
Structure and Functions of HMGB3 Protein


Abstract
:1. Introduction
2. Structure of HMGB3 Protein
3. Post-Translational Modifications of Human HMGB3
3.1. Acetylation
3.2. Methylation
3.3. Phosphorylation
3.4. Glycosylation
3.5. Ubiquitination
3.6. Redox State of HMGB3
4. Molecular Partners of HMGB3
4.1. DNA
4.2. RNA
4.3. p53
4.4. HOX Proteins
4.5. PARP1
4.6. SOX9
4.7. HMGA1
4.8. Receptors
4.8.1. TLRs
4.8.2. TREM1
4.9. Putative HMGB3 Partners

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Partners | Functions | References |
|---|---|---|
| DNA | HMGB1–3 proteins (DNA chaperones) bend DNA and facilitate its binding to transcription factors and various proteins. | [9,11,12,35,36,73,108] |
| In addition, they recognize and preferentially bind to DNA regions with various structural abnormalities. | [9,11,56,118] | |
| Overexpression of HMGB3 can inhibit the DNA bending process and interfere with the binding of DNA and transcription factors. | [39] | |
| RNA | HMGB3 binds RNA, facilitating their binding to pattern recognition receptors, such as TLRs, and control tat mRNA processing and translation. HMGB3 has been found in cytoplasmic ribonucleoprotein (RNP) complexes, which are formed in response to cellular stress. | [14,34,126,127] |
| p53 | The interaction between p53 and DNA requires HMGB proteins. | [9,11,129,130] |
| The impaired binding of HMGB1–3 to DNA interferes with the interaction between p53 and DNA, which leads to disruption of the antitumor activity of p53. | [39] | |
| TLR3, TLR7, TLR9 | In vertebrates, the interaction between HMGB3 and TLR3, TLR7, and TLR9, which occurs on the membrane, leads to upregulation of the production of ROS and activation of NF-kB signaling, which ultimately regulates tumor growth and metastasis. | [34,155,163,164] |
| Activation of TLR2/4 by HMGB1–3 proteins stimulates the release of cytokines and inflammatory factors. | [165] | |
| HOX | Increased expression of HOXB6, HOXB7, and HOXB8 in hematopoietic cell lines blocks myeloid differentiation. The co-expression of HMGB3 and HOXB6 also suggests a joint role of these proteins in erythropoiesis. | [131,132,133,134,135,168] |
| PARP1 | Direct interaction between HMGB3 and the PARP1 protein, which plays a decisive role in DNA repair processes, was revealed. An increase in the amount of HMGB3 leads to increasing enzymatic activity on the part of PARP1. | [139,140,141] |
| HMGB3 promotes ovarian cancer resistance to PARP inhibitors through direct interaction with PARP1. | [137] | |
| SOX9 | A direct physical interaction between HMGB3 protein and SOX9, which promotes the binding of SOX9 to NANOG, has been demonstrated. When HMGB3 binds to SOX9, NANOG transactivation is induced, which can lead to increased cell survival and migration of prostate adenocarcinoma cells. | [143] |
| PAX | PAX5 and PAX6 are tissue-specific transcription factors. PAX5 is associated with neuronal development, spermatogenesis, and B-cell differentiation. In esophageal cancer cell lines, PAX5 knockdown promotes resistance of tumor cells to cisplatin, inhibits proliferation, promotes apoptosis, and induces activation of p53 signaling. Functions of the PAX complex with HMGB3 are unknown. | [167,169,170] |
| HMGA1 | As shown by 2-DE and LC-MS/MS, HMGA is a molecular partner of HMGB3. HMGB1, HMGB2, HMGB3, and HMGA1 are involved in RNA processing. Additionally, their action presumably affects chromatin structure and DNA-dependent processes. | [149,151] |
| HMGB1 | Databases show a direct physical interaction between HMGB1 and HMGB3. This interaction has unknown functional significance. | [151] |
| TERF2 | TRF2 is involved in the processes of DNA replication, RNA-dependent DNA replication, DNA packaging, DNA metabolic processes, and nucleosome assembly, among others. Databases show a direct physical interaction between TERF and HMGB3, although the functions are unknown. | [171] |
| NUP98 | NUP98–HMGB3 acts as an oncogene responsible for rapid and transplantable MPD-like leukemia in recipient mice, which is associated with defects in myelomonocytic cell differentiation. | [172] |
| TREM1 | Suppression of HMGB3 expression inhibits cell viability, promotes cell apoptosis and cell cycle arrest, and suppresses cell migration and invasion in thyroid cancer. Cytoplasmic HMGB3 activates nucleic acid-mediated TLR3/NF-κB signaling, and extracellular HMGB3 interacts with the transmembrane receptor TREM1 in PTC. | [155] |
| HIF1α | HMGB3 prevents mammosphere formation in breast cancer through binding to HIF1α. | [173,174,175] |
| HMGB3 may promote ROS production and tumor cell proliferation by inducing HIF-1α expression. The silencing of HMGB3 attenuates HIF1α expression, leading to suppression of tumor cell proliferation. | [39] | |
| SDCBP | The IntAct database shows a direct physical interaction between SDCBP and HMGB3 (Figure 8), although the functions are unknown. | [128] |
| MEOX | The databases show a direct physical interaction between MEOX and HMGB3 (Figure 8), with the functions unknown. | [128] |
| hTERT | The HMGB3/hTERT axis is important for resistance to radiation therapy in cervical cancer. | [123] |
| MAPK | HMGB3 promotes the development of malignant phenotypes and stemness of ovarian cancer through the MAPK/ERK signaling pathway. | [142] |
| Bcl-2/Bax | In gastric adenocarcinoma, downregulation of HMGB3 expression can dramatically suppress cancer cell proliferation, mainly through inducing G0/G1 blockade in tumor cells, regulating p53 and p21 signaling pathways, and reducing Bcl-2/Bax (anti-apoptotic factor/pro-apoptotic factor) levels. | [176] |
| MMP2, MMP9 | Reducing the expression of HMGB3 leads to inhibition of invasion and migration of gastric cancer cells by suppressing the activation of MMP2 and MMP9. | [177] |
| ARHGAP11A, H1.4, LRIG1, PRR11 and COL4A1 | A prognostic model consisting of six gene products (HMGB3, ARHGAP11A, H1.4, LRIG1, PRR11, and COL4A1) is a reliable predictor of esophageal cancer. | [150] |
| ER, PR | According to bioinformatics analysis, the co-expression of HMGB3 with ER and PR is associated with poor prognosis in breast cancer. | [107] |
| HMGN1, HMGN2, HMGN3, HMGN4 | High rates of expression of HMGB3, HMGN1, HMGN2, HMGN3, and HMGN4 are shown in gastric cancer. | [151] |
| Knockdown of HMGB3 suppresses the proliferation of cancer cells, stops their migration, and affects sensitivity to gastric cancer chemotherapy. | [39,177] | |
| Heparin | The IntAct databases show a direct physical interaction between heparin and HMGB3 (Figure 8), with the functions unknown. | [128] |
5. Expression of HMGB3
5.1. Regulation of HMGB3 Expression
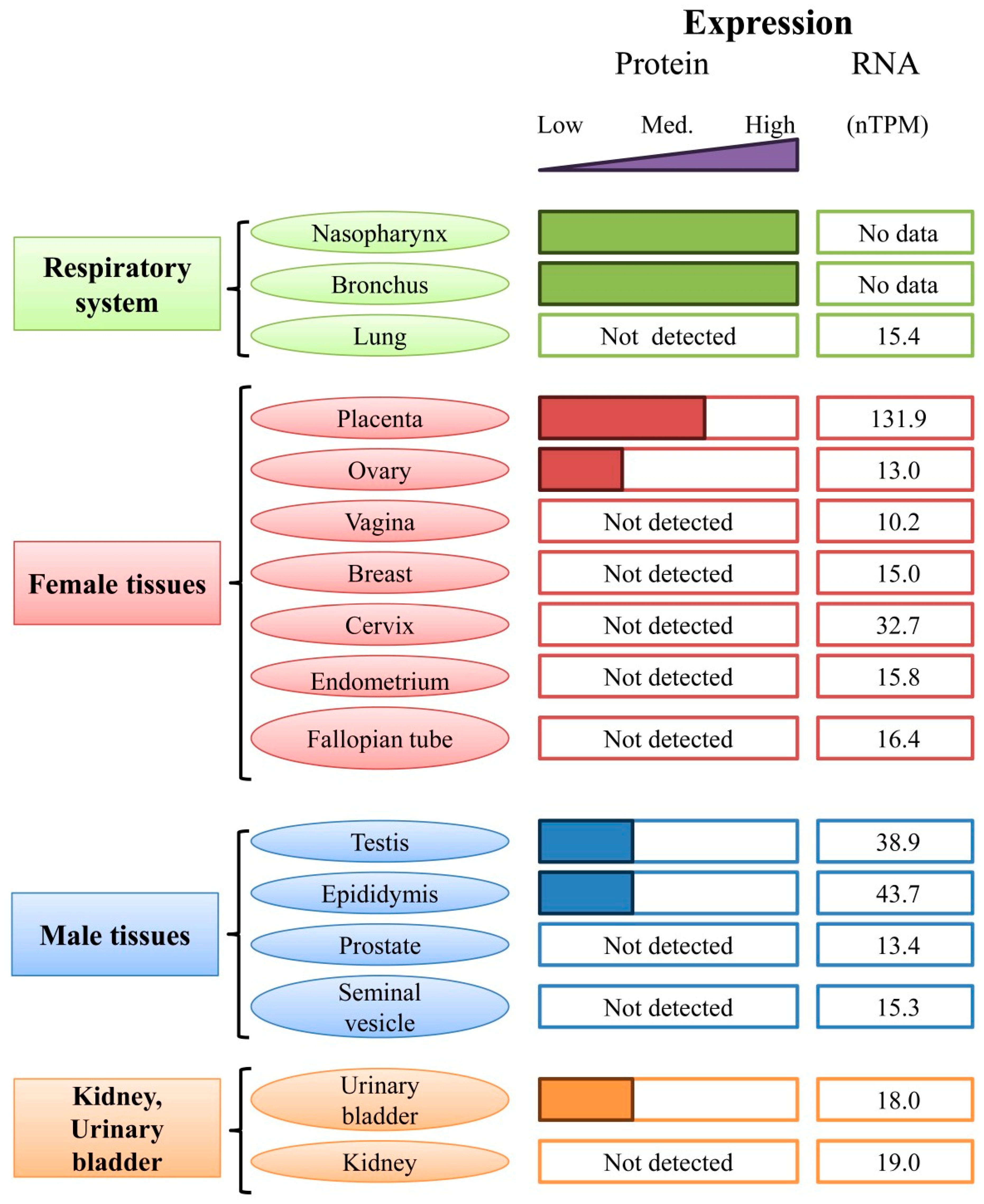
5.2. Expression of HMGB3 in Tissues and Organs
6. Conclusive Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mathew, C.G.; Goodwin, G.H.; Gooderham, K.; Walker, J.M.; Johns, E.W. A comparison of the high mobility group non-histone chromatin protein HMG 2 in chicken thymus and erythrocytes. Biochem. Biophys. Res. Commun. 1979, 87, 1243–1251. [Google Scholar] [CrossRef]
- Wilke, K.; Wiemann, S.; Gaul, R.; Gong, W.; Poustka, A. Isolation of human and mouse HMG2a cDNAs: Evidence for an HMG2a-specific 3′ untranslated region. Gene 1997, 198, 269–274. [Google Scholar] [CrossRef]
- Vaccari, T.; Beltrame, M.; Ferrari, S.; Bianchi, M.E. Hmg4, a new member of the Hmg1/2 gene family. Genomics 1998, 49, 247–252. [Google Scholar] [CrossRef]
- Goodwin, G.H.; Walker, J.M.; Johns, E.W. Studies on the degradation of high mobility group non-histone chromosomal proteins. Biochim. Biophys. Acta 1978, 519, 233–242. [Google Scholar] [CrossRef]
