Abstract
The fructose-1,6-bisphosphate aldolase (FBA) gene family exists in higher plants, with the genes of this family playing significant roles in plant growth and development, as well as response to abiotic stresses. However, systematic reports on the FBA gene family and its functions in cucumber are lacking. In this study, we identified five cucumber FBA genes, named CsFBA1-5, that are distributed randomly across chromosomes. Phylogenetic analyses involving these cucumber FBAs, alongside eight Arabidopsis FBA proteins and eight tomato FBA proteins, were conducted to assess their homology. The CsFBAs were grouped into two clades. We also analyzed the physicochemical properties, motif composition, and gene structure of the cucumber FBAs. This analysis highlighted differences in the physicochemical properties and revealed highly conserved domains within the CsFBA family. Additionally, to explore the evolutionary relationships of the CsFBA family further, we constructed comparative syntenic maps with Arabidopsis and tomato, which showed high homology but only one segmental duplication event within the cucumber genome. Expression profiles indicated that the CsFBA gene family is responsive to various abiotic stresses, including low temperature, heat, and salt. Taken together, the results of this study provide a theoretical foundation for understanding the evolution of and future research into the functional characterization of cucumber FBA genes during plant growth and development.
1. Introduction
Cucumber (Cucumis sativus L.) is among the most crucial vegetable crops globally, having a tropical origin. This crop is sensitive to cold and, therefore requires substantial energy for greenhouse production in cooler seasons [1]. Cucumbers are vulnerable to chilling temperatures, a major environmental stress that substantially affects their growth and development, often resulting in substantial economic losses.
Throughout their long evolutionary history, plants have developed complex regulatory networks to detect stress signals and adapt to adverse conditions through interaction of polysaccharides and water molecules [2] or by regulating metabolic processes. Chloroplasts, which are essential for photosynthesis in green plants, enable plants to respond to environmental stresses and adapt to adverse environmental conditions by regulating metabolism pathways [3,4]. They are the sites of photosynthesis and oxygen release processes that produce energy to support plant growth and yield. Chloroplasts also synthesize essential compounds such as amino acids, phytohormones, nucleotides, vitamins, lipids, and secondary metabolites in response to environmental stresses [5]. They act as environmental sensors, relaying signals to adjust the metabolic processes and gene expressions in plants to cope with stresses [6,7,8]. Studies have shown that chloroplasts enhance photosynthesis and plant resilience under adverse environmental conditions [9]. Under extreme conditions, such as low temperatures, heat, and salinity, chloroplasts produce reactive oxygen species (ROS) that can damage the bio-membrane system and further deteriorate plant health and viability. Excessive ROS can also hinder carbon fixation by deactivating enzymes in the Calvin–Benson cycle, thereby reducing photosynthesis, which is crucial for carbohydrate synthesis. However, ROS can also be served as a feedback signal to manage stresses [9]. In response to low temperatures, the structure and metabolism of chloroplasts undergo alteration, leading to increases in the plant’s tolerance to these conditions. Adaptation to chilling stress involves regulating the activities of dark-reaction-related enzymes in chloroplasts, such as fructose-1,6-diphosphatase (FBPase) and isoheptanone-1,7-diphosphatase (SBPase) [10]. In addition, researches have confirmed that various plant hormones, such as auxin (IAA) [11], abscisic acid (ABA) [12,13,14], salicylic acid (SA) [15,16], jasmonic acid (JA) [17], and ethylene (ET) [18,19,20], as well as related genes and transcription factors, play crucial roles in regulating plants’ chilling tolerance. The transcription factor dehydration-responsive element binding factor 1 (DREB1)/C-repeat binding factor (CBF/DREB1) is particularly known for its role in cold acclimation. The overexpression of this factor in Arabidopsis and cucumber activates the downstream cold-responsive (COR) genes to respond to this stress [21,22,23,24,25,26,27].
Fructose-1,6-bisphosphate aldolase (EC 4.1.2.13, FBA) is a critical enzyme associated with both the glycolysis/gluconeogenesis pathway and the Calvin cycle. It can be divided into class I and class II FBAs. Higher plants contain mainly class I FBAs, which are present in the plastids and cytoplasm and play crucial roles in enzymatic reactions and in regulating stress tolerance [28,29]. Extensive research on FBA genes in various plant species has enhanced our understanding of this gene family. Different members of the FBA family have been identified and characterized in numerous plant species, such as Arabidopsis [28], tomato [29], rice [30], maize [31], spinach [32], soybean [33], potato [34], oat [35], tobacco [36], purslane [37], wheat [38], moso bamboo [39], and tea tree oil [40]. In Arabidopsis and tomato, the FBA gene family consists of eight members, named FBA1-8. However, in rice, the FBA family consists of only seven members, named ALD1-6 and ALDY. These variations suggest that gene families have expanded or contracted during evolution. Studies have also explored the diverse functions of FBA genes in response to various biotic and abiotic stresses. Numerous studies have indicated that the FBA gene can respond to various stresses, such as salinity [36,37], drought [41], and temperature [35,42,43].
