar] [CrossRef]
- Gillen, S.; Schuster, T.; Meyer Zum Buschenfelde, C.; Friess, H.; Kleeff, J. Preoperative/neoadjuvant therapy in pancreatic cancer: A systematic review and meta-analysis of response and resection percentages. PLoS Med. 2010, 7, e1000267. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, W. Pancreatic Cancer: A Review of Risk Factors, Diagnosis, and Treatment. Technol. Cancer Res. Treat. 2020, 19, 1533033820962117. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.W.; O’Reilly, E.M. Next-generation therapies for pancreatic cancer. Expert. Rev. Gastroenterol. Hepatol. 2024, 18, 55–72. [Google Scholar] [CrossRef]
- Reshkin, S.J.; Cardone, R.A.; Koltai, T. Genetic Signature of Human Pancreatic Cancer and Personalized Targeting. Cells 2024, 13, 602. [Google Scholar] [CrossRef]
- Schwartz, P.B.; Nukaya, M.; Berres, M.E.; Rubinstein, C.D.; Wu, G.; Hogenesch, J.B.; Bradfield, C.A.; Ronnekleiv-Kelly, S.M. The circadian clock is disrupted in pancreatic cancer. PLoS Genet. 2023, 19, e1010770. [Google Scholar] [CrossRef]
- Wu, J.; Jing, X.; Du, Q.; Sun, X.; Holgersson, K.; Gao, J.; He, X.; Hosaka, K.; Zhao, C.; Tao, W.; et al. Disruption of the Clock Component Bmal1 in Mice Promotes Cancer Metastasis through the PAI-1-TGF-beta-myoCAF-Dependent Mechanism. Adv. Sci. 2023, 10, e2301505. [Google Scholar] [CrossRef]
- Schwartz, P.B.; Walcheck, M.T.; Nukaya, M.; Pavelec, D.M.; Matkowskyj, K.A.; Ronnekleiv-Kelly, S.M. Chronic jetlag accelerates pancreatic neoplasia in conditional Kras-mutant mice. Chronobiol. Int. 2023, 40, 417–437. [Google Scholar] [CrossRef]
- Rodriguez-Negrete, E.V.; Morales-Gonzalez, A.; Madrigal-Santillan, E.O.; Sanchez-Reyes, K.; Alvarez-Gonzalez, I.; Madrigal-Bujaidar, E.; Valadez-Vega, C.; Chamorro-Cevallos, G.; Garcia-Melo, L.F.; Morales-Gonzalez, J.A. Phytochemicals and Their Usefulness in the Maintenance of Health. Plants 2024, 13, 523. [Google Scholar] [CrossRef]
- Kumar, A.; P, N.; Kumar, M.; Jose, A.; Tomer, V.; Oz, E.; Proestos, C.; Zeng, M.; Elobeid, T.; K, S.; et al. Major Phytochemicals: Recent Advances in Health Benefits and Extraction Method. Molecules 2023, 28, 887. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Huang, L.; Huang, H.; Liu, S.; Dai, W.; Tang, J.; Chen, X.; Lu, X.; Zheng, Q.; Zhou, Z.; et al. Bioactivities and Mechanisms of Action of Sinomenine and Its Derivatives: A Comprehensive Review. Molecules 2024, 29, 540. [Google Scholar] [CrossRef]
- Shim, K.H.; Kang, M.J.; Sharma, N.; An, S.S.A. Beauty of the beast: Anticholinergic tropane alkaloids in therapeutics. Nat. Prod. Bioprospect. 2022, 12, 33. [Google Scholar] [CrossRef]
- Cao, L.H.; Li, H.T.; Lin, W.Q.; Tan, H.Y.; Xie, L.; Zhong, Z.J.; Zhou, J.H. Morphine, a potential antagonist of cisplatin cytotoxicity, inhibits cisplatin-induced apoptosis and suppression of tumor growth in nasopharyngeal carcinoma xenografts. Sci. Rep. 2016, 6, 18706. [Google Scholar] [CrossRef]
- Van Cleemput, M.; Cattoor, K.; De Bosscher, K.; Haegeman, G.; De Keukeleire, D.; Heyerick, A. Hop (Humulus lupulus)-derived bitter acids as multipotent bioactive compounds. J. Nat. Prod. 2009, 72, 1220–1230. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Ji, R.; Qiao, G.; Sun, Z.; Chen, X.; Zhang, Z. Amarogentin inhibits vascular smooth muscle cell proliferation and migration and attenuates neointimal hyperplasia via AMPK activation. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166667. [Google Scholar] [CrossRef] [PubMed]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Abdoul-Latif, F.; Ainane, A.; Houmed Aboubaker, I.; Mohamed, J.; Ainane, T. Exploring the Potent Anticancer Activity of Essential Oils and Their Bioactive Compounds: Mechanisms and Prospects for Future Cancer Therapy. Pharmaceuticals 2023, 16, 1086. [Google Scholar] [CrossRef] [PubMed]
- NIH. Omega-3 Fatty Acids. Available online: https://ods.od.nih.gov/factsheets/Omega3FattyAcids-Consumer/ (accessed on 18 June 2024).
- Kumari, A.; Pabbi, S.; Tyagi, A. Recent advances in enhancing the production of long chain omega-3 fatty acids in microalgae. Crit. Rev. Food Sci. Nutr. 2023, 1–19. [Google Scholar] [CrossRef] [PubMed]
- USDA. Active Plant Ingredients Used for Medicinal Purposes. Available online: https://www.fs.usda.gov/wildflowers/ethnobotany/medicinal/ingredients.shtml (accessed on 19 April 2024).
- Kosuru, R.; Cai, Y.; Tiwari, V. Editorial: Natural products targeting oxidative stress and cell death: Treatment potential in metabolic and cardiovascular diseases. Front. Pharmacol. 2023, 14, 1141878. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Chaudhuri, P.K. Structural characteristics, bioavailability and cardioprotective potential of saponins. Integr. Med. Res. 2018, 7, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, Q.; Yang, Y.; Ke, Z.; Chen, S.; Li, M.; Fan, W.; Wu, H.; Yuan, J.; Wang, Z.; et al. Cardioprotective Effect of Stem-Leaf Saponins From Panax notoginseng on Mice With Sleep Derivation by Inhibiting Abnormal Autophagy Through PI3K/Akt/mTOR Pathway. Front. Cardiovasc. Med. 2021, 8, 694219. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Lv, Q.; Liu, X.; Ye, X.; Cao, L.; Wang, M.; Li, J.; Yang, Y.; Li, L.; Wang, S. Natural compounds from botanical drugs targeting mTOR signaling pathway as promising therapeutics for atherosclerosis: A review. Front. Pharmacol. 2023, 14, 1083875. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Sanches-Silva, A.; Testai, L.; Nabavi, S.F.; Battino, M.; Pandima Devi, K.; Tejada, S.; Sureda, A.; Xu, S.; Yousefi, B.; Majidinia, M.; et al. Therapeutic potential of polyphenols in cardiovascular diseases: Regulation of mTOR signaling pathway. Pharmacol. Res. 2020, 152, 104626. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zheng, Y.; Yan, F.; Dong, M.; Ren, Y. Research progress of quercetin in cardiovascular disease. Front. Cardiovasc. Med. 2023, 10, 1203713. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.-D.; Chen, L.-J.; Xu, X.-Y.; Liu, Y.-J.; Tao, F.; Zhu, M.-H.; Li, C.-Y.; Zhao, D.; Yang, G.-J.; Chen, J. Berberine as a potential agent for breast cancer therapy. Front. Oncol. 2022, 12, 993775. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Sureda, A.; Jafari, S.; Memariani, Z.; Tewari, D.; Annunziata, G.; Barrea, L.; Hassan, S.T.S.; Smejkal, K.; Malanik, M.; et al. Berberine in Cardiovascular and Metabolic Diseases: From Mechanisms to Therapeutics. Theranostics 2019, 9, 1923–1951. [Google Scholar] [CrossRef] [PubMed]
- Ooi, B.K.; Chan, K.G.; Goh, B.H.; Yap, W.H. The Role of Natural Products in Targeting Cardiovascular Diseases via Nrf2 Pathway: Novel Molecular Mechanisms and Therapeutic Approaches. Front. Pharmacol. 2018, 9, 1308. [Google Scholar] [CrossRef]
- Nelson, S.K.; Bose, S.K.; Grunwald, G.K.; Myhill, P.; McCord, J.M. The induction of human superoxide dismutase and catalase in vivo: A fundamentally new approach to antioxidant therapy. Free Radic. Biol. Med. 2006, 40, 341–347. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Suarez, M.; Aragones, G.; Mulero, M.; Avila-Roman, J.; Arola-Arnal, A.; Salvado, M.J.; Arola, L.; Bravo, F.I.; Muguerza, B. Cardioprotective Properties of Phenolic Compounds: A Role for Biological Rhythms. Mol. Nutr. Food Res. 2022, 66, e2100990. [Google Scholar] [CrossRef]
- Han, C.; Wirianto, M.; Kim, E.; Burish, M.J.; Yoo, S.H.; Chen, Z. Clock-Modulating Activities of the Anti-Arrhythmic Drug Moricizine. Clocks Sleep. 2021, 3, 351–365. [Google Scholar] [CrossRef]