- Isackson, P.J.; Beaudoin, J.; Hermodson, M.A.; Reeck, G.R. Production of HMG-3 by limited trypsin digestion of purified high-mobility-group nonhistone chromatin proteins. Biochim. Biophys. Acta 1983, 748, 436–443. [Google Scholar] [CrossRef]
- Bustin, M. Regulation of DNA-dependent activities by the functional motifs of the high-mobility-group chromosomal proteins. Mol. Cell. Biol. 1999, 19, 5237–5246. [Google Scholar] [CrossRef]
- Bustin, M. Revised nomenclature for high mobility group (HMG) chromosomal proteins. Trends Biochem. Sci. 2001, 26, 152–153. [Google Scholar] [CrossRef]
- Read, C.M.; Cary, P.D.; Crane-Robinson, C.; Driscoll, P.C.; Norman, D.G. Solution structure of a DNA-binding domain from HMG1. Nucleic Acids Res. 1993, 21, 3427–3436. [Google Scholar] [CrossRef]
- Starkova, T.; Polyanichko, A.; Tomilin, A.N.; Chikhirzhina, E. Structure and Functions of HMGB2 Protein. Int. J. Mol. Sci. 2023, 24, 8334. [Google Scholar] [CrossRef]
- Reeves, R. High mobility group (HMG) proteins: Modulators of chromatin structure and DNA repair in mammalian cells. DNA Repair. 2015, 36, 122–136. [Google Scholar] [CrossRef]
- Chikhirzhina, E.; Starkova, T.; Beljajev, A.; Polyanichko, A.; Tomilin, A. Functional Diversity of Non-Histone Chromosomal Protein HmgB1. Int. J. Mol. Sci. 2020, 21, 7948. [Google Scholar] [CrossRef]
- Chikhirzhina, E.; Starkova, T.Y.; Polyanichko, A. The Structural Organization of the HMGB1 Nuclear Protein and Its Effect on the Formation of Ordered Supramolecular Complexes. Biophysics 2021, 66, 373–378. [Google Scholar] [CrossRef]
- Stros, M.; Launholt, D.; Grasser, K.D. The HMG-box: A versatile protein domain occurring in a wide variety of DNA-binding proteins. Cell. Mol. Life Sci. CMLS 2007, 64, 2590–2606. [Google Scholar] [CrossRef]
- Hamilton, D.J.; Hein, A.E.; Wuttke, D.S.; Batey, R.T. The DNA binding high mobility group box protein family functionally binds RNA. Wiley Interdiscip. Rev. RNA 2023, 14, e1778. [Google Scholar] [CrossRef]
- Malarkey, C.S.; Churchill, M.E. The high mobility group box: The ultimate utility player of a cell. Trends Biochem. Sci. 2012, 37, 553–562. [Google Scholar] [CrossRef]
- Murphy, E.C.; Zhurkin, V.B.; Louis, J.M.; Cornilescu, G.; Clore, G.M. Structural basis for SRY-dependent 46-X,Y sex reversal: Modulation of DNA bending by a naturally occurring point mutation. J. Mol. Biol. 2001, 312, 481–499. [Google Scholar] [CrossRef]
- Sinclair, A.H.; Berta, P.; Palmer, M.S.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J.; Foster, J.W.; Frischauf, A.M.; Lovell-Badge, R.; Goodfellow, P.N. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef]
- Leis, O.; Eguiara, A.; Lopez-Arribillaga, E.; Alberdi, M.J.; Hernandez-Garcia, S.; Elorriaga, K.; Pandiella, A.; Rezola, R.; Martin, A.G. Sox2 expression in breast tumours and activation in breast cancer stem cells. Oncogene 2012, 31, 1354–1365. [Google Scholar] [CrossRef]
- Grosschedl, R.; Giese, K.; Pagel, J. HMG domain proteins: Architectural elements in the assembly of nucleoprotein structures. Trends Genet. TIG 1994, 10, 94–100. [Google Scholar] [CrossRef]
- Love, J.J.; Li, X.; Case, D.A.; Giese, K.; Grosschedl, R.; Wright, P.E. Structural basis for DNA bending by the architectural transcription factor LEF-1. Nature 1995, 376, 791–795. [Google Scholar] [CrossRef]
- Chi, T. A BAF-centred view of the immune system. Nat. Rev. Immunol. 2004, 4, 965–977. [Google Scholar] [CrossRef]
- Lai, D.; Wan, M.; Wu, J.; Preston-Hurlburt, P.; Kushwaha, R.; Grundstrom, T.; Imbalzano, A.N.; Chi, T. Induction of TLR4-target genes entails calcium/calmodulin-dependent regulation of chromatin remodeling. Proc. Natl. Acad. Sci. USA 2009, 106, 1169–1174. [Google Scholar] [CrossRef]
- Catena, R.; Escoffier, E.; Caron, C.; Khochbin, S.; Martianov, I.; Davidson, I. HMGB4, a novel member of the HMGB family, is preferentially expressed in the mouse testis and localizes to the basal pole of elongating spermatids. Biol. Reprod. 2009, 80, 358–366. [Google Scholar] [CrossRef]
- Rouhiainen, A.; Zhao, X.; Vanttola, P.; Qian, K.; Kulesskiy, E.; Kuja-Panula, J.; Gransalke, K.; Gronholm, M.; Unni, E.; Meistrich, M.; et al. HMGB4 is expressed by neuronal cells and affects the expression of genes involved in neural differentiation. Sci. Rep. 2016, 6, 32960. [Google Scholar] [CrossRef]
- Li, W.; Zhu, J.; Lei, L.; Chen, C.; Liu, X.; Wang, Y.; Hong, X.; Yu, L.; Xu, H.; Zhu, X. The Seasonal and Stage-Specific Expression Patterns of HMGB2 Suggest Its Key Role in Spermatogenesis in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Biochem. Genet. 2022, 60, 2489–2502. [Google Scholar] [CrossRef]
- Stros, M. HMGB proteins: Interactions with DNA and chromatin. Biochim. Biophys. Acta 2010, 1799, 101–113. [Google Scholar] [CrossRef]
- Sullivan, G.J.; McStay, B. Dimerization and HMG box domains 1-3 present in Xenopus UBF are sufficient for its role in transcriptional enhancement. Nucleic Acids Res. 1998, 26, 3555–3561. [Google Scholar] [CrossRef]
- Decoville, M.; Giacomello, E.; Leng, M.; Locker, D. DSP1, an HMG-like protein, is involved in the regulation of homeotic genes. Genetics 2001, 157, 237–244. [Google Scholar] [CrossRef]
- Panday, A.; Grove, A. The high mobility group protein HMO1 functions as a linker histone in yeast. Epigenet. Chromatin 2016, 9, 13. [Google Scholar] [CrossRef]
- Bustin, M.; Reeves, R. High-mobility-group chromosomal proteins: Architectural components that facilitate chromatin function. Prog. Nucleic Acid. Res. Mol. Biol. 1996, 54, 35–100. [Google Scholar] [CrossRef]
- Murphy, F.V.t.; Churchill, M.E. Nonsequence-specific DNA recognition: A structural perspective. Structure 2000, 8, R83–R89. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, T.C.; Wilson, J.J.; Lippard, S.J. Monofunctional and higher-valent platinum anticancer agents. Inorg. Chem. 2013, 52, 12234–12249. [Google Scholar] [CrossRef] [PubMed]
- Totsingan, F.; Bell, A.J., Jr. Interaction of HMG proteins and H1 with hybrid PNA-DNA junctions. Protein Sci. A Publ. Protein Soc. 2013, 22, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Ban, T.; Wang, Z.; Choi, M.K.; Kawamura, T.; Negishi, H.; Nakasato, M.; Lu, Y.; Hangai, S.; Koshiba, R.; et al. HMGB proteins function as universal sentinels for nucleic-acid-mediated innate immune responses. Nature 2009, 462, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Raucci, A.; Di Maggio, S.; Scavello, F.; D’Ambrosio, A.; Bianchi, M.E.; Capogrossi, M.C. The Janus face of HMGB1 in heart disease: A necessary update. Cell. Mol. Life Sci. CMLS 2019, 76, 211–229. [Google Scholar] [CrossRef]
- Chikhirzhina, E.V.; Polyanichko, A.M.; Starkova, T.Y. Extranuclear functions of nonhistone protein HMGB1. Tsitologiya 2020, 62, 716–725. [Google Scholar] [CrossRef]
- Hock, R.; Furusawa, T.; Ueda, T.; Bustin, M. HMG chromosomal proteins in development and disease. Trends Cell Biol. 2007, 17, 72–79. [Google Scholar] [CrossRef]
- Fang, J.; Ge, X.; Xu, W.; Xie, J.; Qin, Z.; Shi, L.; Yin, W.; Bian, M.; Wang, H. Bioinformatics analysis of the prognosis and biological significance of HMGB1, HMGB2, and HMGB3 in gastric cancer. J. Cell. Physiol. 2020, 235, 3438–3446. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Wei, Y.T.; Zhao, K. The role of high mobility group protein B3 (HMGB3) in tumor proliferation and drug resistance. Mol. Cell. Biochem. 2021, 476, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Sehnal, D.; Bittrich, S.; Deshpande, M.; Svobodova, R.; Berka, K.; Bazgier, V.; Velankar, S.; Burley, S.K.; Koca, J.; Rose, A.S. Mol* Viewer: Modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Res. 2021, 49, W431–W437. [Google Scholar] [CrossRef]
- AlphaFold. Available online: https://alphafold.ebi.ac.uk/entry/ (accessed on 10 April 2024).