Although FBA genes have been extensively studied in many species, little is known regarding this gene family in the cucumber (C. sativus L.) genome. However, the recent publication of the cucumber genome sequence (available at http://cucurbitgenomics.org/; accessed on 8 April 2024) provides an opportunity to explore the FBA gene family in cucumber, as well as its evolutionary history. To address this research gap and to predict the phenotype, which is one of the central tasks of genome research [44], we used various bioinformatics methods to identify FBA genes in the cucumber genome and analyzed their expression levels under different abiotic stresses because the network-based method achieves more accurate predictions and better interpretability [45]. In addition, we identified some candidate genes for further functional research in the field.
In this study, we identified five CsFBA genes and divided them into two subgroups. We conducted phylogenetic and synteny analyses, assessed gene structures, and examined conserved motifs. In addition, we analyzed the expression profiles of CsFBA genes under low-temperature, heat, and salt conditions to understand how these genes respond to these abiotic stresses.
2. Results
2.1. Genome-Wide Identification of CsFBA Genes in Cucumber
Five members of the FBA gene family in the cucumber genome were identified through BlastP analysis, all previously annotated as fructose phosphor aldolase. These members were designated as CsFBA1 to CsFBA5 based on their chromosomal locations in the official database. Specific details of these genes, including sequence ID, amino acid count (length), protein molecular weight (MW), theoretical isoelectric point (pI), instability index, aliphatic index, and grand average of hydropathicity, are presented in Table 1. Among the identified members, CsFBA1 had the most amino acids (397), whereas CsFBA5 had the fewest amino acids (357). The MW ranged from 37.97 kDa (CsFBA5) to 42.89 kDa (CsFBA1). The pI values ranged from 6.07 for CsFBA5 to 8.69 for CsFBA3, with three of these members having acidic properties. The grand average of hydropathicity indicates that all five members were hydrophilic proteins.