- Xu, T.; Lu, B. The effects of phytochemicals on circadian rhythm and related diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Kratz, E.M.; Solkiewicz, K.; Kubis-Kubiak, A.; Piwowar, A. Sirtuins as Important Factors in Pathological States and the Role of Their Molecular Activity Modulators. Int. J. Mol. Sci. 2021, 22, 630. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, C.R.; Schaefer, C.A.; Kosok, C.; Abdallah, Y.; Walther, S.; Ludders, D.W.; Neumann, T.; Tillmanns, H.; Schafer, C.; Piper, H.M.; et al. Quercetin-induced induction of the NO/cGMP pathway depends on Ca2+-activated K+ channel-induced hyperpolarization-mediated Ca2+-entry into cultured human endothelial cells. Planta Med. 2005, 71, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, M.; Moreno, L.; Vera, R.; Cogolludo, A.; Duarte, J.; Tamargo, J.; Perez-Vizcaino, F. Effects of the flavonoid quercetin and its methylated metabolite isorhamnetin in isolated arteries from spontaneously hypertensive rats. Planta Med. 2003, 69, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, G.; Liu, R.; Luo, L.; Zhang, Y.; Wan, Z. Quercetin improved hepatic circadian rhythm dysfunction in middle-aged mice fed with vitamin D-deficient diet. J. Physiol. Biochem. 2024, 80, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Long, H.Z.; Cheng, Y.; Zhou, Z.W.; Luo, H.Y.; Wen, D.D.; Gao, L.C. PI3K/AKT Signal Pathway: A Target of Natural Products in the Prevention and Treatment of Alzheimer’s Disease and Parkinson’s Disease. Front. Pharmacol. 2021, 12, 648636. [Google Scholar] [CrossRef] [PubMed]
- Benameur, T.; Giacomucci, G.; Panaro, M.A.; Ruggiero, M.; Trotta, T.; Monda, V.; Pizzolorusso, I.; Lofrumento, D.D.; Porro, C.; Messina, G. New Promising Therapeutic Avenues of Curcumin in Brain Diseases. Molecules 2021, 27, 236. [Google Scholar] [CrossRef] [PubMed]
- Limanaqi, F.; Biagioni, F.; Busceti, C.L.; Ryskalin, L.; Polzella, M.; Frati, A.; Fornai, F. Phytochemicals Bridging Autophagy Induction and Alpha-Synuclein Degradation in Parkinsonism. Int. J. Mol. Sci. 2019, 20, 3274. [Google Scholar] [CrossRef]
- Park, K.; Lee, M.S. Current Status of Autophagy Enhancers in Metabolic Disorders and Other Diseases. Front. Cell Dev. Biol. 2022, 10, 811701. [Google Scholar] [CrossRef]
- Tian, E.; Sharma, G.; Dai, C. Neuroprotective Properties of Berberine: Molecular Mechanisms and Clinical Implications. Antioxidants 2023, 12, 1883. [Google Scholar] [CrossRef]
- Cui, H.S.; Li, Y.M.; Fang, W.; Li, J.K.; Dai, Y.M.; Zheng, L.S. Effect of berberine on lipopolysaccharide-induced monocyte chemotactic protein-1 and interleukin-8 expression in a human retinal pigment epithelial cell line. Int. Ophthalmol. 2018, 38, 2053–2060. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.Y.; Tang, C.H.; Chen, Y.H.; Wei, I.H. Berberine suppresses neuroinflammatory responses through AMP-activated protein kinase activation in BV-2 microglia. J. Cell Biochem. 2010, 110, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Domanowska, A.; Wódkiewicz, E.; Malinowski, B. Neuroprotective Properties of Resveratrol and Its Derivatives-Influence on Potential Mechanisms Leading to the Development of Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21, 2749. [Google Scholar] [CrossRef]
- Wiciński, M.; Socha, M.; Walczak, M.; Wódkiewicz, E.; Malinowski, B.; Rewerski, S.; Górski, K.; Pawlak-Osińska, K. Beneficial Effects of Resveratrol Administration-Focus on Potential Biochemical Mechanisms in Cardiovascular Conditions. Nutrients 2018, 10, 1813. [Google Scholar] [CrossRef] [PubMed]
- Donmez, G.; Outeiro, T.F. SIRT1 and SIRT2: Emerging targets in neurodegeneration. EMBO Mol. Med. 2013, 5, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Q.; Cheng, X.; Li, X.; Li, N.; Liu, T.; Li, J.; Yang, Q.; Dong, R.; Zhang, Y.; et al. Inhibitive Effect of Resveratrol on the Inflammation in Cultured Astrocytes and Microglia Induced by Aβ1–42. Neuroscience 2018, 379, 390–404. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Shang, L.; Liao, Z.; Su, H.; Jing, H.; Wu, B.; Bi, K.; Jia, Y. Intracerebroventricular injection of resveratrol ameliorated Aβ-induced learning and cognitive decline in mice. Metab. Brain Dis. 2019, 34, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Ide, K.; Matsuoka, N.; Yamada, H.; Furushima, D.; Kawakami, K. Effects of Tea Catechins on Alzheimer’s Disease: Recent Updates and Perspectives. Molecules 2018, 23, 2357. [Google Scholar] [CrossRef]
- Noori, T.; Dehpour, A.R.; Sureda, A.; Sobarzo-Sanchez, E.; Shirooie, S. Role of natural products for the treatment of Alzheimer’s disease. Eur. J. Pharmacol. 2021, 898, 173974. [Google Scholar] [CrossRef]
- Singh, N.A.; Bhardwaj, V.; Ravi, C.; Ramesh, N.; Mandal, A.K.A.; Khan, Z.A. EGCG nanoparticles attenuate aluminum chloride induced neurobehavioral deficits, beta amyloid and tau pathology in a rat model of Alzheimer’s disease. Front. Aging Neurosci. 2018, 10, 244. [Google Scholar] [CrossRef]
- Guéroux, M.; Fleau, C.; Slozeck, M.; Laguerre, M.; Pianet, I. Epigallocatechin 3-Gallate as an Inhibitor of Tau Phosphorylation and Aggregation: A Molecular and Structural Insight. J. Prev. Alzheimer’s Dis. 2017, 4, 218–225. [Google Scholar] [CrossRef]
- Kim, E.; Nohara, K.; Wirianto, M.; Escobedo, G.; Lim, J.Y.; Morales, R.; Yoo, S.-H.; Chen, Z. Effects of the Clock Modulator Nobiletin on Circadian Rhythms and Pathophysiology in Female Mice of an Alzheimer’s Disease Model. Biomolecules 2021, 11, 1004. [Google Scholar] [CrossRef] [PubMed]
- Wirianto, M.; Wang, C.Y.; Kim, E.; Koike, N.; Gomez-Gutierrez, R.; Nohara, K.; Escobedo, G.; Choi, J.M.; Han, C.; Yagita, K.; et al. The clock modulator Nobiletin mitigates astrogliosis-associated neuroinflammation and disease hallmarks in an Alzheimer’s disease model. FASEB J. 2022, 36, e22186. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W. Neuroprotective effects of melatonin in neurodegenerative and autoimmune central nervous system diseases. Encephalitis 2023, 3, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P. Melatonin: Clinical Perspectives in Neurodegeneration. Front. Endocrinol. 2019, 10, 480. [Google Scholar] [CrossRef] [PubMed]
- Avram, S.; Puia, A.; Udrea, A.M.; Mihailescu, D.; Mernea, M.; Dinischiotu, A.; Oancea, F.; Stiens, J. Natural Compounds Therapeutic Features in Brain Disorders by Experimental, Bioinformatics and Cheminformatics Methods. Curr. Med. Chem. 2020, 27, 78–98. [Google Scholar] [CrossRef] [PubMed]
- Jagota, A.; Kowshik, K. Therapeutic Effects of Ashwagandha in Brain Aging and Clock Dysfunction. In Science of Ashwagandha: Preventive and Therapeutic Potentials; Springer: Cham, Switzerland, 2017; pp. 437–456. [Google Scholar]
- Fernandez-Fernandez, A.M.; Iriondo-DeHond, A.; Nardin, T.; Larcher, R.; Dellacassa, E.; Medrano-Fernandez, A.; Castillo, M.D.D. In Vitro Bioaccessibility of Extractable Compounds from Tannat Grape Skin Possessing Health Promoting Properties with Potential to Reduce the Risk of Diabetes. Foods 2020, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Su, F.; Wang, G.; Peng, Z.; Xu, Y.; Zhang, Y.; Xu, N.; Hou, K.; Hu, Z.; Chen, Y.; et al. Glucose-lowering effect of berberine on type 2 diabetes: A systematic review and meta-analysis. Front. Pharmacol. 2022, 13, 1015045. [Google Scholar] [CrossRef]
- Guo, J.; Chen, H.; Zhang, X.; Lou, W.; Zhang, P.; Qiu, Y.; Zhang, C.; Wang, Y.; Liu, W.J. The Effect of Berberine on Metabolic Profiles in Type 2 Diabetic Patients: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Oxid. Med. Cell Longev. 2021, 2021, 2074610. [Google Scholar] [CrossRef]
- Zhou, R.; Xiang, C.; Cao, G.; Xu, H.; Zhang, Y.; Yang, H.; Zhang, J. Berberine accelerated wound healing by restoring TrxR1/JNK in diabetes. Clin Sci 2021, 135, 613–627. [Google Scholar] [CrossRef]
- Bian, W.; Zhang, W.; Liang, H.; Xie, X.; Lai, L. Silybin A enhances circadian clock by targeting CRY1 and disrupting its interaction with CLOCK. Pharmacol. Res. Mod. Chin. Med. 2022, 5, 100159. [Google Scholar] [CrossRef]
- Marcheva, B.; Weidemann, B.J.; Taguchi, A.; Perelis, M.; Ramsey, K.M.; Newman, M.V.; Kobayashi, Y.; Omura, C.; Manning Fox, J.E.; Lin, H.; et al. P2Y1 purinergic receptor identified as a diabetes target in a small-molecule screen to reverse circadian beta-cell failure. Elife 2022, 11, 75132. [Google Scholar] [CrossRef] [PubMed]