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Weir, H.M.; Kraulis, P.J.; Hill, C.S.; Raine, A.R.; Laue, E.D.; Thomas, J.O. Structure of the HMG box motif in the B-domain of HMG1. EMBO J. 1993, 12, 1311–1319. [Google Scholar] [CrossRef]
- Hardman, C.H.; Broadhurst, R.W.; Raine, A.R.; Grasser, K.D.; Thomas, J.O.; Laue, E.D. Structure of the A-domain of HMG1 and its interaction with DNA as studied by heteronuclear three- and four-dimensional NMR spectroscopy. Biochemistry 1995, 34, 16596–16607. [Google Scholar] [CrossRef]
- Broadhurst, R.W.; Hardman, C.H.; Thomas, J.O.; Laue, E.D. Backbone dynamics of the A-domain of HMG1 as studied by 15N NMR spectroscopy. Biochemistry 1995, 34, 16608–16617. [Google Scholar] [CrossRef]
- Kwak, M.S.; Kim, H.S.; Lee, B.; Kim, Y.H.; Son, M.; Shin, J.S. Immunological Significance of HMGB1 Post-Translational Modification and Redox Biology. Front. Immunol. 2020, 11, 1189. [Google Scholar] [CrossRef]
- Chen, R.; Kang, R.; Tang, D. The mechanism of HMGB1 secretion and release. Exp. Mol. Med. 2022, 54, 91–102. [Google Scholar] [CrossRef]
- Rowell, J.P.; Simpson, K.L.; Stott, K.; Watson, M.; Thomas, J.O. HMGB1-facilitated p53 DNA binding occurs via HMG-Box/p53 transactivation domain interaction, regulated by the acidic tail. Structure 2012, 20, 2014–2024. [Google Scholar] [CrossRef]
- RCSB. Available online: https://www.rcsb.org/structure/ (accessed on 20 May 2024).
- Masse, J.E.; Wong, B.; Yen, Y.M.; Allain, F.H.; Johnson, R.C.; Feigon, J. The S. cerevisiae architectural HMGB protein NHP6A complexed with DNA: DNA and protein conformational changes upon binding. J. Mol. Biol. 2002, 323, 263–284. [Google Scholar] [CrossRef]
- Rubio-Cosials, A.; Sidow, J.F.; Jimenez-Menendez, N.; Fernandez-Millan, P.; Montoya, J.; Jacobs, H.T.; Coll, M.; Bernado, P.; Sola, M. Human mitochondrial transcription factor A induces a U-turn structure in the light strand promoter. Nat. Struct. Mol. Biol. 2011, 18, 1281–1289. [Google Scholar] [CrossRef]
- Sanchez-Giraldo, R.; Acosta-Reyes, F.J.; Malarkey, C.S.; Saperas, N.; Churchill, M.E.; Campos, J.L. Two high-mobility group box domains act together to underwind and kink DNA. Acta Crystallogr. Sect. D Struct. Biol. 2015, 71, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Farid, R.S.; Bianchi, M.E.; Falciola, L.; Engelsberg, B.N.; Billings, P.C. Differential binding of HMG1, HMG2, and a single HMG box to cisplatin-damaged DNA. Toxicol. Appl. Pharmacol. 1996, 141, 532–539. [Google Scholar] [CrossRef]
- Bonaldi, T.; Talamo, F.; Scaffidi, P.; Ferrera, D.; Porto, A.; Bachi, A.; Rubartelli, A.; Agresti, A.; Bianchi, M.E. Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. EMBO J. 2003, 22, 5551–5560. [Google Scholar] [CrossRef] [PubMed]
- Ohndorf, U.M.; Rould, M.A.; He, Q.; Pabo, C.O.; Lippard, S.J. Basis for recognition of cisplatin-modified DNA by high-mobility-group proteins. Nature 1999, 399, 708–712. [Google Scholar] [CrossRef]
- Anggayasti, W.L.; Ogino, K.; Yamamoto, E.; Helmerhorst, E.; Yasuoka, K.; Mancera, R.L. The acidic tail of HMGB1 regulates its secondary structure and conformational flexibility: A circular dichroism and molecular dynamics simulation study. Comput. Struct. Biotechnol. J. 2020, 18, 1160–1172. [Google Scholar] [CrossRef] [PubMed]
- Knapp, S.; Muller, S.; Digilio, G.; Bonaldi, T.; Bianchi, M.E.; Musco, G. The long acidic tail of high mobility group box 1 (HMGB1) protein forms an extended and flexible structure that interacts with specific residues within and between the HMG boxes. Biochemistry 2004, 43, 11992–11997. [Google Scholar] [CrossRef]
- Marintcheva, B.; Marintchev, A.; Wagner, G.; Richardson, C.C. Acidic C-terminal tail of the ssDNA-binding protein of bacteriophage T7 and ssDNA compete for the same binding surface. Proc. Natl. Acad. Sci. USA 2008, 105, 1855–1860. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Xia, M.; Liu, C.; Zu, X.; Zhong, J. High Mobility Group A1 (HMGA1): Structure, Biological Function, and Therapeutic Potential. Int. J. Biol. Sci. 2022, 18, 4414–4431. [Google Scholar] [CrossRef]
- Belgrano, F.S.; de Abreu da Silva, I.C.; Bastos de Oliveira, F.M.; Fantappie, M.R.; Mohana-Borges, R. Role of the acidic tail of high mobility group protein B1 (HMGB1) in protein stability and DNA bending. PLoS ONE 2013, 8, e79572. [Google Scholar] [CrossRef]
- Anggayasti, W.L.; Mancera, R.L.; Bottomley, S.; Helmerhorst, E. The self-association of HMGB1 and its possible role in the binding to DNA and cell membrane receptors. FEBS Lett. 2017, 591, 282–294. [Google Scholar] [CrossRef]
- Stott, K.; Watson, M.; Howe, F.S.; Grossmann, J.G.; Thomas, J.O. Tail-mediated collapse of HMGB1 is dynamic and occurs via differential binding of the acidic tail to the A and B domains. J. Mol. Biol. 2010, 403, 706–722. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.B.; Thomas, J.O. The effect of the acidic tail on the DNA-binding properties of the HMG1,2 class of proteins: Insights from tail switching and tail removal. J. Mol. Biol. 2000, 304, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Li, Y.; Chao, F.; Huang, G.; He, F. Amino acid residues 201-205 in C-terminal acidic tail region plays a crucial role in antibacterial activity of HMGB1. J. Biomed. Sci. 2009, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Borde, C.; Dillard, C.; L’Honore, A.; Quignon, F.; Hamon, M.; Marchand, C.H.; Faccion, R.S.; Costa, M.G.S.; Pramil, E.; Larsen, A.K.; et al. The C-Terminal Acidic Tail Modulates the Anticancer Properties of HMGB1. Int. J. Mol. Sci. 2022, 23, 7865. [Google Scholar] [CrossRef]
- Roesgaard, M.A.; Lundsgaard, J.E.; Newcombe, E.A.; Jacobsen, N.L.; Pesce, F.; Tranchant, E.E.; Lindemose, S.; Prestel, A.; Hartmann-Petersen, R.; Lindorff-Larsen, K.; et al. Deciphering the Alphabet of Disorder-Glu and Asp Act Differently on Local but Not Global Properties. Biomolecules 2022, 12, 1426. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef] [PubMed]
- Venereau, E.; Casalgrandi, M.; Schiraldi, M.; Antoine, D.J.; Cattaneo, A.; De Marchis, F.; Liu, J.; Antonelli, A.; Preti, A.; Raeli, L.; et al. Mutually exclusive redox forms of HMGB1 promote cell recruitment or proinflammatory cytokine release. J. Exp. Med. 2012, 209, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y. High mobility group proteins and their post-translational modifications. Biochim. Biophys. Acta 2008, 1784, 1159–1166. [Google Scholar] [CrossRef]
- Oh, Y.J.; Youn, J.H.; Ji, Y.; Lee, S.E.; Lim, K.J.; Choi, J.E.; Shin, J.S. HMGB1 is phosphorylated by classical protein kinase C and is secreted by a calcium-dependent mechanism. J. Immunol. 2009, 182, 5800–5809. [Google Scholar] [CrossRef]
- Starkova, T.Y.; Polyanichko, A.M.; Artamonova, T.O.; Tsimokha, A.S.; Tomilin, A.N.; Chikhirzhina, E.V. Structural Characteristics of High-Mobility Group Proteins HMGB1 and HMGB2 and Their Interaction with DNA. Int. J. Mol. Sci. 2023, 24, 3577. [Google Scholar] [CrossRef]
- Ito, I.; Fukazawa, J.; Yoshida, M. Post-translational methylation of high mobility group box 1 (HMGB1) causes its cytoplasmic localization in neutrophils. J. Biol. Chem. 2007, 282, 16336–16344. [Google Scholar] [CrossRef]
- Chen, L.; Long, X.; Xu, Q.; Tan, J.; Wang, G.; Cao, Y.; Wei, J.; Luo, H.; Zhu, H.; Huang, L.; et al. Elevated serum levels of S100A8/A9 and HMGB1 at hospital admission are correlated with inferior clinical outcomes in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 992–994. [Google Scholar] [CrossRef]
- De Martinis, M.; Ginaldi, L.; Sirufo, M.M.; Pioggia, G.; Calapai, G.; Gangemi, S.; Mannucci, C. Alarmins in Osteoporosis, RAGE, IL-1, and IL-33 Pathways: A Literature Review. Medicina 2020, 56, 138. [Google Scholar] [CrossRef]
- Ginaldi, L.; De Martinis, M.; Saitta, S.; Sirufo, M.M.; Mannucci, C.; Casciaro, M.; Ciccarelli, F.; Gangemi, S. Interleukin-33 serum levels in postmenopausal women with osteoporosis. Sci. Rep. 2019, 9, 3786. [Google Scholar] [CrossRef]
- Ugrinova, I.; Pashev, I.G.; Pasheva, E.A. Nucleosome binding properties and Co-remodeling activities of native and in vivo acetylated HMGB-1 and HMGB-2 proteins. Biochemistry 2009, 48, 6502–6507. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, Y. HMG modifications and nuclear function. Biochim. Biophys. Acta 2010, 1799, 28–36. [Google Scholar] [CrossRef]
- Stott, K.; Tang, G.S.; Lee, K.B.; Thomas, J.O. Structure of a complex of tandem HMG boxes and DNA. J. Mol. Biol. 2006, 360, 90–104. [Google Scholar] [CrossRef]
- Gong, W.; Guo, Y.; Yuan, H.; Hu, X.; Chai, R.; Zheng, B.; Wan, Z.; Tu, S. HMGB3 is a Potential Therapeutic Target by Affecting the Migration and Proliferation of Colorectal Cancer. Front. Cell Dev. Biol. 2022, 10, 891482. [Google Scholar] [CrossRef]
- Zheng, M.; Wu, L.; Xiao, R.; Zhou, Y.; Cai, J.; Chen, W.; Chen, C.; Sun, K.; Shen, S. Integrated analysis of coexpression and a tumor-specific ceRNA network revealed a potential prognostic biomarker in breast cancer. Transl. Cancer Res. 2023, 12, 949–964. [Google Scholar] [CrossRef]
- Ramachandran, C.; Yau, P.; Bradbury, E.M.; Shyamala, G.; Yasuda, H.; Walsh, D.A. Phosphorylation of high-mobility-group proteins by the calcium-phospholipid-dependent protein kinase and the cyclic AMP-dependent protein kinase. J. Biol. Chem. 1984, 259, 13495–13503. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kwak, M.S.; Park, J.B.; Lee, S.A.; Choi, J.E.; Cho, H.S.; Shin, J.S. N-linked glycosylation plays a crucial role in the secretion of HMGB1. J. Cell Sci. 2016, 129, 29–38. [Google Scholar] [CrossRef]