Table 1.
Summary information of FBA gene family in cucumber.
2.2. Phylogenetic Analysis of the FBA Family from Arabidopsis, Tomato, and Cucumber Genomes
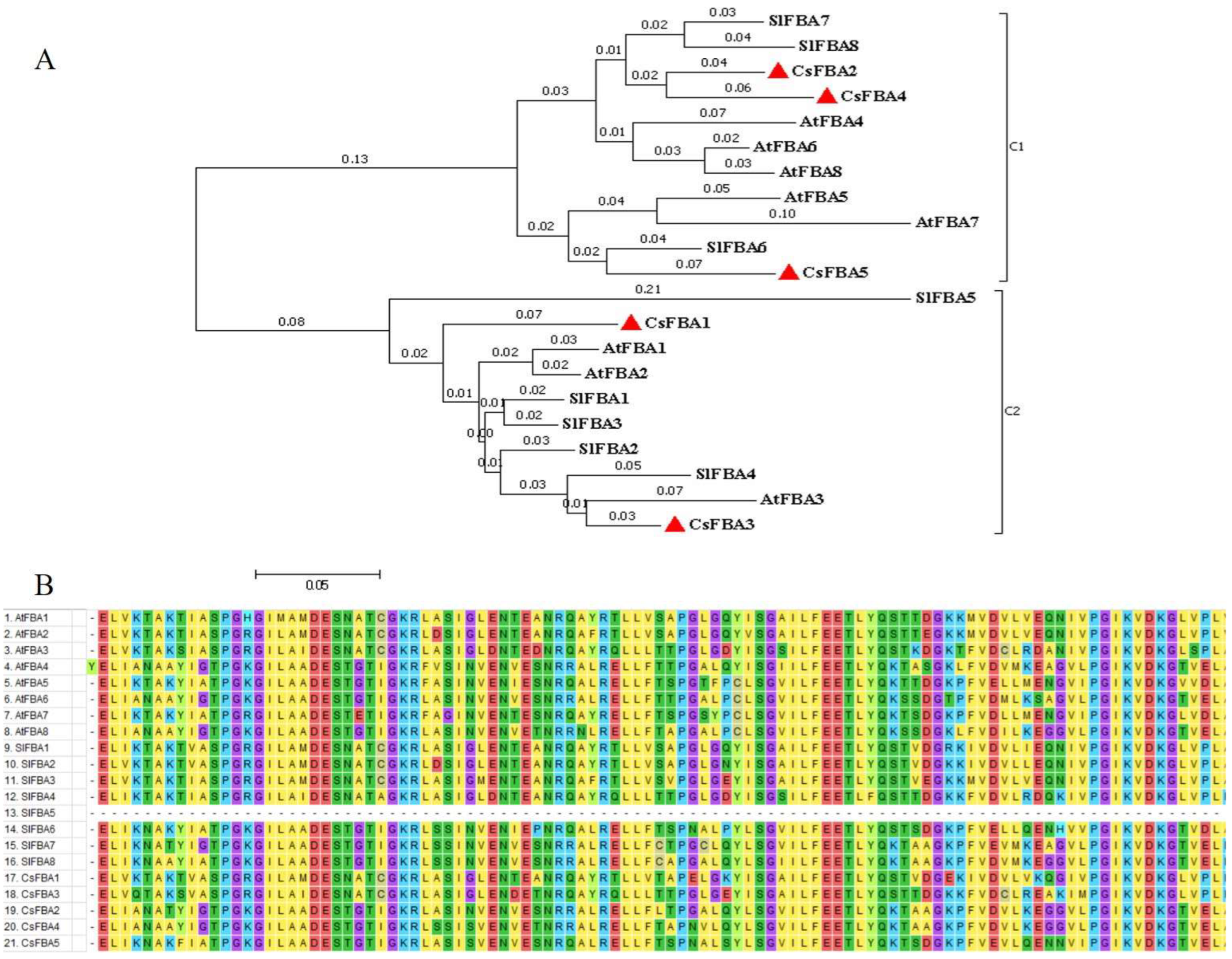
Based on the full-length amino acid sequences of the five cucumber FBA proteins (CsFBAs), along with eight Arabidopsis FBAs (AtFBAs) and eight tomato FBAs (SlFBAs) identified previously, a phylogenetic tree was constructed using MEGA 7.0 software (Figure 1A). The FBA proteins were classified into two clades. Clade 1 comprised CsFBA2, CsFBA4, and CsFBA5, together with AtFBA4-8 and SlFBA6-8, which were predicted to be located in the cytoplasm. Conversely, CsFBA1 and CsFBA3, along with AtFBA1-3 and SlFBA1-5, with their subcellular location predicted to be the chloroplast, fell into clade 2. Furthermore, the phylogenetic tree showed close clustering of CsFBA2 and CsFBA4 with SlFBA7 and CsFBA3 with SlFBA4, suggesting high homology among these proteins and potentially similar gene functions.
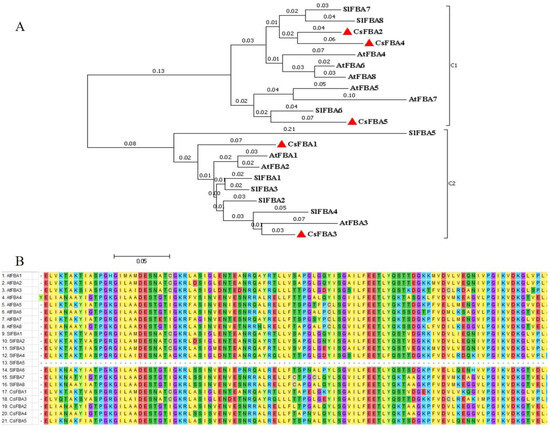
Figure 1.
Evolutionary relationships of FBA family in cucumber, Arabidopsis, and tomato. (A) Phylogenetic tree of the relationship between the FBA proteins of cucumber, Arabidopsis, and tomato. C1 and C2 represent different subfamilies. The evolutionary history was inferred using the neighbor-joining method [46]. The optimal tree with a sum of branch length = 1.60678193 is shown. The tree is drawn to scale, with branch lengths (above the branches) in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the p-distance method [47] and are presented as the number of amino acid differences per site. The analysis involved 21 amino acid sequences. All positions containing gaps and missing data were eliminated, resulting in a final dataset comprising a total of 132 positions. Evolutionary analyses were conducted using MEGA7.0 [48]. (B) Multiple sequence alignment of the glycolysis domains of 21 FBA proteins from cucumber, A. thaliana, and tomato using 100 amino acids on either side of the structural domain.