- Nitulescu, G.M.; Van De Venter, M.; Nitulescu, G.; Ungurianu, A.; Juzenas, P.; Peng, Q.; Olaru, O.T.; Gradinaru, D.; Tsatsakis, A.; Tsoukalas, D.; et al. The Akt pathway in oncology therapy and beyond (Review). Int. J. Oncol. 2018, 53, 2319–2331. [Google Scholar] [CrossRef]
- Wajid, F.; Poolacherla, R.; Mim, F.K.; Bangash, A.; Rutkofsky, I.H. Therapeutic potential of melatonin as a chronobiotic and cytoprotective agent in diabetes mellitus. J. Diabetes Metab. Disord. 2020, 19, 1797–1825. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, S.; Lin, S.; Lv, Z. Melatonin ameliorates diabetic hyperglycaemia-induced impairment of Leydig cell steroidogenic function through activation of SIRT1 pathway. Reprod. Biol. Endocrinol. 2022, 20, 117. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Robertson, W.B.; Zhao, J.; Chen, W.; Xu, J. Emerging Trend in the Pharmacotherapy of Osteoarthritis. Front Endocrinol 2019, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Crofford, L.J. Use of NSAIDs in treating patients with arthritis. Arthritis Res. Ther. 2013, 15 (Suppl. S3), S2. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zheng, Y.; Chen, X. Drugs for Autoimmune Inflammatory Diseases: From Small Molecule Compounds to Anti-TNF Biologics. Front. Pharmacol. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Zahan, O.M.; Serban, O.; Gherman, C.; Fodor, D. The evaluation of oxidative stress in osteoarthritis. Med. Pharm. Rep. 2020, 93, 12–22. [Google Scholar] [CrossRef]
- Nisar, A.; Jagtap, S.; Vyavahare, S.; Deshpande, M.; Harsulkar, A.; Ranjekar, P.; Prakash, O. Phytochemicals in the treatment of inflammation-associated diseases: The journey from preclinical trials to clinical practice. Front. Pharmacol. 2023, 14, 1177050. [Google Scholar] [CrossRef]
- Aloke, C.; Ohanenye, I.C.; Aja, P.M.; Ejike, C.E.C.C. Phytochemicals from medicinal plants from African forests with potentials in rheumatoid arthritis management. J. Pharm. Pharmacol. 2022, 74, 1205–1219. [Google Scholar] [CrossRef]
- Zhang, L.; Li, T.; Wang, R.; Xu, J.; Zhou, L.; Yan, L.; Hu, Z.; Li, H.; Liu, F.; Du, W.; et al. Evaluation of Long-Time Decoction-Detoxicated Hei-Shun-Pian (Processed Aconitum carmichaeli Debeaux Lateral Root With Peel) for Its Acute Toxicity and Therapeutic Effect on Mono-Iodoacetate Induced Osteoarthritis. Front. Pharmacol. 2020, 11, 1053. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Yunus, M.H.M.; Ugusman, A.; Yazid, M.D. Natural Compounds Affecting Inflammatory Pathways of Osteoarthritis. Antioxidants 2022, 11, 1722. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Mandal, S.K.; Akhtar, M.S.; Dastider, D.; Sarkar, S.; Bose, S.; Bose, A.; Mandal, S.; Kolay, A.; Sen, D.J.; et al. Phytochemicals in the Treatment of Arthritis: Current Knowledge. Int. J. Curr. Pharm. Res. 2020, 12, 1–6. [Google Scholar] [CrossRef]
- Dimai, H.P.; Fahrleitner-Pammer, A. Osteoporosis and Fragility Fractures: Currently available pharmacological options and future directions. Best. Pract. Res. Clin. Rheumatol. 2022, 36, 101780. [Google Scholar] [CrossRef] [PubMed]
- Sakai, E.; Farhana, F.; Yamaguchi, Y.; Tsukuba, T. Potentials of natural antioxidants from plants as antiosteoporotic agents. Stud. Nat. Prod. Chem. 2022, 72, 1–28. [Google Scholar]
- Jia, M.; Dahlman-Wright, K.; Gustafsson, J.A. Estrogen receptor alpha and beta in health and disease. Best. Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 557–568. [Google Scholar] [CrossRef]
- Erdayanti, F.Y.; Atmaka, D.R.; Kagawa, M. The Potential of Phytochemicals Lycopene in Prevention of Bone Loss due to Decreased Estrogen Hormone in Humans and Experimental Animals. Media Gizi Indones. 2022, 17, 191–203. [Google Scholar] [CrossRef]
- Lei, S.S.; Su, J.; Zhang, Y.; Huang, X.W.; Wang, X.P.; Huang, M.C.; Li, B.; Shou, D. Benefits and mechanisms of polysaccharides from Chinese medicinal herbs for anti-osteoporosis therapy: A review. Int. J. Biol. Macromol. 2021, 193, 1996–2005. [Google Scholar] [CrossRef]
- Cao, L.; Wang, J.; Zhang, Y.; Tian, F.; Wang, C. Osteoprotective effects of flavonoids: Evidence from in vivo and in vitro studies (Review). Mol. Med. Rep. 2022, 25, 200. [Google Scholar] [CrossRef]
- Yang, X.; Jiang, T.; Wang, Y.; Guo, L. The Role and Mechanism of SIRT1 in Resveratrol-regulated Osteoblast Autophagy in Osteoporosis Rats. Sci. Rep. 2019, 9, 18424. [Google Scholar] [CrossRef]
- Yu, T.; Wang, Z.; You, X.; Zhou, H.; He, W.; Li, B.; Xia, J.; Zhu, H.; Zhao, Y.; Yu, G.; et al. Resveratrol promotes osteogenesis and alleviates osteoporosis by inhibiting p53. Aging 2020, 12, 10359–10369. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Sun, Y.; Kapilevich, L.; Zhang, X.; Huang, Y. Protective effects of curcumin against osteoporosis and its molecular mechanisms: A recent review in preclinical trials. Front. Pharmacol. 2023, 14, 1249418. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Xue, Y.; Wang, T.; Xie, Q.; Lin, J.; Wang, Y. Curcumin inhibits the migration of osteoclast precursors and osteoclastogenesis by repressing CCL3 production. BMC Complement. Med. Ther. 2020, 20, 234. [Google Scholar] [CrossRef]
- Bukhari, S.N.A.; Hussain, F.; Thu, H.E.; Hussain, Z. Synergistic effects of combined therapy of curcumin and Fructus Ligustri Lucidi for treatment of osteoporosis: Cellular and molecular evidence of enhanced bone formation. J. Integr. Med. 2019, 17, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xue, J.; Shen, T.; Mu, S.; Fu, Q. Curcumin alleviates glucocorticoid-induced osteoporosis through the regulation of the Wnt signaling pathway. Int. J. Mol. Med. 2016, 37, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Lei, Y.H.; Krishnadath, D.C.; Zhu, B.Y.; Zhou, X.W. Curcumin regulates EZH2/Wnt/beta-Catenin pathway in the mandible and femur of ovariectomized osteoporosis rats. Kaohsiung J. Med. Sci. 2021, 37, 513–519. [Google Scholar] [CrossRef]
- Saud Gany, S.L.; Chin, K.-Y.; Tan, J.K.; Aminuddin, A.; Makpol, S. Curcumin as a Therapeutic Agent for Sarcopenia. Nutrients 2023, 15, 2526. [Google Scholar] [CrossRef]
- Bagherniya, M.; Mahdavi, A.; Shokri-Mashhadi, N.; Banach, M.; Von Haehling, S.; Johnston, T.P.; Sahebkar, A. The beneficial therapeutic effects of plant-derived natural products for the treatment of sarcopenia. J. Cachexia Sarcopenia Muscle 2022, 13, 2772–2790. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.Y.; Kim, C.Y. A Comprehensive Review of Pathological Mechanisms and Natural Dietary Ingredients for the Management and Prevention of Sarcopenia. Nutrients 2023, 15, 2625. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kwon, S.P.; Park, S.; Cho, S.H.; Oh, Y.; Kim, S.H.; Park, Y.H.; Jung, H.S.; Lee, D.C.; Jeong, H.; et al. Effect of Pinoresinol and Vanillic Acid Isolated from Catalpa bignonioides on Mouse Myoblast Proliferation via the Akt/mTOR Signaling Pathway. Molecules 2022, 27, 5397. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kang, M.J.; Kim, S.; Han, I.-H.; Bae, H. Screening of phytochemicals effective on relieving cancer cachexia in cisplatin-induced in vitro sarcopenia model. Mol. Cell Toxicol. 2021, 18, 111–120. [Google Scholar] [CrossRef]
- Kim, C.; Hwang, J.-K. The 5,7-Dimethoxyflavone Suppresses Sarcopenia by Regulating Protein Turnover and Mitochondria Biogenesis-Related Pathways. Nutrients 2020, 12, 1079. [Google Scholar] [CrossRef] [PubMed]
- Salucci, S.; Bartoletti-Stella, A.; Bavelloni, A.; Aramini, B.; Blalock, W.L.; Fabbri, F.; Vannini, I.; Sambri, V.; Stella, F.; Faenza, I. Extra Virgin Olive Oil (EVOO), a Mediterranean Diet Component, in the Management of Muscle Mass and Function Preservation. Nutrients 2022, 14, 3567. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.; Zhou, S.; Li, P.; Liu, C.; Yuan, B.; Zhang, S.; Liu, A. Natural products and skeletal muscle health. J. Nutr. Biochem. 2021, 93, 108619. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Yu, S.; Chen, G.; Zheng, W.; Peng, J.; Huang, X.; Chen, L. Insight into the roles of melatonin in bone tissue and bone-related diseases (Review). Int. J. Mol. Med. 2021, 47, 82. [Google Scholar] [CrossRef] [PubMed]