- Husnjak, K.; Dikic, I. Ubiquitin-binding proteins: Decoders of ubiquitin-mediated cellular functions. Annu. Rev. Biochem. 2012, 81, 291–322. [Google Scholar] [CrossRef]
- Shilatifard, A. Chromatin modifications by methylation and ubiquitination: Implications in the regulation of gene expression. Annu. Rev. Biochem. 2006, 75, 243–269. [Google Scholar] [CrossRef]
- Acconcia, F.; Sigismund, S.; Polo, S. Ubiquitin in trafficking: The network at work. Exp. Cell Res. 2009, 315, 1610–1618. [Google Scholar] [CrossRef]
- Piper, R.C.; Dikic, I.; Lukacs, G.L. Ubiquitin-dependent sorting in endocytosis. Cold Spring Harb. Perspect. Biol. 2014, 6, a016808. [Google Scholar] [CrossRef]
- Finley, D. Recognition and processing of ubiquitin-protein conjugates by the proteasome. Annu. Rev. Biochem. 2009, 78, 477–513. [Google Scholar] [CrossRef]
- Khaminets, A.; Behl, C.; Dikic, I. Ubiquitin-Dependent And Independent Signals In Selective Autophagy. Trends Cell Biol. 2016, 26, 6–16. [Google Scholar] [CrossRef]
- Dong, S.; Pan, J.; Shen, Y.B.; Zhu, L.X.; Chen, L.; Zhu, F.; Li, H.; Shen, H.X.; Xia, Q.; Wu, Y.J.; et al. SYT7 plays a role in promoting thyroid cancer by mediating HMGB3 ubiquitination. Endocr.-Relat. Cancer 2022, 29, 175–189. [Google Scholar] [CrossRef]
- UbiBrowser. Available online: http://ubibrowser.bio-it.cn/ubibrowser_v3/ (accessed on 25 May 2024).
- Hu, J.; Ding, R.; Liu, S.; Wang, J.; Li, J.; Shang, Y. Hypermethylation of RNF125 promotes autophagy-induced oxidative stress in asthma by increasing HMGB1 stability. iScience 2023, 26, 107503. [Google Scholar] [CrossRef]
- Du, J.; Ji, X.; Xu, B.; Du, Q.; Li, Y.; Zhou, B.; Liu, X.; Xu, Z.; Jiang, Y.; Kou, B.; et al. Ubiquitination of cytoplasmic HMGB1 by RNF186 regulates hepatic lipophagy in non-alcoholic fatty liver disease. Metabolism 2024, 152, 155769. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, Q.; Wang, M.; Sun, F.; Qiao, P.; Jiang, A.; Ren, C.; Yu, Z.; Yang, T. CHIP induces ubiquitination and degradation of HMGB1 to regulate glycolysis in ovarian endometriosis. Cell. Mol. Life Sci. CMLS 2022, 80, 13. [Google Scholar] [CrossRef]
- PhosphoSite. Available online: https://www.phosphosite.org/ (accessed on 25 May 2024).
- Azevedo, C.; Saiardi, A. Why always lysine? The ongoing tale of one of the most modified amino acids. Adv. Biol. Regul. 2016, 60, 144–150. [Google Scholar] [CrossRef]
- Qin, X.; Niu, Z.; Chen, H.; Hu, Y. Macrophage-derived exosomal HMGB3 regulates silica-induced pulmonary inflammation by promoting M1 macrophage polarization and recruitment. Part. Fibre Toxicol. 2024, 21, 12. [Google Scholar] [CrossRef]
- Hu, P.; Yan, T.; Lv, S.; Ye, M.; Wu, M.; Fang, H.; Xiao, B. Exosomal HMGB3 released by glioma cells confers the activation of NLRP3 inflammasome and pyroptosis in tumor-associated macrophages. Tissue Cell 2024, 88, 102406. [Google Scholar] [CrossRef]
- Smith, V.L.; Jackson, L.; Schorey, J.S. Ubiquitination as a Mechanism To Transport Soluble Mycobacterial and Eukaryotic Proteins to Exosomes. J. Immunol. 2015, 195, 2722–2730. [Google Scholar] [CrossRef]
- Barreiro-Alonso, A.; Lamas-Maceiras, M.; Rodriguez-Belmonte, E.; Vizoso-Vazquez, A.; Quindos, M.; Cerdan, M.E. High Mobility Group B Proteins, Their Partners, and Other Redox Sensors in Ovarian and Prostate Cancer. Oxidative Med. Cell. Longev. 2016, 2016, 5845061. [Google Scholar] [CrossRef]
- Antoine, D.J.; Harris, H.E.; Andersson, U.; Tracey, K.J.; Bianchi, M.E. A systematic nomenclature for the redox states of high mobility group box (HMGB) proteins. Mol. Med. 2014, 20, 135–137. [Google Scholar] [CrossRef]
- Hoppe, G.; Talcott, K.E.; Bhattacharya, S.K.; Crabb, J.W.; Sears, J.E. Molecular basis for the redox control of nuclear transport of the structural chromatin protein Hmgb1. Exp. Cell Res. 2006, 312, 3526–3538. [Google Scholar] [CrossRef]
- Andersson, U.; Ottestad, W.; Tracey, K.J. Extracellular HMGB1: A therapeutic target in severe pulmonary inflammation including COVID-19? Mol. Med. 2020, 26, 42. [Google Scholar] [CrossRef]
- Morris, G.; Bortolasci, C.C.; Puri, B.K.; Olive, L.; Marx, W.; O’Neil, A.; Athan, E.; Carvalho, A.F.; Maes, M.; Walder, K.; et al. The pathophysiology of SARS-CoV-2: A suggested model and therapeutic approach. Life Sci. 2020, 258, 118166. [Google Scholar] [CrossRef]
- Polanska, E.; Pospisilova, S.; Stros, M. Binding of histone H1 to DNA is differentially modulated by redox state of HMGB1. PLoS ONE 2014, 9, e89070. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Q.; Liang, G.; Liang, X.; Luo, B. Overexpression of HMGB3 and its prognostic value in breast cancer. Front. Oncol. 2022, 12, 1048921. [Google Scholar] [CrossRef]
- Niu, L.; Yang, W.; Duan, L.; Wang, X.; Li, Y.; Xu, C.; Liu, C.; Zhang, Y.; Zhou, W.; Liu, J.; et al. Biological functions and theranostic potential of HMGB family members in human cancers. Ther. Adv. Med. Oncol. 2020, 12, 1758835920970850. [Google Scholar] [CrossRef]
- Polyanichko, A.M.; Davydenko, S.G.; Chikhirzhina, E.V.; Vorob’ev, V.I. Interaction of superhelical DNA with the nonhistone protein HMG1. Tsitologiia 2000, 42, 787–793. [Google Scholar]
- Polyanichko, A.M.; Leonenko, Z.V.; Kramb, D.; Vieser, G.; Vorob’ev, V.I.; Chikhirzhina, E.V. Visualization of DNA Complexes with HMGB1 and Its C-Truncated Form HMGB1(A+B). Biophysics 2008, 53, 202–206. [Google Scholar] [CrossRef]
- Park, S.; Lippard, S.J. Redox state-dependent interaction of HMGB1 and cisplatin-modified DNA. Biochemistry 2011, 50, 2567–2574. [Google Scholar] [CrossRef]
- Jung, Y.; Lippard, S.J. Nature of full-length HMGB1 binding to cisplatin-modified DNA. Biochemistry 2003, 42, 2664–2671. [Google Scholar] [CrossRef]
- Wang, Q.; Zeng, M.; Wang, W.; Tang, J. The HMGB1 acidic tail regulates HMGB1 DNA binding specificity by a unique mechanism. Biochem. Biophys. Res. Commun. 2007, 360, 14–19. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Ngo, H.B.; Lovely, G.A.; Phillips, R.; Chan, D.C. Distinct structural features of TFAM drive mitochondrial DNA packaging versus transcriptional activation. Nat. Commun. 2014, 5, 3077. [Google Scholar] [CrossRef]
- Chakraborty, A.; Lyonnais, S.; Battistini, F.; Hospital, A.; Medici, G.; Prohens, R.; Orozco, M.; Vilardell, J.; Sola, M. DNA structure directs positioning of the mitochondrial genome packaging protein Abf2p. Nucleic Acids Res. 2017, 45, 951–967. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R. Nuclear functions of the HMG proteins. Biochim. Biophys. Acta 2010, 1799, 3–14. [Google Scholar] [CrossRef]
- Wang, J.; Tochio, N.; Takeuchi, A.; Uewaki, J.; Kobayashi, N.; Tate, S. Redox-sensitive structural change in the A-domain of HMGB1 and its implication for the binding to cisplatin modified DNA. Biochem. Biophys. Res. Commun. 2013, 441, 701–706. [Google Scholar] [CrossRef]
- Mukherjee, A.; Huynh, V.; Gaines, K.; Reh, W.A.; Vasquez, K.M. Targeting the High-Mobility Group Box 3 Protein Sensitizes Chemoresistant Ovarian Cancer Cells to Cisplatin. Cancer Res. 2019, 79, 3185–3191. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Du, Z.; Luo, Q.; Zhao, Y.; Wang, Y.; Wu, K.; Jia, F.; Zhang, Y.; Wang, F. Proteomic Strategy for Identification of Proteins Responding to Cisplatin-Damaged DNA. Anal. Chem. 2019, 91, 6035–6042. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, Y.; Liu, A.; Tang, X. MiR-27b is epigenetically downregulated in tamoxifen resistant breast cancer cells due to promoter methylation and regulates tamoxifen sensitivity by targeting HMGB3. Biochem. Biophys. Res. Commun. 2016, 477, 768–773. [Google Scholar] [CrossRef]
- Guggenheim, E.R.; Xu, D.; Zhang, C.X.; Chang, P.V.; Lippard, S.J. Photoaffinity isolation and identification of proteins in cancer cell extracts that bind to platinum-modified DNA. ChemBioChem A Eur. J. Chem. Biol. 2009, 10, 141–157. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Sui, S.; Hua, Y.; Zhao, A.; Tian, X.; Wang, R.; Guo, W.; Yu, W.; Zou, K.; et al. Targeting HMGB3/hTERT axis for radioresistance in cervical cancer. J. Exp. Clin. Cancer Res. CR 2020, 39, 243. [Google Scholar] [CrossRef]
- Saldivar, J.S.; Wu, X.; Follen, M.; Gershenson, D. Nucleotide excision repair pathway review I: Implications in ovarian cancer and platinum sensitivity. Gynecol. Oncol. 2007, 107, S56–S71. [Google Scholar] [CrossRef]
- Chu, G. Cellular responses to cisplatin. The roles of DNA-binding proteins and DNA repair. J. Biol. Chem. 1994, 269, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Zhao, D.; Wang, C.; Wang, H.; Guan, X.; Gao, Y.; Zhang, X.; Zhang, N.; Chen, R. Characterization of RNA-binding proteins in the cell nucleus and cytoplasm. Anal. Chim. Acta 2021, 1168, 338609. [Google Scholar] [CrossRef]
- Khoury, G.; Lee, M.Y.; Ramarathinam, S.H.; McMahon, J.; Purcell, A.W.; Sonza, S.; Lewin, S.R.; Purcell, D.F.J. The RNA-Binding Proteins SRP14 and HMGB3 Control HIV-1 Tat mRNA Processing and Translation During HIV-1 Latency. Front. Genet. 2021, 12, 680725. [Google Scholar] [CrossRef] [PubMed]
- IntAct. Available online: https://www.ebi.ac.uk/intact/search?query=ENSG00000029993 (accessed on 7 April 2024).