- Butola, K.; Bisht, V.; Kumar, R. Recent Approaches of Ocular Disease and Its Herbal Product Treatment: An Updates. J. Res. Appl. Sci. Biotechnol. 2023, 2, 102–114. [Google Scholar] [CrossRef]
- Maiuolo, J.; Bulotta, R.M.; Oppedisano, F.; Bosco, F.; Scarano, F.; Nucera, S.; Guarnieri, L.; Ruga, S.; Macri, R.; Caminiti, R.; et al. Potential Properties of Natural Nutraceuticals and Antioxidants in Age-Related Eye Disorders. Life 2022, 13, 77. [Google Scholar] [CrossRef]
- Razavi, M.S.; Ebrahimnejad, P.; Fatahi, Y.; D’Emanuele, A.; Dinarvand, R. Recent Developments of Nanostructures for the Ocular Delivery of Natural Compounds. Front. Chem. 2022, 10, 850757. [Google Scholar] [CrossRef]
- Scuderi, L.; Davinelli, S.; Iodice, C.M.; Bartollino, S.; Scapagnini, G.; Costagliola, C.; Scuderi, G. Melatonin: Implications for Ocular Disease and Therapeutic Potential. Curr. Pharm. Des. 2019, 25, 4185–4191. [Google Scholar] [CrossRef]
- Martínez-Águila, A.; Martín-Gil, A.; Carpena-Torres, C.; Pastrana, C.; Carracedo, G. Influence of Circadian Rhythm in the Eye: Significance of Melatonin in Glaucoma. Biomolecules 2021, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.A.; Mikail, M.A.; Zamakshshari, N.; Abdullah, A.H. Natural anti-aging skincare: Role and potential. Biogerontology 2020, 21, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, A.K.; Sagar, S.; Kumar, R. Past and Current Prospectives of Herbal Product for Skin Care. J. Res. Appl. Sci. Biotechnol. 2022, 1, 145–160. [Google Scholar] [CrossRef]
- Fernandes, A.; Rodrigues, P.M.; Pintado, M.; Tavaria, F.K. A systematic review of natural products for skin applications: Targeting inflammation, wound healing, and photo-aging. Phytomedicine 2023, 115, 154824. [Google Scholar] [CrossRef] [PubMed]
- Zouboulis, C.C.; Ganceviciene, R.; Liakou, A.I.; Theodoridis, A.; Elewa, R.; Makrantonaki, E. Aesthetic aspects of skin aging, prevention, and local treatment. Clin. Dermatol. 2019, 37, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Gromkowska-Kępka, K.J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Socha, K. The impact of ultraviolet radiation on skin photoaging—Review of in vitro studies. J. Cosmet. Dermatol. 2021, 20, 3427–3431. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Kim, W.J.; Park, S.Y.; Kim, H.; Chung, D.K. In vitro anti-wrinkle and skin-moisturizing effects of evening primrose (Oenothera biennis) sprout and identification of its active components. Processes 2021, 9, 145. [Google Scholar] [CrossRef]
- Younis, M.M.; Ayoub, I.M.; Mostafa, N.M.; El Hassab, M.A.; Eldehna, W.M.; Al-Rashood, S.T.; Eldahshan, O.A. GC/MS Profiling, Anti-Collagenase, Anti-Elastase, Anti-Tyrosinase and Anti-Hyaluronidase Activities of a Stenocarpus sinuatus Leaves Extract. Plants 2022, 11, 918. [Google Scholar] [CrossRef] [PubMed]
- Chaiyana, W.; Anuchapreeda, S.; Punyoyai, C.; Neimkhum, W.; Lee, K.-H.; Lin, W.-C.; Lue, S.-C.; Viernstein, H.; Mueller, M. Ocimum sanctum Linn. as a natural source of skin anti-ageing compounds. Ind. Crops Prod. 2019, 127, 217–224. [Google Scholar] [CrossRef]
- Yende, S.R.; Harle, U.N.; Chaugule, B.B. Therapeutic potential and health benefits of Sargassum species. Pharmacogn. Rev. 2014, 8, 1–7. [Google Scholar] [CrossRef]
- Jesumani, V.; Du, H.; Pei, P.; Zheng, C.; Cheong, K.L.; Huang, N. Unravelling property of polysaccharides from Sargassum sp. as an anti-wrinkle and skin whitening property. Int. J. Biol. Macromol. 2019, 140, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Egambaram, O.P.; Kesavan Pillai, S.; Ray, S.S. Materials Science Challenges in Skin UV Protection: A Review. Photochem. Photobiol. 2020, 96, 779–797. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, J.Y.; Martinez, R.M.; Morocho-Jacome, A.L.; Castillo-Gomez, T.S.; Pereda-Contreras, V.J.; Rosado, C.; Velasco, M.V.R.; Baby, A.R. Skin impacts from exposure to ultraviolet, visible, infrared, and artificial lights—A review. J. Cosmet. Laser Ther. 2021, 23, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Oh, J.; Averilla, J.N.; Kim, H.J.; Kim, J.S.; Kim, J.S. Grape peel extract and resveratrol inhibit wrinkle formation in mice model through activation of Nrf2/HO-1 signaling pathway. J. Food Sci. 2019, 84, 1600–1608. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.S.; Kim, H.J.; Na, C.; Jang, D.S.; Shin, Y.K.; Lee, S.H. The Protective Effect of Adenocaulon himalaicum Edgew. and Its Bioactive Compound Neochlorogenic Acid against UVB-Induced Skin Damage in Human Dermal Fibroblasts and Epidermal Keratinocytes. Plants 2021, 10, 1669. [Google Scholar] [CrossRef] [PubMed]
- Madan, K.; Nanda, S. In-vitro evaluation of antioxidant, anti-elastase, anti-collagenase, anti-hyaluronidase activities of safranal and determination of its sun protection factor in skin photoaging. Bioorg Chem. 2018, 77, 159–167. [Google Scholar] [CrossRef]
- Lee, K.K.; Kim, J.H.; Cho, J.J.; Choi, J.D. Inhibitory Effects of 150 Plant Extracts on Elastase Activity, and Their Anti-inflammatory Effects. Int. J. Cosmet. Sci. 2001, 21, 71–82. [Google Scholar] [CrossRef]
- Tabanelli, R.; Brogi, S.; Calderone, V. Improving Curcumin Bioavailability: Current Strategies and Future Perspectives. Pharmaceutics 2021, 13, 1715. [Google Scholar] [CrossRef]
- Zheng, Y.; Pan, C.; Zhang, Z.; Luo, W.; Liang, X.; Shi, Y.; Liang, L.; Zheng, X.; Zhang, L.; Du, Z. Antiaging effect of Curcuma longa L. essential oil on ultraviolet-irradiated skin. Microchem. J. 2020, 154, 104608. [Google Scholar] [CrossRef]
- Perva-Uzunalić, A.; Škerget, M.; Knez, Ž.; Weinreich, B.; Otto, F.; Grüner, S. Extraction of active ingredients from green tea (Camellia sinensis): Extraction efficiency of major catechins and caffeine. Food Chem. 2006, 96, 597–605. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial Properties of Green Tea Catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Li, C.; Wang, S.; Song, X. Green Tea (Camellia sinensis): A Review of Its Phytochemistry, Pharmacology, and Toxicology. Molecules 2022, 27, 3909. [Google Scholar] [CrossRef]
- Park, S.; Lee, E.-S.; Park, N.-H.; Hwang, K.; Cho, E.-G. Circadian expression of TIMP3 is disrupted by UVB irradiation and recovered by green tea extracts. Int. J. Mol. Sci. 2019, 20, 862. [Google Scholar] [CrossRef] [PubMed]
- Morgado-Carrasco, D.; Gil-Lianes, J.; Jourdain, E.; Piquero-Casals, J. [Translated article] Oral Supplementation and Systemic Drugs for Skin Aging: A Narrative Review. Actas Dermo-Sifiliogr. 2023, 114, T114–T124. [Google Scholar] [CrossRef]
- Li, M.; Guo, T.; Lin, J.; Huang, X.; Ke, Q.; Wu, Y.; Fang, C.; Hu, C. Curcumin inhibits the invasion and metastasis of triple negative breast cancer via Hedgehog/Gli1 signaling pathway. J. Ethnopharmacol. 2022, 283, 114689. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hafeez, A.A.; Khalifa, H.O.; Mahdy, E.A.M.; Sharma, V.; Hosoi, T.; Ghosh, P.; Ozawa, K.; Montano, M.M.; Fujimura, T.; Ibrahim, A.R.N.; et al. Anticancer effect of nor-wogonin (5,7,8-trihydroxyflavone) on human triple-negative breast cancer cells via downregulation of TAK1, NF-kappaB, and STAT3. Pharmacol. Rep. 2019, 71, 289–298. [Google Scholar] [CrossRef]
- Abd El-Hafeez, A.A.; Marzouk, H.M.M.; Abdelhamid, M.A.; Khalifa, H.O.; Hasanin, T.H.; Habib, A.G.; Abdelwahed, F.M.; Barakat, F.M.; Bastawy, E.M.; Abdelghani, E.M. Anti-cancer effect of Hyoscyamus muticus extract via its activation of Fas/FasL-ASK1-p38 pathway. Biotechnol. Bioprocess Eng. 2022, 27, 833–845. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020, 60, 3290–3303. [Google Scholar] [CrossRef]
- Numata, M.; Hirano, A.; Yamamoto, Y.; Yasuda, M.; Miura, N.; Sayama, K.; Shibata, M.-A.; Asai, T.; Oku, N.; Miyoshi, N. Metastasis of breast cancer promoted by circadian rhythm disruption due to light/dark shift and its prevention by dietary quercetin in mice. J. Circadian Rhythm. 2021, 19, 2. [Google Scholar] [CrossRef]
- El Khalki, L.; Maire, V.; Dubois, T.; Zyad, A. Berberine Impairs the Survival of Triple Negative Breast Cancer Cells: Cellular and Molecular Analyses. Molecules 2020, 25, 506. [Google Scholar] [CrossRef]
- Han, B.; Wang, K.; Tu, Y.; Tan, L.; He, C. Low-Dose Berberine Attenuates the Anti-Breast Cancer Activity of Chemotherapeutic Agents via Induction of Autophagy and Antioxidation. Dose Response 2020, 18, 1559325820939751. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Fokou, P.V.T.; Yamthe, L.R.T.; Tali, B.T.; Adetunji, C.O.; Rahavian, A.; Mudau, F.N.; Martorell, M.; Setzer, W.N.; Rodrigues, C.F.; et al. Phytochemicals in Prostate Cancer: From Bioactive Molecules to Upcoming Therapeutic Agents. Nutrients 2019, 11, 1483. [Google Scholar] [CrossRef] [PubMed]
- Campanelli, G.; Francois, E.; Parupathi, P.; Devarakonda, L.S.; Yang, C.; Kumar, A.; Levenson, A.S. The Therapeutic Efficacy and Mechanism of Action of Gnetin C, a Natural Compound from the Melinjo Plant, in a Preclinical Mouse Model of Advanced Prostate Cancer. Cancers 2024, 16, 1344. [Google Scholar] [CrossRef] [PubMed]
- Poustforoosh, A.; Faramarz, S.; Negahdaripour, M.; Tuzun, B.; Hashemipour, H. Investigation on the mechanisms by which the herbal remedies induce anti-prostate cancer activity: Uncovering the most practical natural compound. J. Biomol. Struct. Dyn. 2024, 42, 3349–3362. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zhang, C.; Huang, H.; Yang, B.; Xiao, G.; Kong, D.; Tian, Q.; Song, Q.; Song, Y.; Tan, H.; et al. The Natural Compound Myricetin Effectively Represses the Malignant Progression of Prostate Cancer by Inhibiting PIM1 and Disrupting the PIM1/CXCR4 Interaction. Cell Physiol. Biochem. 2018, 48, 1230–1244. [Google Scholar] [CrossRef] [PubMed]
- Bolat, Z.B.; Islek, Z.; Sahin, F.; Ucisik, M.H. Delivery of curcumin within emulsome nanoparticles enhances the anti-cancer activity in androgen-dependent prostate cancer cell. Mol. Biol. Rep. 2023, 50, 2531–2543. [Google Scholar] [CrossRef]
- Ide, H.; Lu, Y.; Noguchi, T.; Muto, S.; Okada, H.; Kawato, S.; Horie, S. Modulation of AKR1C2 by curcumin decreases testosterone production in prostate cancer. Cancer Sci. 2018, 109, 1230–1238. [Google Scholar] [CrossRef]
- Nakayama, A.; Ide, H.; Lu, Y.; Takei, A.; Fukuda, K.; Osaka, A.; Arai, G.; Horie, S.; Okada, H.; Saito, K. Effects of Curcumin Combined With the 5-alpha Reductase Inhibitor Dutasteride on LNCaP Prostate Cancer Cells. In Vivo 2021, 35, 1443–1450. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Lertpiriyapong, K.; Steelman, L.S.; Abrams, S.L.; Yang, L.V.; Murata, R.M.; Rosalen, P.L.; Scalisi, A.; Neri, L.M.; Cocco, L.; et al. Effects of resveratrol, curcumin, berberine and other nutraceuticals on aging, cancer development, cancer stem cells and microRNAs. Aging 2017, 9, 1477–1536. [Google Scholar] [CrossRef]
- Stafford, M.Y.C.; Willoughby, C.E.; Walsh, C.P.; McKenna, D.J. Prognostic value of miR-21 for prostate cancer: A systematic review and meta-analysis. Biosci. Rep. 2022, 42, BSR20211972. [Google Scholar] [CrossRef]
- Kanagasabai, T.; Li, G.; Shen, T.H.; Gladoun, N.; Castillo-Martin, M.; Celada, S.I.; Xie, Y.; Brown, L.K.; Mark, Z.A.; Ochieng, J.; et al. MicroRNA-21 deficiency suppresses prostate cancer progression through downregulation of the IRS1-SREBP-1 signaling pathway. Cancer Lett. 2022, 525, 46–54. [Google Scholar] [CrossRef]
- Li, X.; Zhang, A.; Sun, H.; Liu, Z.; Zhang, T.; Qiu, S.; Liu, L.; Wang, X. Metabolic characterization and pathway analysis of berberine protects against prostate cancer. Oncotarget 2017, 8, 65022. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.H.; Zheng, H.F.; Wang, W.L.; Wang, Y.; Zhong, L.F.; Wu, J.L.; Li, Q.X. Berberine targets epidermal growth factor receptor signaling to suppress prostate cancer proliferation in vitro. Mol. Med. Rep. 2015, 11, 2125–2128. [Google Scholar] [CrossRef]
- Liu, C.H.; Tang, W.C.; Sia, P.; Huang, C.C.; Yang, P.M.; Wu, M.H.; Lai, I.L.; Lee, K.H. Berberine inhibits the metastatic ability of prostate cancer cells by suppressing epithelial-to-mesenchymal transition (EMT)-associated genes with predictive and prognostic relevance. Int. J. Med. Sci. 2015, 12, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Wu, H.; Du, Y.; Liu, S.; Zhao, F.; Mou, X. Synthesis and biological evaluation of resveratrol amide derivatives as selective COX-2 inhibitors. Chem. Biol. Interact. 2023, 380, 110522. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, F.; Wang, F.; Li, J.; Lin, C.; Du, J. Resveratrol inhibits the proliferation of A549 cells by inhibiting the expression of COX-2. Onco Targets Ther. 2018, 11, 2981–2989. [Google Scholar] [CrossRef]
- Hsieh, T.C.; Wu, J.M. Resveratrol Suppresses Prostate Cancer Epithelial Cell Scatter/Invasion by Targeting Inhibition of Hepatocyte Growth Factor (HGF) Secretion by Prostate Stromal Cells and Upregulation of E-cadherin by Prostate Cancer Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 1760. [Google Scholar] [CrossRef]
- Aktepe, N.; Yukselten, Y. Induction of apoptosis in human hormone-refractory prostate cancer cell lines by using resveratrol in combination with AT-101. Andrologia 2022, 54, e14267. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shabestari, F.A.; Vaezi, S.; Abak, A.; Shoorei, H.; Karimi, A.; Taheri, M.; Basiri, A. Emerging impact of quercetin in the treatment of prostate cancer. Biomed. Pharmacother. 2021, 138, 111548. [Google Scholar] [CrossRef]
- Xing, N.; Chen, Y.; Mitchell, S.H.; Young, C.Y. Quercetin inhibits the expression and function of the androgen receptor in LNCaP prostate cancer cells. Carcinogenesis 2001, 22, 409–414. [Google Scholar] [CrossRef]
- Yuan, H.; Pan, Y.; Young, C.Y. Overexpression of c-Jun induced by quercetin and resverol inhibits the expression and function of the androgen receptor in human prostate cancer cells. Cancer Lett. 2004, 213, 155–163. [Google Scholar] [CrossRef]
- Van der Eecken, H.; Joniau, S.; Berghen, C.; Rans, K.; De Meerleer, G. The Use of Soy Isoflavones in the Treatment of Prostate Cancer: A Focus on the Cellular Effects. Nutrients 2023, 15, 4856. [Google Scholar] [CrossRef] [PubMed]
- Ammar, O.A.; El-Missiry, M.A.; Othman, A.I.; Amer, M.E. Melatonin is a potential oncostatic agent to inhibit HepG2 cell proliferation through multiple pathways. Heliyon 2022, 8, e08837. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Wu, Y.; Vadgama, J.V.; Wang, P. Phytochemicals in Inhibition of Prostate Cancer: Evidence from Molecular Mechanisms Studies. Biomolecules 2022, 12, 1306. [Google Scholar] [CrossRef]
- Zhang, Q.; Ruan, F.; Yang, M.; Wen, Q. Natural compound chaetocin induced DNA damage and apoptosis through reactive oxygen species-dependent pathways in A549 lung cancer cells and in vitro evaluations. IET Nanobiotechnol. 2023, 17, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Sindhoor, M.S.; Naveen, N.R.; Rao, G.K.; Gopan, G.; Chopra, H.; Park, M.N.; Alshahrani, M.M.; Jose, J.; Emran, T.B.; Kim, B. A spotlight on alkaloid nanoformulations for the treatment of lung cancer. Front. Oncol. 2022, 12, 994155. [Google Scholar] [CrossRef]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [PubMed]
- Ghareghomi, S.; Moosavi-Movahedi, F.; Saso, L.; Habibi-Rezaei, M.; Khatibi, A.; Hong, J.; Moosavi-Movahedi, A.A. Modulation of Nrf2/HO-1 by Natural Compounds in Lung Cancer. Antioxidants 2023, 12, 735. [Google Scholar] [CrossRef] [PubMed]
- Nath, R.; Das, C.; Kityania, S.; Nath, D.; Das, S.; Choudhury, M.D.; Patra, J.K.; Talukdar, A.D. Natural Flavonoids in the Prevention and Treatment of Lung Cancer: A Pharmacological Aspect. Comb. Chem. High. Throughput Screen. 2023, 26, 863–879. [Google Scholar] [CrossRef]
- Ortigosa-Palomo, A.; Quinonero, F.; Ortiz, R.; Sarabia, F.; Prados, J.; Melguizo, C. Natural Products Derived from Marine Sponges with Antitumor Potential against Lung Cancer: A Systematic Review. Mar. Drugs 2024, 22, 101. [Google Scholar] [CrossRef]