- McKinney, K.; Prives, C. Efficient specific DNA binding by p53 requires both its central and C-terminal domains as revealed by studies with high-mobility group 1 protein. Mol. Cell. Biol. 2002, 22, 6797–6808. [Google Scholar] [CrossRef] [PubMed]
- Stros, M.; Kucirek, M.; Sani, S.A.; Polanska, E. HMGB1-mediated DNA bending: Distinct roles in increasing p53 binding to DNA and the transactivation of p53-responsive gene promoters. Biochim. Biophys. Acta. Gene Regul. Mech. 2018, 1861, 200–210. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, A. ST8SIA6-AS1 promotes hepatocellular carcinoma by absorbing miR-5195-3p to regulate HOXB6. Cancer Biol. Ther. 2020, 21, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Giampaolo, A.; Felli, N.; Diverio, D.; Morsilli, O.; Samoggia, P.; Breccia, M.; Lo Coco, F.; Peschle, C.; Testa, U. Expression pattern of HOXB6 homeobox gene in myelomonocytic differentiation and acute myeloid leukemia. Leukemia 2002, 16, 1293–1301. [Google Scholar] [CrossRef]
- Lill, M.C.; Fuller, J.F.; Herzig, R.; Crooks, G.M.; Gasson, J.C. The role of the homeobox gene, HOX B7, in human myelomonocytic differentiation. Blood 1995, 85, 692–697. [Google Scholar] [CrossRef]
- Fujino, T.; Yamazaki, Y.; Largaespada, D.A.; Jenkins, N.A.; Copeland, N.G.; Hirokawa, K.; Nakamura, T. Inhibition of myeloid differentiation by Hoxa9, Hoxb8, and Meis homeobox genes. Exp. Hematol. 2001, 29, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, M.J.; Curtis, D.J.; Kirby, M.R.; Garrett-Beal, L.J.; Seidel, N.E.; Cline, A.P.; Bodine, D.M. Hmgb3: An HMG-box family member expressed in primitive hematopoietic cells that inhibits myeloid and B-cell differentiation. Blood 2003, 102, 1298–1306. [Google Scholar] [CrossRef]
- Kappen, C. Disruption of the homeobox gene Hoxb-6 in mice results in increased numbers of early erythrocyte progenitors. Am. J. Hematol. 2000, 65, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Qi, G.; Han, F.; Lu, W.; Peng, J.; Li, R.; Yan, S.; Yuan, C.; Kong, B. HMGB3 promotes PARP inhibitor resistance through interacting with PARP1 in ovarian cancer. Cell Death Dis. 2022, 13, 263. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Nace, G.W.; McDonald, K.A.; Tai, S.; Klune, J.R.; Rosborough, B.R.; Ding, Q.; Loughran, P.; Zhu, X.; Beer-Stolz, D.; et al. Hepatocyte-specific high-mobility group box 1 deletion worsens the injury in liver ischemia/reperfusion: A role for intracellular high-mobility group box 1 in cellular protection. Hepatology 2014, 59, 1984–1997. [Google Scholar] [CrossRef] [PubMed]
- Franzese, E.; Centonze, S.; Diana, A.; Carlino, F.; Guerrera, L.P.; Di Napoli, M.; De Vita, F.; Pignata, S.; Ciardiello, F.; Orditura, M. PARP inhibitors in ovarian cancer. Cancer Treat. Rev. 2019, 73, 1–9. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, D.M.; Krivak, T.C.; Kabil, N.; Munley, J.; Moore, K.N. PARP Inhibitors in Ovarian Cancer: A Review. Target. Oncol. 2023, 18, 471–503. [Google Scholar] [CrossRef] [PubMed]
- Ray Chaudhuri, A.; Nussenzweig, A. The multifaceted roles of PARP1 in DNA repair and chromatin remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 610–621. [Google Scholar] [CrossRef]
- Ma, H.; Qi, G.; Han, F.; Gai, P.; Peng, J.; Kong, B. HMGB3 promotes the malignant phenotypes and stemness of epithelial ovarian cancer through the MAPK/ERK signaling pathway. Cell Commun. Signal. CCS 2023, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xu, M.; Li, X.; Weng, X.; Su, Z.; Zhang, M.; Tan, J.; Zeng, H.; Li, X.; Nie, L.; et al. SOX9 and HMGB3 co-operatively transactivate NANOG and promote prostate cancer progression. Prostate 2023, 83, 440–453. [Google Scholar] [CrossRef] [PubMed]
- Leung, V.Y.; Gao, B.; Leung, K.K.; Melhado, I.G.; Wynn, S.L.; Au, T.Y.; Dung, N.W.; Lau, J.Y.; Mak, A.C.; Chan, D.; et al. SOX9 governs differentiation stage-specific gene expression in growth plate chondrocytes via direct concomitant transactivation and repression. PLoS Genet. 2011, 7, e1002356. [Google Scholar] [CrossRef]
- Yao, C.; Su, L.; Shan, J.; Zhu, C.; Liu, L.; Liu, C.; Xu, Y.; Yang, Z.; Bian, X.; Shao, J.; et al. IGF/STAT3/NANOG/Slug Signaling Axis Simultaneously Controls Epithelial-Mesenchymal Transition and Stemness Maintenance in Colorectal Cancer. Stem Cells 2016, 34, 820–831. [Google Scholar] [CrossRef]
- Lu, G.; Li, Y.; Ma, Y.; Lu, J.; Chen, Y.; Jiang, Q.; Qin, Q.; Zhao, L.; Huang, Q.; Luo, Z.; et al. Long noncoding RNA LINC00511 contributes to breast cancer tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J. Exp. Clin. Cancer Res. CR 2018, 37, 289. [Google Scholar] [CrossRef] [PubMed]
- Jeter, C.R.; Liu, B.; Liu, X.; Chen, X.; Liu, C.; Calhoun-Davis, T.; Repass, J.; Zaehres, H.; Shen, J.J.; Tang, D.G. NANOG promotes cancer stem cell characteristics and prostate cancer resistance to androgen deprivation. Oncogene 2011, 30, 3833–3845. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Kang, J.; Ma, Y.; Wang, Q.; Li, W.; Su, J.; Zhang, X. Plasmacytoma variant translocation 1 inhibits miR-515-5p function and modulates high mobility group B3 to promote the growth of prostate cancer cells. Andrology 2023, 11, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Sgarra, R.; Furlan, C.; Zammitti, S.; Lo Sardo, A.; Maurizio, E.; Di Bernardo, J.; Giancotti, V.; Manfioletti, G. Interaction proteomics of the HMGA chromatin architectural factors. Proteomics 2008, 8, 4721–4732. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Zhang, S.; Fan, Z.; Che, J.; Li, S.; Li, Y.; Pan, K.; Lv, J.; Wang, C.; Zhang, T.; et al. Prognostic model construction and immune microenvironment analysis of esophageal cancer based on gene expression data and microRNA target genes. Transl. Cancer Res. 2023, 12, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Huang, Y.; Yuan, W.; Wu, X.; Shi, H.; Lu, M.; Xu, A. Expression, tumor immune infiltration, and prognostic impact of HMGs in gastric cancer. Front. Oncol. 2022, 12, 1056917. [Google Scholar] [CrossRef] [PubMed]
- Brien, M.E.; Baker, B.; Duval, C.; Gaudreault, V.; Jones, R.L.; Girard, S. Alarmins at the maternal-fetal interface: Involvement of inflammation in placental dysfunction and pregnancy complications (1). Can. J. Physiol. Pharmacol. 2019, 97, 206–212. [Google Scholar] [CrossRef]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in plant innate immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef]
- Venereau, E.; Ceriotti, C.; Bianchi, M.E. DAMPs from Cell Death to New Life. Front. Immunol. 2015, 6, 422. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lv, H.J.; Deng, X.Y.; Chen, P.; Garstka, M.A.; Shi, B.Y.; Fu, J. Translocated HMGB3 is involved in papillary thyroid cancer progression by activating cytoplasmic TLR3 and transmembrane TREM1. Cell Cycle 2023, 22, 2584–2601. [Google Scholar] [CrossRef]
- Tirone, M.; Tran, N.L.; Ceriotti, C.; Gorzanelli, A.; Canepari, M.; Bottinelli, R.; Raucci, A.; Di Maggio, S.; Santiago, C.; Mellado, M.; et al. High mobility group box 1 orchestrates tissue regeneration via CXCR4. J. Exp. Med. 2018, 215, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Tang, J.; Zheng, P.; Liu, Y. CD24 and Siglec-10 selectively repress tissue damage-induced immune responses. Science 2009, 323, 1722–1725. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R.; Zeh, H.J., 3rd; Lotze, M.T. High-mobility group box 1, oxidative stress, and disease. Antioxid. Redox Signal. 2011, 14, 1315–1335. [Google Scholar] [CrossRef] [PubMed]
- Schiraldi, M.; Raucci, A.; Munoz, L.M.; Livoti, E.; Celona, B.; Venereau, E.; Apuzzo, T.; De Marchis, F.; Pedotti, M.; Bachi, A.; et al. HMGB1 promotes recruitment of inflammatory cells to damaged tissues by forming a complex with CXCL12 and signaling via CXCR4. J. Exp. Med. 2012, 209, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Mantonico, M.V.; De Leo, F.; Quilici, G.; Colley, L.S.; De Marchis, F.; Crippa, M.; Mezzapelle, R.; Schulte, T.; Zucchelli, C.; Pastorello, C.; et al. The acidic intrinsically disordered region of the inflammatory mediator HMGB1 mediates fuzzy interactions with CXCL12. Nat. Commun. 2024, 15, 1201. [Google Scholar] [CrossRef] [PubMed]
- Paudel, Y.N.; Angelopoulou, E.; Piperi, C.; Balasubramaniam, V.; Othman, I.; Shaikh, M.F. Enlightening the role of high mobility group box 1 (HMGB1) in inflammation: Updates on receptor signalling. Eur. J. Pharmacol. 2019, 858, 172487. [Google Scholar] [CrossRef]