- Jiang, P.; Xu, C.; Zhang, P.; Ren, J.; Mageed, F.; Wu, X.; Chen, L.; Zeb, F.; Feng, Q.; Li, S. Epigallocatechin-3-gallate inhibits self-renewal ability of lung cancer stem-like cells through inhibition of CLOCK. Int. J. Mol. Med. 2020, 46, 2216–2224. [Google Scholar] [CrossRef] [PubMed]
- Ngai, Z.N.; Chok, K.C.; Ng, K.Y.; Koh, R.Y.; Chye, S.M. Potential role of melatonin in prevention and treatment of lung cancer. Horm. Mol. Biol. Clin. Investig. 2022, 43, 485–503. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Yang, Y.; Fan, C.; Han, J.; Wang, D.; Di, S.; Hu, W.; Liu, D.; Li, X.; Reiter, R.J.; et al. Melatonin as a potential anticarcinogen for non-small-cell lung cancer. Oncotarget 2016, 7, 46768–46784. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Chen, P.C.; Chiou, P.C.; Hsu, C.J.; Liu, P.I.; Yang, Y.C.; Reiter, R.J.; Yang, S.F.; Tang, C.H. Melatonin suppresses lung cancer metastasis by inhibition of epithelial-mesenchymal transition through targeting to Twist. Clin. Sci. 2019, 133, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, X.; Chen, T.; Cheng, X.; Xiao, H.; Meng, X.; Jiang, Y. Inhibition and potential treatment of colorectal cancer by natural compounds via various signaling pathways. Front. Oncol. 2022, 12, 956793. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Xie, H.; Shen, W.; Shao, L.; Zeng, L.; Huang, X.; Zhu, Q.; Zhai, X.; Li, K.; Qiu, Z.; et al. The Synergism of Natural Compounds and Conventional Therapeutics against Colorectal Cancer Progression and Metastasis. Front. Biosci. 2022, 27, 263. [Google Scholar] [CrossRef] [PubMed]
- Rejhova, A.; Opattova, A.; Cumova, A.; Sliva, D.; Vodicka, P. Natural compounds and combination therapy in colorectal cancer treatment. Eur. J. Med. Chem. 2018, 144, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Shen, Y. Curcumin affects apoptosis of colorectal cancer cells through ATF6-mediated endoplasmic reticulum stress. Chem. Biol. Drug Des. 2024, 103, e14433. [Google Scholar] [CrossRef]
- Ismail, N.I.; Othman, I.; Abas, F.; Lajis, N.H.; Naidu, R. Mechanism of Apoptosis Induced by Curcumin in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 2454. [Google Scholar] [CrossRef]
- Chen, M.; Tan, A.H.; Li, J. Curcumin Represses Colorectal Cancer Cell Proliferation by Triggering Ferroptosis via PI3K/Akt/mTOR Signaling. Nutr. Cancer 2023, 75, 726–733. [Google Scholar] [CrossRef]
- Amin, M.; Razi, M.; Sarrafzadeh-Rezaei, F.; Shalizar Jalali, A.; Najafi, G. Berberine inhibits experimental varicocele-induced cell cycle arrest via regulating cyclin D1, cdk4 and p21 proteins expression in rat testicles. Andrologia 2018, 50, e12984. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, X.; Tan, H.Y.; Li, S.; Tsang, C.M.; Tsao, S.W.; Feng, Y. Berberine Suppresses Cyclin D1 Expression through Proteasomal Degradation in Human Hepatoma Cells. Int. J. Mol. Sci. 2016, 17, 1899. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Shi, C.J.; Fu, W.M.; Zhang, J.F. Berberine inhibits tumour growth in vivo and in vitro through suppressing the lincROR-Wnt/beta-catenin regulatory axis in colorectal cancer. J. Pharm. Pharmacol. 2023, 75, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Zhan, Y.Y.; Hou, J.; Xu, B.; Chen, B.; Tian, Y.; Wu, D.; Zhao, Y.; Zhang, Y.; Chen, X.; et al. Berberine binds RXRalpha to suppress beta-catenin signaling in colon cancer cells. Oncogene 2017, 36, 6906–6918. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.X.; Xiong, R.G.; Huang, S.Y.; Zhou, D.D.; Saimaiti, A.; Zhao, C.N.; Shang, A.; Zhang, Y.J.; Gan, R.Y.; Li, H.B. Effects and mechanisms of resveratrol for prevention and management of cancers: An updated review. Crit. Rev. Food Sci. Nutr. 2023, 63, 12422–12440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Y.; Sun, C.; Chen, X.; Han, L.; Wang, T.; Liu, J.; Chen, X.; Zhao, D. Effect of Pterostilbene, a Natural Derivative of Resveratrol, in the Treatment of Colorectal Cancer through Top1/Tdp1-Mediated DNA Repair Pathway. Cancers 2021, 13, 4002. [Google Scholar] [CrossRef] [PubMed]
- Beshay, O.N.; Ewees, M.G.; Abdel-Bakky, M.S.; Hafez, S.; Abdelrehim, A.B.; Bayoumi, A.M.A. Resveratrol reduces gentamicin-induced EMT in the kidney via inhibition of reactive oxygen species and involving TGF-beta/Smad pathway. Life Sci. 2020, 258, 118178. [Google Scholar] [CrossRef]
- Li, R.; Liang, M.; Liang, X.; Yang, L.; Su, M.; Lai, K.P. Chemotherapeutic Effectiveness of Combining Cetuximab for Metastatic Colorectal Cancer Treatment: A System Review and Meta-Analysis. Front. Oncol. 2020, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Yang, S. Epigallocatechin-3-gallate inhibits proliferation and triggers apoptosis in colon cancer via the hedgehog/phosphoinositide 3-kinase pathways. Can. J. Physiol. Pharmacol. 2021, 99, 910–920. [Google Scholar] [CrossRef]
- Perez-Valero, A.; Magadan-Corpas, P.; Ye, S.; Serna-Diestro, J.; Sordon, S.; Huszcza, E.; Poplonski, J.; Villar, C.J.; Lombo, F. Antitumor Effect and Gut Microbiota Modulation by Quercetin, Luteolin, and Xanthohumol in a Rat Model for Colorectal Cancer Prevention. Nutrients 2024, 16, 1161. [Google Scholar] [CrossRef]
- Liu, Y.; Lang, T.; Jin, B.; Chen, F.; Zhang, Y.; Beuerman, R.W.; Zhou, L.; Zhang, Z. Luteolin inhibits colorectal cancer cell epithelial-to-mesenchymal transition by suppressing CREB1 expression revealed by comparative proteomics study. J. Proteom. 2017, 161, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, B.; Gao, F.; Shi, R. Modulation of G2/M cell cycle arrest and apoptosis by luteolin in human colon cancer cells and xenografts. Oncol. Lett. 2018, 15, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, H.; Kim, H. Anti-Inflammatory Effect of Protopine through MAPK and NF-κB Signaling Regulation in HepG2 Cell. Molecules 2022, 27, 4601. [Google Scholar] [CrossRef] [PubMed]
- Mohebali, N.; Pandurangan, A.K.; Mustafa, M.R.; Anandasadagopan, S.K.; Alagumuthu, T. Vernodalin induces apoptosis through the activation of ROS/JNK pathway in human colon cancer cells. J. Biochem. Mol. Toxicol. 2020, 34, e22587. [Google Scholar] [CrossRef] [PubMed]
- Sampath, G.; Shyu, D.J.H.; Rameshkumar, N.; Krishnan, M.; Sivasankar, P.; Kayalvizhi, N. Synthesis and Characterization of Pyrogallol Capped Silver Nanoparticles and Evaluation of Their In Vitro Anti-Bacterial, Anti-cancer Profile Against AGS Cells. J. Clust. Sci. 2021, 32, 549–557. [Google Scholar] [CrossRef]
- Zhong, Y.; Tian, X.; Jiang, X.; Dang, W.; Cheng, M.; Li, N.; Liu, Y. Novel Ziyuglycoside II derivatives inhibit MCF-7 cell proliferation via inducing apoptosis and autophagy. Bioorg. Chem. 2023, 139, 106752. [Google Scholar] [CrossRef] [PubMed]
- Radajewska, A.; Moreira, H.; Bęben, D.; Siwiela, O.; Szyjka, A.; Gębczak, K.; Nowak, P.; Frąszczak, J.; Emhemmed, F.; Muller, C.D.; et al. Combination of Irinotecan and Melatonin with the Natural Compounds Wogonin and Celastrol for Colon Cancer Treatment. Int. J. Mol. Sci. 2023, 24, 9544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, C.; Goel, A. A combined treatment with melatonin and andrographis promotes autophagy and anticancer activity in colorectal cancer. Carcinogenesis 2022, 43, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, K.; Mohamed, H.; Shah, A.M.; Yu, S.; Akhlaq, M.; Xiao, H.; Li, S.; Naz, T.; Nosheen, S.; Bai, X.; et al. In Vitro Anticancer Potential of Berberis lycium Royle Extracts against Human Hepatocarcinoma (HepG2) Cells. BioMed Res. Int. 2020, 2020, 8256809. [Google Scholar] [CrossRef]
- Zhu, Y.; Xie, N.; Chai, Y.; Nie, Y.; Liu, K.; Liu, Y.; Yang, Y.; Su, J.; Zhang, C. Apoptosis Induction, a Sharp Edge of Berberine to Exert Anti-Cancer Effects, Focus on Breast, Lung, and Liver Cancer. Front. Pharmacol. 2022, 13, 803717. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, K.; Gu, C.; Yu, G.; Zhao, D.; Mai, W.; Zhong, Y.; Liu, S.; Nie, Y.; Yang, H. Berberine, a natural plant alkaloid, synergistically sensitizes human liver cancer cells to sorafenib. Oncol. Rep. 2018, 40, 1525–1532. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Li, M.; Cao, Y.; Li, C.; Zhou, W.; Ji, G.; Zhang, L. Berberine Alleviates Non-alcoholic Steatohepatitis Through Modulating Gut Microbiota Mediated Intestinal FXR Activation. Front. Pharmacol. 2021, 12, 750826. [Google Scholar] [CrossRef]