- Aljohani, A.I.; Alsaeed, S.A.; Toss, M.S.; Raafat, S.A.; Green, A.R.; Rakha, E.A. The expression of high mobility group protein 3 (HMGB3) in breast cancer with emphasis on its role in lymphovascular invasion. Am. J. Cancer Res. 2023, 13, 5334–5351. [Google Scholar] [PubMed]
- Lv, Z.; Zhang, Z.; Wei, Z.; Li, C.; Shao, Y.; Zhang, W.; Zhao, X.; Xiong, J. HMGB3 modulates ROS production via activating TLR cascade in Apostichopus japonicus. Dev. Comp. Immunol. 2017, 77, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Yanai, H.; Ban, T.; Taniguchi, T. Essential role of high-mobility group box proteins in nucleic acid-mediated innate immune responses. J. Intern. Med. 2011, 270, 301–308. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Lumpp, T.; Stosser, S.; Fischer, F.; Hartwig, A.; Koberle, B. Role of Epigenetics for the Efficacy of Cisplatin. Int. J. Mol. Sci. 2024, 25, 1130. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yan, W.; Qian, N.; Han, Q.; Zhang, W.; Dai, G. Paired box 5 increases the chemosensitivity of esophageal squamous cell cancer cells by promoting p53 signaling activity. Chin. Med. J. 2022, 135, 606–618. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, M.J.; Kirby, M.R.; Bodine, D.M. Hmgb3 regulates the balance between hematopoietic stem cell self-renewal and differentiation. Proc. Natl. Acad. Sci. USA 2006, 103, 13783–13788. [Google Scholar] [CrossRef] [PubMed]
- Huilgol, D.; Venkataramani, P.; Nandi, S.; Bhattacharjee, S. Transcription Factors That Govern Development and Disease: An Achilles Heel in Cancer. Genes 2019, 10, 794. [Google Scholar] [CrossRef] [PubMed]
- Chi, N.; Epstein, J.A. Getting your Pax straight: Pax proteins in development and disease. Trends Genet. TIG 2002, 18, 41–47. [Google Scholar] [CrossRef]
- Lee, O.H.; Kim, H.; He, Q.; Baek, H.J.; Yang, D.; Chen, L.Y.; Liang, J.; Chae, H.K.; Safari, A.; Liu, D.; et al. Genome-wide YFP fluorescence complementation screen identifies new regulators for telomere signaling in human cells. Mol. Cell. Proteom. MCP 2011, 10, M110.001628. [Google Scholar] [CrossRef] [PubMed]
- Petit, A.; Ragu, C.; Della-Valle, V.; Mozziconacci, M.J.; Lafage-Pochitaloff, M.; Soler, G.; Schluth, C.; Radford, I.; Ottolenghi, C.; Bernard, O.A.; et al. NUP98-HMGB3: A novel oncogenic fusion. Leukemia 2010, 24, 654–658. [Google Scholar] [CrossRef]
- Manhas, A.; Tripathi, D.; Jagavelu, K. Involvement of HIF1alpha/Reg protein in the regulation of HMGB3 in myocardial infarction. Vasc. Pharmacol. 2023, 152, 107197. [Google Scholar] [CrossRef]
- Gu, J.; Xu, T.; Huang, Q.H.; Zhang, C.M.; Chen, H.Y. HMGB3 silence inhibits breast cancer cell proliferation and tumor growth by interacting with hypoxia-inducible factor 1alpha. Cancer Manag. Res. 2019, 11, 5075–5089. [Google Scholar] [CrossRef]
- Gu, J.; Xu, T.; Zhang, C.M.; Chen, H.Y.; Huang, Q.H.; Zhang, Q. HMGB3 small interfere RNA suppresses mammosphere formation of MDA-MB-231 cells by down-regulating expression of HIF1alpha. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9506–9516. [Google Scholar] [CrossRef]
- Tang, H.R.; Luo, X.Q.; Xu, G.; Wang, Y.; Feng, Z.J.; Xu, H.; Shi, Y.W.; Zhang, Q.; Wu, L.G.; Xue, C.Q.; et al. High mobility group-box 3 overexpression is associated with poor prognosis of resected gastric adenocarcinoma. World J. Gastroenterol. 2012, 18, 7319–7326. [Google Scholar] [CrossRef]
- Guo, S.; Wang, Y.; Gao, Y.; Zhang, Y.; Chen, M.; Xu, M.; Hu, L.; Jing, Y.; Jing, F.; Li, C.; et al. Knockdown of High Mobility Group-Box 3 (HMGB3) Expression Inhibits Proliferation, Reduces Migration, and Affects Chemosensitivity in Gastric Cancer Cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 3951–3960. [Google Scholar] [CrossRef]
- Yang, H.; He, P.; Luo, W.; Liu, S.; Yang, Y. circRNA TATA-box binding protein associated factor 15 acts as an oncogene to facilitate bladder cancer progression through targeting miR-502-5p/high mobility group box 3. Mol. Carcinog. 2024, 63, 629–646. [Google Scholar] [CrossRef]
- Zhuang, S.; Yu, X.; Lu, M.; Li, Y.; Ding, N.; Ding, Y. High mobility group box 3 promotes cervical cancer proliferation by regulating Wnt/beta-catenin pathway. J. Gynecol. Oncol. 2020, 31, e91. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; He, S.; Zhang, D.; Lv, Z.; Yu, J.; Li, Q.; Li, M.; Guo, W.; Qi, F. LINC00857 promotes colorectal cancer progression by sponging miR-150-5p and upregulating HMGB3 (high mobility group box 3) expression. Bioengineered 2021, 12, 12107–12122. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Xi, Y.; Zhang, Z.; Hao, Y.; Wu, F.; Bian, B.; Hao, G.; Li, W.; Zhang, S. Hsa_circ_0060937 accelerates non-small cell lung cancer progression via modulating miR-195-5p/HMGB3 pathway. Cell Cycle 2021, 20, 2040–2052. [Google Scholar] [CrossRef]
- Orang, A.; Ali, S.R.; Petersen, J.; McKinnon, R.A.; Aloia, A.L.; Michael, M.Z. A functional screen with metformin identifies microRNAs that regulate metabolism in colorectal cancer cells. Sci. Rep. 2022, 12, 2889. [Google Scholar] [CrossRef] [PubMed]
- Voropaeva, E.N.; Berezina, O.V.; Churkina, M.I.; Pospelova, T.I.; Lyzlova, A.A.; Maksimov, V.N. Aberrant expression and methylation of individual microRNAs genes in lymphoproliferative diseases: A literature review. J. Sib. Med. Sci. 2021, 4, 108–133. [Google Scholar] [CrossRef]
- Lindberg, R.L.; Hoffmann, F.; Mehling, M.; Kuhle, J.; Kappos, L. Altered expression of miR-17-5p in CD4+ lymphocytes of relapsing-remitting multiple sclerosis patients. Eur. J. Immunol. 2010, 40, 888–898. [Google Scholar] [CrossRef]
- Tang, J.; Ahmad, A.; Sarkar, F.H. The role of microRNAs in breast cancer migration, invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 13414–13437. [Google Scholar] [CrossRef]
- Aggarwal, T.; Wadhwa, R.; Gupta, R.; Paudel, K.R.; Collet, T.; Chellappan, D.K.; Gupta, G.; Perumalsamy, H.; Mehta, M.; Satija, S.; et al. MicroRNAs as Biomarker for Breast Cancer. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 1597–1610. [Google Scholar] [CrossRef] [PubMed]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef]
- Lei, M.; Zheng, G.; Ning, Q.; Zheng, J.; Dong, D. Translation and functional roles of circular RNAs in human cancer. Mol. Cancer 2020, 19, 30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.D.; Jiang, L.H.; Sun, D.W.; Hou, J.C.; Ji, Z.L. CircRNA: A novel type of biomarker for cancer. Breast Cancer 2018, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vannini, I.; Fanini, F.; Fabbri, M. Emerging roles of microRNAs in cancer. Curr. Opin. Genet. Dev. 2018, 48, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Yu, S.; Chen, Z.; Meng, Z.; Huang, S.; Wang, P. The emerging role of circRNAs and their clinical significance in human cancers. Biochim. Biophys. Acta. Rev. Cancer 2018, 1870, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Wang, J.; Guo, Z.; Li, M.; Li, M.; Liu, S.; Liu, H.; Li, W.; Yin, X.; Tao, J.; et al. Emerging function and potential diagnostic value of circular RNAs in cancer. Mol. Cancer 2018, 17, 123. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Nishikawa, R.; Kato, M.; Okato, A.; Arai, T.; Kojima, S.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of HMGB3 by antitumor miR-205-5p inhibits cancer cell aggressiveness and is involved in prostate cancer pathogenesis. J. Hum. Genet. 2018, 63, 195–205. [Google Scholar] [CrossRef]
- Elgamal, O.A.; Park, J.K.; Gusev, Y.; Azevedo-Pouly, A.C.; Jiang, J.; Roopra, A.; Schmittgen, T.D. Tumor suppressive function of mir-205 in breast cancer is linked to HMGB3 regulation. PLoS ONE 2013, 8, e76402. [Google Scholar] [CrossRef]
- Su, Z.; Wang, C.; Chang, D.; Zhu, X.; Sai, C.; Pei, J. Limonin attenuates the stemness of breast cancer cells via suppressing MIR216A methylation. Biomed. Pharmacother.=Biomed. Pharmacother. 2019, 112, 108699. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Zhang, C.; Li, X.; Yu, J. MiR-216a-3p inhibits colorectal cancer cell proliferation through direct targeting COX-2 and ALOX5. J. Cell. Biochem. 2018, 119, 1755–1766. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Lu, H.N.; Chen, X.; Jiang, X.F.; Yang, Y.; Feng, J. MiR-216a-3p promotes differentiation of BMMSCs into ACE II cells via Wnt/beta-catenin pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7849–7857. [Google Scholar] [CrossRef] [PubMed]
- Felix, T.F.; Lopez Lapa, R.M.; de Carvalho, M.; Bertoni, N.; Tokar, T.; Oliveira, R.A.; MA, M.R.; Hasimoto, C.N.; Oliveira, W.K.; Pelafsky, L.; et al. MicroRNA modulated networks of adaptive and innate immune response in pancreatic ductal adenocarcinoma. PLoS ONE 2019, 14, e0217421. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.K.; Xie, X.N.; Song, X.H.; Su, T.; Chang, X.L.; Xu, M.; Liang, B.; Huang, D.Y. Upregulation of miR-200b Inhibits Hepatocellular Carcinoma Cell Proliferation and Migration by Targeting HMGB3 Protein. Technol. Cancer Res. Treat. 2018, 17, 1533033818806475. [Google Scholar] [CrossRef] [PubMed]