- Diab, R.F.; Abdelghany, T.M.; Gad, S.; Elbakry, A.M. Novel resveratrol smart lipids; design, formulation, and biological evaluation of anticancer activity. J. Pharm. Pharmacol. 2024, 76, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yang, M.; Qu, Z.; Zhou, J.; Zhang, Q. Resveratrol enhances anticancer effects of paclitaxel in HepG2 human liver cancer cells. BMC Complement. Altern. Med. 2017, 17, 477. [Google Scholar] [CrossRef]
- Guo, A.; Chang, Y.; Lin, J.; Guo, J.; He, Y.; Wang, C.; Wu, Z.; Xing, Y.; Jin, F.; Deng, Y. Resveratrol Enhances Anticancer Effects of Silybin on HepG2 Cells and H22 Tumor-bearing Mice via Inducing G2/M Phase Arrest and Increasing Bax/Bcl-2 Ratio. Comb. Chem. High. Throughput Screen. 2024. [Google Scholar] [CrossRef]
- Amiri, F.; Zarnani, A.H.; Zand, H.; Koohdani, F.; Jeddi-Tehrani, M.; Vafa, M. Synergistic anti-proliferative effect of resveratrol and etoposide on human hepatocellular and colon cancer cell lines. Eur. J. Pharmacol. 2013, 718, 34–40. [Google Scholar] [CrossRef]
- Sethi, G.; Rath, P.; Chauhan, A.; Ranjan, A.; Choudhary, R.; Ramniwas, S.; Sak, K.; Aggarwal, D.; Rani, I.; Tuli, H.S. Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements. Pharmaceutics 2023, 15, 712. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.A.S.; Kalinina, E.V.; Tatarskiy, V.V.; Volodina, Y.L.; Petrova, A.C.; Novichkova, M.D.; Zhdanov, D.D.; Shtil, A.A. Suppression of the Antioxidant System and PI3K/Akt/mTOR Signaling Pathway in Cisplatin-Resistant Cancer Cells by Quercetin. Bull. Exp. Biol. Med. 2022, 173, 760–764. [Google Scholar] [CrossRef]
- Seo, H.S.; Ku, J.M.; Choi, H.S.; Choi, Y.K.; Woo, J.K.; Kim, M.; Kim, I.; Na, C.H.; Hur, H.; Jang, B.H.; et al. Quercetin induces caspase-dependent extrinsic apoptosis through inhibition of signal transducer and activator of transcription 3 signaling in HER2-overexpressing BT-474 breast cancer cells. Oncol. Rep. 2016, 36, 31–42. [Google Scholar] [CrossRef]
- Huang, J.; Xu, H. Matrine: Bioactivities and Structural Modifications. Curr. Top. Med. Chem. 2016, 16, 3365–3378. [Google Scholar] [CrossRef]
- Qiu, G.; Xie, J.; Li, F.; Han, K.; Long, Q.; Kowah, J.A.H.; Gao, R.; Wang, L.; Liu, X. Design, synthesis and biological evaluation of matrine contains benzimidazole derivatives as dual TOPOI and PARP inhibitors for cancer therapy. Eur. J. Med. Chem. 2024, 270, 116348. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, J.; Guo, Z.; Hua, H.; Zhang, H.; Liu, Y.; Jiang, Y. Targeting HSP70 chaperones by rhein sensitizes liver cancer to artemisinin derivatives. Phytomedicine 2024, 122, 155156. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.X.; Xia, W.; Yue, W.; Peng, C.; Rahman, K.; Zhang, H. Rhein: A Review of Pharmacological Activities. Evid. Based Complement. Alternat Med. 2015, 2015, 578107. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J.; Xia, L. Plant-derived natural products and combination therapy in liver cancer. Front. Oncol. 2023, 13, 1116532. [Google Scholar] [CrossRef]
- Mawatari, K.; Koike, N.; Nohara, K.; Wirianto, M.; Uebanso, T.; Shimohata, T.; Shikishima, Y.; Miura, H.; Nii, Y.; Burish, M.J.; et al. The Polymethoxyflavone Sudachitin Modulates the Circadian Clock and Improves Liver Physiology. Mol. Nutr. Food Res. 2023, 67, e2200270. [Google Scholar] [CrossRef]
- Fernandez-Palanca, P.; Mendez-Blanco, C.; Fondevila, F.; Tunon, M.J.; Reiter, R.J.; Mauriz, J.L.; Gonzalez-Gallego, J. Melatonin as an Antitumor Agent against Liver Cancer: An Updated Systematic Review. Antioxidants 2021, 10, 103. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, R.; Wang, Z.; Chen, Z.; Wang, G.; Guan, S.; Lu, J. Melatonin Prevents against Ethanol-Induced Liver Injury by Mitigating Ferroptosis via Targeting Brain and Muscle ARNT-like 1 in Mice Liver and HepG2 Cells. J. Agric. Food Chem. 2022, 70, 12953–12967. [Google Scholar] [CrossRef]
- Crespo, M.; Leiva, M.; Sabio, G. Circadian Clock and Liver Cancer. Cancers 2021, 13, 3631. [Google Scholar] [CrossRef] [PubMed]
- Manieri, E.; Folgueira, C.; Rodriguez, M.E.; Leiva-Vega, L.; Esteban-Lafuente, L.; Chen, C.; Cubero, F.J.; Barrett, T.; Cavanagh-Kyros, J.; Seruggia, D.; et al. JNK-mediated disruption of bile acid homeostasis promotes intrahepatic cholangiocarcinoma. Proc. Natl. Acad. Sci. USA 2020, 117, 16492–16499. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Xu, S.; Fayed, M. Natural Bioactive Compounds Promote Cell Apoptosis in Gastric Cancer Treatment: Evidence from Network Pharmacological Study and Experimental Analysis. J. Chem. 2023, 2023, 6316589. [Google Scholar] [CrossRef]
- Peng, X.; Shi, J.; Zhao, Z.; Tong, R.; Zhang, X.; Zhong, L. Emetine, a small molecule natural product, displays potent anti-gastric cancer activity via regulation of multiple signaling pathways. Cancer Chemother. Pharmacol. 2023, 91, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Li, Z.; Li, Y.; Deng, X.; Chen, Y.; Ma, X.; Zeng, J.; Zhao, Y. Natural products targeting signaling pathways associated with regulated cell death in gastric cancer: Recent advances and perspectives. Phytother. Res. 2023, 37, 2661–2692. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Wei, F.; Sun, G.; Wen, Y.; Xiang, J.; Su, F.; Zhan, L.; Nian, Q.; Chen, Y.; Zeng, J. Natural compounds targeting glycolysis as promising therapeutics for gastric cancer: A review. Front. Pharmacol. 2022, 13, 1004383. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Q.; Hu, X.; Zhang, S.; Jiang, Y.; Yao, G.; Hu, K.; Xu, X.; Liang, B.; Wu, Q.; et al. Disrupting Circadian Rhythm via the PER1-HK2 Axis Reverses Trastuzumab Resistance in Gastric Cancer. Cancer Res. 2022, 82, 1503–1517. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Huang, Q.; Zhu, H. Melatonin Inhibits the Proliferation of Gastric Cancer Cells Through Regulating the miR-16-5p-Smad3 Pathway. DNA Cell Biol. 2018, 37, 244–252. [Google Scholar] [CrossRef]
- Hofmeister, E.N.; Fisher, S.; Palesh, O.; Innominato, P.F. Does circadian rhythm influence gastrointestinal toxicity? Curr. Opin. Support. Palliat. Care 2020, 14, 120–126. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhao, Y.; Chen, Q.; Zhu, S.; Niu, Y.; Ye, Z.; Hu, P.; Chen, D.; Xu, P.; Chen, J. Hypoxic exosomal HIF-1α-stabilizing circZNF91 promotes chemoresistance of normoxic pancreatic cancer cells via enhancing glycolysis. Oncogene 2021, 40, 5505–5517. [Google Scholar] [CrossRef]
- Guo, W.; Ding, Y.; Pu, C.; Wang, Z.; Deng, W.; Jin, X. Curcumin inhibits pancreatic cancer cell proliferation by regulating Beclin1 expression and inhibiting the hypoxia-inducible factor-1alpha-mediated glycolytic pathway. J. Gastrointest. Oncol. 2022, 13, 3254–3262. [Google Scholar] [CrossRef]
- He, X.Y.; Wu, L.J.; Wang, W.X.; Xie, P.J.; Chen, Y.H.; Wang, F. Amygdalin—A pharmacological and toxicological review. J. Ethnopharmacol. 2020, 254, 112717. [Google Scholar] [CrossRef]
- Shi, J.; Chen, Q.; Xu, M.; Xia, Q.; Zheng, T.; Teng, J.; Li, M.; Fan, L. Recent updates and future perspectives about amygdalin as a potential anticancer agent: A review. Cancer Med. 2019, 8, 3004–3011. [Google Scholar] [CrossRef]
- Aamazadeh, F.; Ostadrahimi, A.; Rahbar Saadat, Y.; Barar, J. Bitter apricot ethanolic extract induces apoptosis through increasing expression of Bax/Bcl-2 ratio and caspase-3 in PANC-1 pancreatic cancer cells. Mol. Biol. Rep. 2020, 47, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Z. The Holistic Approach to Cancer Pain Management. Ulster Med. J. 2022, 91, 45–49. [Google Scholar]