- Maciotta, S.; Meregalli, M.; Cassinelli, L.; Parolini, D.; Farini, A.; Fraro, G.D.; Gandolfi, F.; Forcato, M.; Ferrari, S.; Gabellini, D.; et al. Hmgb3 is regulated by microRNA-206 during muscle regeneration. PLoS ONE 2012, 7, e43464. [Google Scholar] [CrossRef] [PubMed]
- Minemura, C.; Asai, S.; Koma, A.; Kikkawa, N.; Kato, M.; Kasamatsu, A.; Uzawa, K.; Hanazawa, T.; Seki, N. Identification of Antitumor miR-30e-5p Controlled Genes; Diagnostic and Prognostic Biomarkers for Head and Neck Squamous Cell Carcinoma. Genes 2022, 13, 1225. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Pan, J.; Han, X.; Chen, W. Downregulation of microRNA-532-5p promotes the proliferation and invasion of bladder cancer cells through promotion of HMGB3/Wnt/beta-catenin signaling. Chem.-Biol. Interact. 2019, 300, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Li, H.L.; Song, Y.H.; Du, Z.P.; Hu, Y.H.; Wang, Z.X.; Chen, X.; Lu, X.M.; Chen, Y.X.; Duan, Y.Q.; Zhu, X.D. Overexpression of MiR-633 Suppresses the Tumorigenicity of Gastric Cancer Cells and Induces Apoptosis by Targeting MAPK1. Curr. Med. Sci. 2022, 42, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Hou, X.; Lin, B. MicroRNA-758 inhibits cervical cancer cell proliferation and metastasis by targeting HMGB3 through the WNT/beta-catenin signaling pathway. Oncol. Lett. 2019, 18, 1786–1792. [Google Scholar] [CrossRef]
- Sun, C.X.; Zhu, F.; Qi, L. Demethylated miR-216a Regulates High Mobility Group Box 3 Promoting Growth of Esophageal Cancer Cells Through Wnt/beta-Catenin Pathway. Front. Oncol. 2021, 11, 622073. [Google Scholar] [CrossRef]
- Zhou, G.H.; Lu, Y.Y.; Xie, J.L.; Gao, Z.K.; Wu, X.B.; Yao, W.S.; Gu, W.G. Overexpression of miR-758 inhibited proliferation, migration, invasion, and promoted apoptosis of non-small cell lung cancer cells by negatively regulating HMGB. Biosci. Rep. 2019, 39, BSR20180855. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Y.; Zhang, T.; Feng, C.; Liu, Y. High-mobility group box 3 (HMGB3) silencing inhibits non-small cell lung cancer development through regulating Wnt/beta-catenin pathway. Biol. Chem. 2020, 401, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Wang, B.; Feng, G.; Duan, L.; Yuan, S.; Jia, W.; Liu, Y. Knockdown of high mobility group box 3 impairs cell viability and colony formation but increases apoptosis in A549 human non-small cell lung cancer cells. Oncol. Lett. 2019, 17, 2937–2945. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Yadav, P.; Jain, R.P.; Bera, A.K.; Karunagaran, D. miR-142-3p simultaneously targets HMGA1, HMGA2, HMGB1, and HMGB3 and inhibits tumorigenic properties and in-vivo metastatic potential of human cervical cancer cells. Life Sci. 2022, 291, 120268. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Yadav, P.; Sundaram, S.; Venkatraman, G.; Bera, A.K.; Karunagaran, D. HMGB3 inhibition by miR-142-3p/sh-RNA modulates autophagy and induces apoptosis via ROS accumulation and mitochondrial dysfunction and reduces the tumorigenic potential of human breast cancer cells. Life Sci. 2022, 304, 120727. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Liu, B.; Wu, J.L.; Zhang, R.F.; Duan, L.; He, W.S.; Zhang, C.M. Prognostic value of HMGB3 expression in patients with non-small cell lung cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2013, 34, 2599–2603. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Pei, L.; Zhang, D.; Xu, F.; Zhou, E.; Chen, X. HDAC3 increases HMGB3 expression to facilitate the immune escape of breast cancer cells via down-regulating microRNA-130a-3p. Int. J. Biochem. Cell Biol. 2021, 135, 105967. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, H.; Feng, W.; Huang, S.; An, J.; Qiu, Y.; Wu, K. Long non-coding RNA HOTTIP promotes hypoxia-induced glycolysis through targeting miR-615-3p/HMGB3 axis in non-small cell lung cancer cells. Eur. J. Pharmacol. 2019, 862, 172615. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sheng, Z.; Cai, Y. Effects of microRNA-513b on cell proliferation, apoptosis, invasion, and migration by targeting HMGB3 through regulation of mTOR signaling pathway in non-small-cell lung cancer. J. Cell. Physiol. 2019, 234, 10934–10941. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, G.; Wang, F.; Gao, F.; Luo, H.; Wang, Y.; Du, Y.; Chen, X.; Xue, C.; Dong, Z.; et al. Upregulation of miR-513b inhibits cell proliferation, migration, and promotes apoptosis by targeting high mobility group-box 3 protein in gastric cancer. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2014, 35, 11081–11089. [Google Scholar] [CrossRef]
- Liu, D.; Wang, Y.; Zhao, Y.; Gu, X. LncRNA SNHG5 promotes nasopharyngeal carcinoma progression by regulating miR-1179/HMGB3 axis. BMC Cancer 2020, 20, 178. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, H.; Chen, W.; Huang, X.; Fan, R.; Xu, M.; Zou, L. CircRUNX1 drives the malignant phenotypes of lung adenocarcinoma through mediating the miR-5195-3p/HMGB3 network. Gen. Thorac. Cardiovasc. Surg. 2023, 72, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Yang, Y.; Cai, J.; Cui, K.; Li, R.X.; Wang, H.; Shang, X.; Wei, D. MiR-30a-5p Suppresses Tumor Metastasis of Human Colorectal Cancer by Targeting ITGB3. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 39, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, X.; He, Y.; Ge, H. Systematic analysis of expression profiles of HMGB family members for prognostic application in non-small cell lung cancer. Front. Mol. Biosci. 2022, 9, 844618. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Lv, M.; Ji, X.; Xue, L.; Rui, C.; Yin, L.; Ding, H.; Miao, Z. Down-regulated expressed protein HMGB3 inhibits proliferation and migration, promotes apoptosis in the placentas of fetal growth restriction. Int. J. Biochem. Cell Biol. 2019, 107, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xin, K.; Leng, C.; Ge, J. Down-regulation of SNHG16 alleviates the acute lung injury in sepsis rats through miR-128-3p/HMGB3 axis. BMC Pulm. Med. 2021, 21, 191. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Pan, J.; Jin, Y.; Li, M.; Chen, M. MiR-195-5p Inhibits Proliferation and Induces Apoptosis of Non-Small Cell Lung Cancer Cells by Targeting CEP55. OncoTargets Ther. 2019, 12, 11465–11474. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ren, Y.; Liu, R.; Ma, J.; Shi, Y.; Zhang, L.; Bu, R. miR-195-5p Suppresses the Proliferation, Migration, and Invasion of Oral Squamous Cell Carcinoma by Targeting TRIM14. BioMed Res. Int. 2017, 2017, 7378148. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Lv, S.G.; Wu, M.J.; Shen, X.L.; Tu, W.; Ye, M.H.; Zhu, X.G. Circ_CLIP2 promotes glioma progression through targeting the miR-195-5p/HMGB3 axis. J. Neuro-Oncol. 2021, 154, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, C.; Zhao, Z. CircSLCO3A1 depletion ameliorates lipopolysaccharide-induced inflammation and apoptosis of human pulmonary alveolar epithelial cells through the miR-424-5p/HMGB3 pathway. Mol. Cell. Toxicol. 2023, 20, 187–198. [Google Scholar] [CrossRef]
- Zhang, Z.; Chang, Y.; Zhang, J.; Lu, Y.; Zheng, L.; Hu, Y.; Zhang, F.; Li, X.; Zhang, W.; Li, X. HMGB3 promotes growth and migration in colorectal cancer by regulating WNT/beta-catenin pathway. PLoS ONE 2017, 12, e0179741. [Google Scholar] [CrossRef]
- Yuan, H.; Chen, B.; Chai, R.; Gong, W.; Wan, Z.; Zheng, B.; Hu, X.; Guo, Y.; Gao, S.; Dai, Q.; et al. Loss of exosomal micro-RNA-200b-3p from hypoxia cancer-associated fibroblasts reduces sensitivity to 5-flourouracil in colorectal cancer through targeting high-mobility group box 3. Front. Oncol. 2022, 12, 920131. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Yao, Z.; Liu, C.; Fu, S.; Ren, B.; Liu, Y.; Ma, L.; Wei, J.; Sun, D. A novel lnc-LAMC2-1:1 SNP promotes colon adenocarcinoma progression by targeting miR-216a-3p/HMGB3. Heliyon 2022, 8, e12342. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, J.; Zheng, Y.; Ma, C.; Liu, X.E.; Sun, X. miR-216a-3p Inhibits the Proliferation, Migration, and Invasion of Human Gastric Cancer Cells via Targeting RUNX1 and Activating the NF-kappaB Signaling Pathway. Oncol. Res. 2018, 26, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Jiang, Z.; Li, H.; Peng, J.; Chen, X.; Tang, M. MiR-93/HMGB3 regulatory axis exerts tumor suppressive effects in colorectal carcinoma cells. Exp. Mol. Pathol. 2021, 120, 104635. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Chang, J.; Zhang, N.; Wu, S.; Liu, H.; Yu, J. MicroRNA-429 acts as a tumor suppressor in colorectal cancer by targeting high mobility group box 3. Oncol. Lett. 2021, 21, 250. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhang, X.; Bai, W.; Wang, J.; Jiang, B. Circ-IGF1R Affects the Progression of Colorectal Cancer by Activating the miR-362-5p/HMGB3-Mediated Wnt/beta-Catenin Signal Pathway. Biochem. Genet. 2023, 61, 1210–1229. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xiang, W.; Chen, W.; Sun, Y.; Qin, F.; Wei, J.; Yuan, L.; Zheng, L.; Li, S. Circ-IGF1R inhibits cell invasion and migration in non-small cell lung cancer. Thorac. Cancer 2020, 11, 875–887. [Google Scholar] [CrossRef] [PubMed]
- Ke, Y.; Mai, J.; Liu, Z.; Xu, Y.; Zhao, C.; Wang, B. Interfering HMGB3 release from cancer-associated fibroblasts by miR-200b represses chemoresistance and epithelial-mesenchymal transition of gastric cancer cells. J. Gastrointest. Oncol. 2022, 13, 2197–2218. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Li, X. HMGB3 promotes the proliferation and metastasis of glioblastoma and is negatively regulated by miR-200b-3p and miR-200c-3p. Cell Biochem. Funct. 2018, 36, 357–365. [Google Scholar] [CrossRef]
- Song, X.; Wang, H.; Wu, J.; Sun, Y. Long Noncoding RNA SOX2-OT Knockdown Inhibits Proliferation and Metastasis of Prostate Cancer Cells Through Modulating the miR-452-5p/HMGB3 Axis and Inactivating Wnt/beta-Catenin Pathway. Cancer Biother. Radiopharm. 2020, 35, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Staal, F.J.; de Ridder, D.; Szczepanski, T.; Schonewille, T.; van der Linden, E.C.; van Wering, E.R.; van der Velden, V.H.; van Dongen, J.J. Genome-wide expression analysis of paired diagnosis-relapse samples in ALL indicates involvement of pathways related to DNA replication, cell cycle and DNA repair, independent of immune phenotype. Leukemia 2010, 24, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Somervaille, T.C.; Matheny, C.J.; Spencer, G.J.; Iwasaki, M.; Rinn, J.L.; Witten, D.M.; Chang, H.Y.; Shurtleff, S.A.; Downing, J.R.; Cleary, M.L. Hierarchical maintenance of MLL myeloid leukemia stem cells employs a transcriptional program shared with embryonic rather than adult stem cells. Cell Stem Cell 2009, 4, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Clara, J.A.; Monge, C.; Yang, Y.; Takebe, N. Targeting signalling pathways and the immune microenvironment of cancer stem cells—A clinical update. Nat. Rev. Clin. Oncol. 2020, 17, 204–232. [Google Scholar] [CrossRef] [PubMed]
- Calogero, S.; Grassi, F.; Aguzzi, A.; Voigtlander, T.; Ferrier, P.; Ferrari, S.; Bianchi, M.E. The lack of chromosomal protein Hmg1 does not disrupt cell growth but causes lethal hypoglycaemia in newborn mice. Nat. Genet. 1999, 22, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Ronfani, L.; Ferraguti, M.; Croci, L.; Ovitt, C.E.; Scholer, H.R.; Consalez, G.G.; Bianchi, M.E. Reduced fertility and spermatogenesis defects in mice lacking chromosomal protein Hmgb2. Development 2001, 128, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- The human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 10 April 2024).
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. MCP 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Nemeth, M.J.; Cline, A.P.; Anderson, S.M.; Garrett-Beal, L.J.; Bodine, D.M. Hmgb3 deficiency deregulates proliferation and differentiation of common lymphoid and myeloid progenitors. Blood 2005, 105, 627–634. [Google Scholar] [CrossRef]
- Zhong, X.; Zhang, S.; Zhang, Y.; Jiang, Z.; Li, Y.; Chang, J.; Niu, J.; Shi, Y. HMGB3 is Associated With an Unfavorable Prognosis of Neuroblastoma and Promotes Tumor Progression by Mediating TPX2. Front. Cell Dev. Biol. 2021, 9, 769547. [Google Scholar] [CrossRef]
- Zhang, K.; Liu, D.; Zhao, J.; Shi, S.; He, X.; Da, P.; You, Y.; You, B. Nuclear exosome HMGB3 secreted by nasopharyngeal carcinoma cells promotes tumour metastasis by inducing angiogenesis. Cell Death Dis. 2021, 12, 554. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Ronfani, L.; Bianchi, M.E. Regulated expression and subcellular localization of HMGB1, a chromatin protein with a cytokine function. J. Intern. Med. 2004, 255, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Chikhirzhina, E.; Polyanichko, A.; Skvortsov, A.; Kostyleva, E.; Houssier, C.; Vorob’ev, V. HMG1 domains: The victims of the circumstances. Mol. Biol. 2002, 36, 412–418. [Google Scholar] [CrossRef]
- Polyanichko, A.M.; Chikhirzhina, E.V.; Skvortsov, A.N.; Kostyleva, E.I.; Colson, P.; Houssier, C.; Vorob’ev, V.I. The HMG1 ta(i)le. J. Biomol. Struct. Dyn. 2002, 19, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Chikhirzhina, E.; Polyanichko, A.; Leonenko, Z.; Wieser, H.; Vorob’Ev, V. C-terminal domain of nonhistone protein HMGB1 as a modulator of HMGB1–DNA structural interactions. Spectroscopy 2010, 24, 361–366. [Google Scholar] [CrossRef]
- Gao, J.; Zou, Z.; Gao, J.; Zhang, H.; Lin, Z.; Zhang, Y.; Luo, X.; Liu, C.; Xie, J.; Cai, C. Increased expression of HMGB3: A novel independent prognostic marker of worse outcome in patients with esophageal squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 345–352. [Google Scholar] [PubMed]
- Wu, H.; Feng, H.; Miao, X.; Ma, J.; Liu, C.; Zhang, L.; Yang, L. Construction and validation of a prognostic model based on 11 lymph node metastasis-related genes for overall survival in endometrial cancer. Cancer Med. 2022, 11, 4641–4655. [Google Scholar] [CrossRef]
- Cui, Q.; Zhao, J.; Deng, Q.; Liu, Y.; Fu, C.; Wu, Q.; Sun, Q.; Weng, H.; Dai, H.; Yang, W. HMGB3 is a potential diagnostic marker for early cervical lesion screening. Genes Dis. 2023, 10, 2202–2205. [Google Scholar] [CrossRef]











| Protein Name | Description | |
|---|---|---|
| Old Nomenclature (before 2001) | Current Nomenclature (after 2001) | |
| HMG-1 | HMGB1 | Proteins contain structural and functional motifs, known as HMG-box. HMG-Box binds DNA in the minor groove, inducing DNA that bends toward the major groove. The proteins containing HMG-box preferentially bind DNA regions with various structural abnormalities. |
| HMG-2 | HMGB2 | |
| HMG-2a (chicken) | HMGB3 | |
| ssHMG-2b (chicken) | HMGB2 | |
| HMG-3 | - | |
| HMG-4 | HMGB3 | |
| (Unknown until 2008) | HMGB4 | |
| HMG-14 | HMGN1 | The proteins contain structural and functional motifs, known as nucleosomal-binding domain (NBD). Proteins specifically recognize the overall structure of the nucleosome. |
| HMG-17 | HMGN2 | |
| HMG-I | HMGA1a | The functional motif of the HMGA family is called the “AT-hook”. ATH domains bind AT-rich DNA regions with varying affinities but very little DNA sequence specificity. |
| HMG-Y | HMGA1b | |
| Tissue | RNA/Protein Expression | ||
|---|---|---|---|
| HMGB3 | HMGB2 | HMGB1 | |
| Female tissues | high/medium | low/medium | medium/high |
| Endocrine tissues | high/– | very low/medium | medium/high |
| Liver and gallbladder | high/– | very low/low | medium/high |
| Male tissues | medium/low | low/high | medium/high |
| Gastrointestinal tract | medium/– | low/high | medium/high |
| Muscle tissues | medium/– | very low/medium | medium/high |
| Bone marrow and lymphoid tissues | medium/– | high/high | high/high |
| Respiratory system | low/high | very low/medium | medium/high |
| Kidney and urinary bladder | low/low | very low/medium | medium/high |
| Brain | low/– | very low/low | medium/high |
| Proximal digestive tract | low/– | very low/medium | medium/high |
| Skin | low/– | very low/high | low/high |
| Pancreas | very low/– | very low/low | low/high |
| Connective and soft tissue | very low/– | very low/low | medium/high |
| Eye | very low/– | low/– | low/– |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chikhirzhina, E.; Tsimokha, A.; Tomilin, A.N.; Polyanichko, A. Structure and Functions of HMGB3 Protein. Int. J. Mol. Sci. 2024, 25, 7656. https://doi.org/10.3390/ijms25147656
Chikhirzhina E, Tsimokha A, Tomilin AN, Polyanichko A. Structure and Functions of HMGB3 Protein. International Journal of Molecular Sciences. 2024; 25(14):7656. https://doi.org/10.3390/ijms25147656
Chicago/Turabian StyleChikhirzhina, Elena, Anna Tsimokha, Alexey N. Tomilin, and Alexander Polyanichko. 2024. "Structure and Functions of HMGB3 Protein" International Journal of Molecular Sciences 25, no. 14: 7656. https://doi.org/10.3390/ijms25147656