- Chen, T.; Xiao, Z.; Liu, X.; Wang, T.; Wang, Y.; Ye, F.; Su, J.; Yao, X.; Xiong, L.; Yang, D.-H. Natural products for combating multidrug resistance in cancer. Pharmacol. Res. 2024, 202, 107099. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Lu, J.-J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect. 2021, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.; Brazeau, D.; Amin, A.R. Perspectives on natural compounds in chemoprevention and treatment of cancer: An update with new promising compounds. Eur. J. Cancer 2021, 149, 165–183. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, S. Dietary phytochemicals and their role in cancer chemoprevention. J. Cancer Metastasis Treat. 2021, 7, 51. [Google Scholar] [CrossRef]
- Barba-Ostria, C.; Carrera-Pacheco, S.E.; Gonzalez-Pastor, R.; Heredia-Moya, J.; Mayorga-Ramos, A.; Rodriguez-Polit, C.; Zuniga-Miranda, J.; Arias-Almeida, B.; Guaman, L.P. Evaluation of Biological Activity of Natural Compounds: Current Trends and Methods. Molecules 2022, 27, 4490. [Google Scholar] [CrossRef]
- Khadka, B.; Lee, J.-Y.; Park, D.H.; Kim, K.-T.; Bae, J.-S. The Role of Natural Compounds and their Nanocarriers in the Treatment of CNS Inflammation. Biomolecules 2020, 10, 1401. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef]
- Zakharenkova, S.A.; Abzianidze, V.V.; Moiseeva, N.I.; Lukina, D.S.; Chistyi, L.S.; Krivorotov, D.V.; Trishin, Y.G. Antitumor activity of phaeosphaeride A modified with nitrogen heterocyclic groups. Mendeleev Commun. 2021, 31, 662–663. [Google Scholar] [CrossRef]
- Huang, L.; Huang, X.H.; Yang, X.; Hu, J.Q.; Zhu, Y.Z.; Yan, P.Y.; Xie, Y. Novel nano-drug delivery system for natural products and their application. Pharmacol. Res. 2024, 201, 107100. [Google Scholar] [CrossRef]
- Pavithra, R.; Khan, M.R.; Khan, M.S. Recent advancements in natural compounds for cancer therapy and prevention. Phytochem. Rev. 2024. [Google Scholar] [CrossRef]
- Abd-Alrahman, A.M.; Ramadan, M.M.; Maraay, M.F.; Salem, R.; Saleh, F.M.; Hashim, M.A.; Zhernyakova, A.; El-Messery, T.M. Production of natural flavor compounds using Bacillus subtilis-fermented soybean meal extract and their biological potential: A comprehensive in vitro study. Front. Nutr. 2024, 10, 1280209. [Google Scholar] [CrossRef]
- Ueberschlag, S.L.; Seay, J.R.; Roberts, A.H.; DeSpirito, P.C.; Stith, J.M.; Folz, R.J.; Carter, K.A.; Weiss, E.P.; Zavorsky, G.S. The Effect of Protandim(R) Supplementation on Athletic Performance and Oxidative Blood Markers in Runners. PLoS ONE 2016, 11, e0160559. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Hu, Q.; Shen, S. Enhanced antitumor efficacy and attenuated cardiotoxicity of doxorubicin in combination with lycopene liposomes. J. Liposome Res. 2019, 30, 37–44. [Google Scholar] [CrossRef]
- Amiama-Roig, A.; Verdugo-Sivianes, E.M.; Carnero, A.; Blanco, J.R. Chronotherapy: Circadian Rhythms and Their Influence in Cancer Therapy. Cancers 2022, 14, 5071. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Monika, M.; Singh, M. Circadian rhythms regulated pulsatile drug delivery system. Int. J. Health Sci. 2022, 6, 7944–7965. [Google Scholar] [CrossRef]
- Fishbein, A.B.; Knutson, K.L.; Zee, P.C. Circadian disruption and human health. J. Clin. Investig. 2021, 131, e148286. [Google Scholar] [CrossRef]
- Zhu, X.; Maier, G.; Panda, S. Learning from circadian rhythm to transform cancer prevention, prognosis, and survivorship care. Trends Cancer 2024, 10, 196–207. [Google Scholar] [CrossRef]
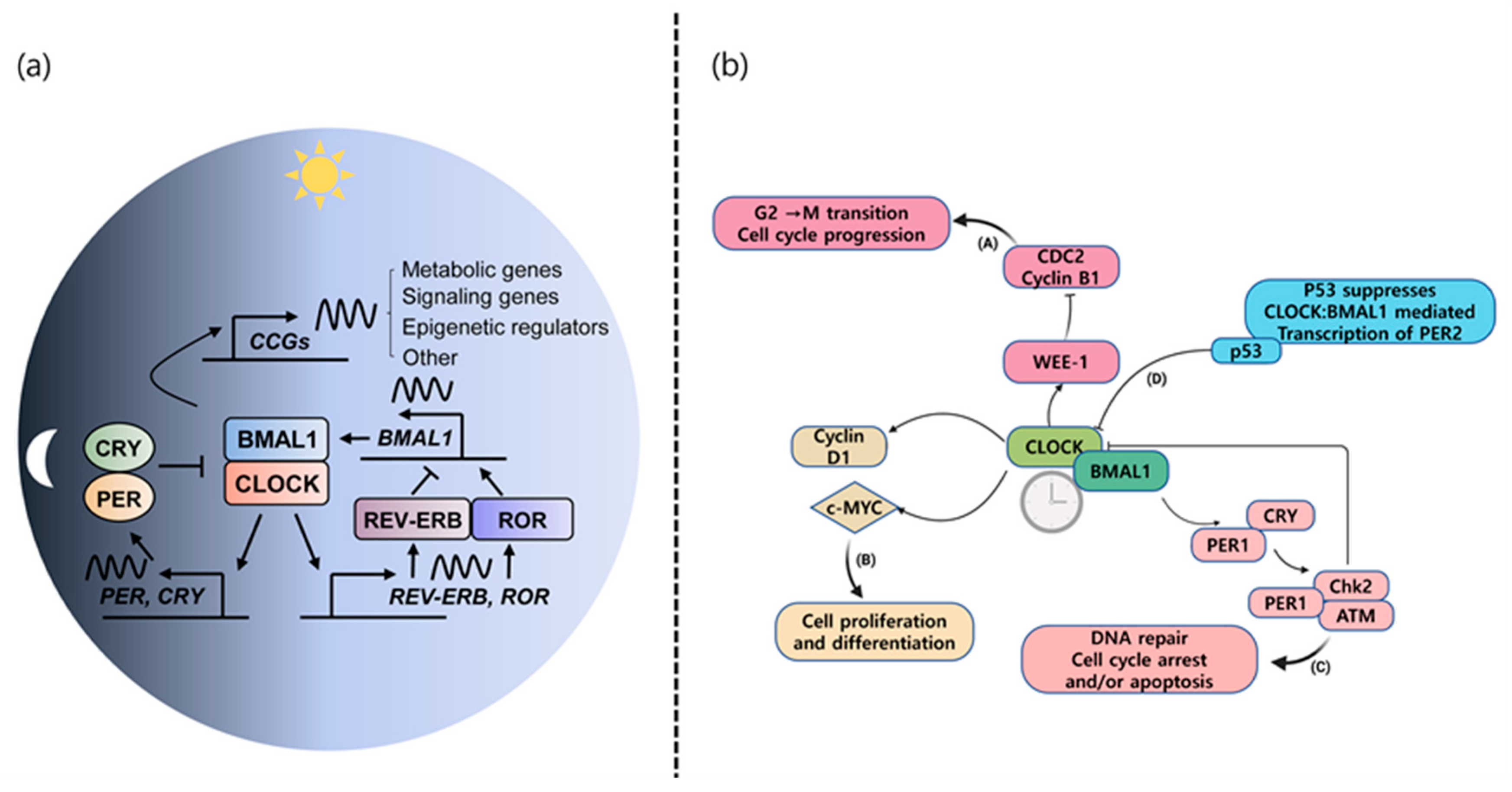
| Group | Description |
|---|---|
| Alkaloids | Nitrogen-bearing molecules found in a variety of plants. Used in drugs like vincristine (from Madagascar periwinkle) and sinomenine (from Sinomenium acutum) [325] for cancer treatment; atropine (from deadly nightshade) for emergency medicine, anesthesia, cardiology, and other specialties as well as reversing bradycardia and managing organophosphate poisoning [326]; and morphine (from the poppy plant, Papaver somniferum) for pain relief in cancer patients [327]. |
| Bitters | Plants with a bitter taste that stimulate salivary glands and digestive organs. Examples include hop acids from the hop plant (Humulus lupulus) exhibiting anticancer activity [328], and amarogentin (Gentiana lutea radix, L.), a bitter taste receptor activator regulating a variety of cell signaling including AMP-activated protein kinase (AMPK), STAT3, Akt, ERK, and p53 [329] |
| Cardiac glycosides | Compounds (digitoxin, digoxin, and convallotoxin) in medicinal plants like Foxglove and Lily of the Valley that support heart strength and stimulate urine production. |
| Cyanogenic glycosides | Glycosides based on toxic cyanide found in plants like wild cherry and elderberry. Used in small doses as a muscle relaxant and to soothe dry coughs. |
| Flavonoids | Compounds found in many plants that often act as pigments and have antiviral and anti-inflammatory properties. They generally have a 15-carbon skeleton and are known to strengthen capillaries and prevent leakage into tissues. Examples include lemon and buckwheat. |
| Minerals | Many plants draw minerals from the soil and convert them into a form easily used by the human body. Example: horsetail, which is high in silica and used for arthritis because it supports the repair of connective tissue. |
| Phenols | Plant compounds thought to protect against infection and herbivory. Often anti-inflammatory and antiseptic. Phenols vary in structure and range from salicylic acid to complex sugar-containing phenolic acids. Examples include wintergreen, willow, and mint family. |
| Polysaccharides | Sugar molecules found in all plants that can form jelly-like masses used to treat dry or irritated tissues. |
| Proanthocyanins | Pigments related to tannins and flavonoids that protect circulation. Found in red grapes, blackberries, and hawthorn berries. |
| Saponins | Active compounds that produce lather in water. Found in plants like agave and wild yam. Steroidal saponins are very similar to the chemical structures of many of the human body’s hormones including estrogen and cortisol. Some of them are used to produce synthetic hormones. |
| Tannins | Compounds produced by most plants that deter herbivory and are useful in curing leather. Found in plants like oak bark and black catechu. |
| Vitamins | Many plants contain high levels of useful vitamins. Examples include watercress, rose hips, and sea buckthorn, which have high levels of vitamins B, C, and E. |
| Terpenes/terpenoids | Terpenes and terpenoids are the main bioactive compounds of essential oils, volatile and concentrated liquids extracted from different parts of plants. Example: tea tree oil, a strong antiseptic. Carvacrol, carvone, eugenol, geraniol, and thymol for antimicrobial activity [330]. Paclitaxel (taxol from Taxus brevifolia tree) for cancer treatment [331] |
| Fatty acids | Omega-3 fatty acids play an important role in regulating brain function and lowering cardiovascular disease risk; alpha-linolenic acid from nuts and seeds, flaxseed oil, soybean oil, and canola oil [332]; eicosapentaenoic and docosahexaenoic acid from microalgae [333] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ki, M.-R.; Youn, S.; Kim, D.H.; Pack, S.P. Natural Compounds for Preventing Age-Related Diseases and Cancers. Int. J. Mol. Sci. 2024, 25, 7530. https://doi.org/10.3390/ijms25147530
Ki M-R, Youn S, Kim DH, Pack SP. Natural Compounds for Preventing Age-Related Diseases and Cancers. International Journal of Molecular Sciences. 2024; 25(14):7530. https://doi.org/10.3390/ijms25147530
Chicago/Turabian StyleKi, Mi-Ran, Sol Youn, Dong Hyun Kim, and Seung Pil Pack. 2024. "Natural Compounds for Preventing Age-Related Diseases and Cancers" International Journal of Molecular Sciences 25, no. 14: 7530. https://doi.org/10.3390/ijms25147530
APA StyleKi, M.-R., Youn, S., Kim, D. H., & Pack, S. P. (2024). Natural Compounds for Preventing Age-Related Diseases and Cancers. International Journal of Molecular Sciences, 25(14), 7530. https://doi.org/10.3390/ijms25147530