To further explore the evolutionary relationships and structural domains of CsFBA proteins, multiple sequence alignments of protein sequences from AtFBAs, SlFBAs, and CsFBAs were conducted (Figure 1B). Specifically, 100 amino acid sequences from the N terminal, containing the structural domains, were selected for this analysis. The results indicated that the sequences within the structural domain were highly conserved, with the exception of SlFBA5.
2.3. Gene Structure and Conserved Motif Analysis
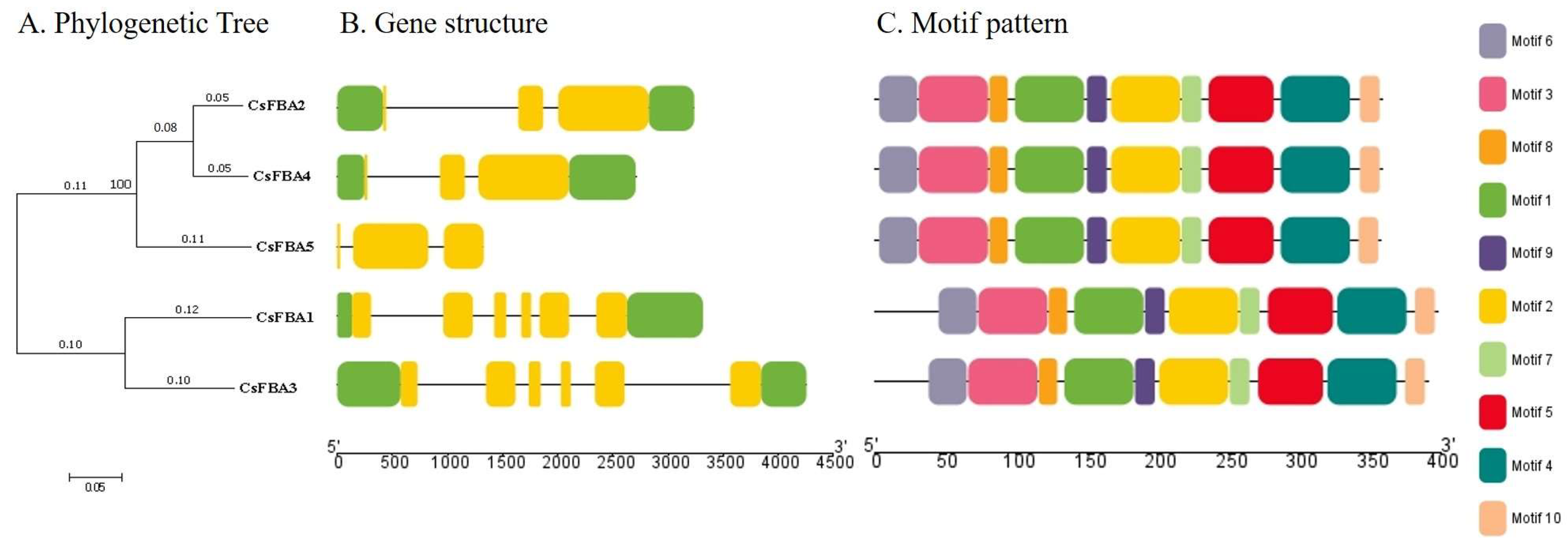
To analyze the structural composition of CsFBA genes, a structural map was created using the cucumber genome sequence. This map included the untranslated regions (UTRs), coding sequences (CDSs), and introns (Figure 2A,B). A comparison of the number and placement of exons and introns across the CsFBA genes revealed variability; CsFBA genes in clade 1 (C1) had three exons each, whereas those in clade 2 (C2) contained six exons each. Notably, CsFBA5 had no UTRs. To further explore the conservation of the CsFBA gene family throughout evolution, we analyzed their conserved motif patterns using TBtools. Ten conserved motifs, designated as motifs 1 to 10 and distinguished by various colors, were identified. The presence of all 10 motifs in each of the five CsFBA proteins suggested a high degree of conservation throughout evolutionary history.
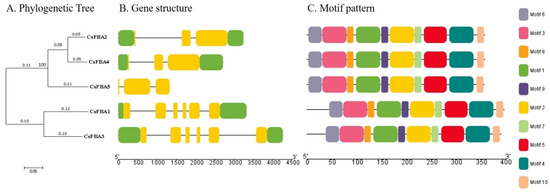
Figure 2.
Phylogenetic tree, gene structure, and motif pattern of CsFBA proteins. (A) The phylogenetic tree was constructed using the full-length sequences of CsFBA proteins with 1000 replicates on each node. (B) Green rectangles, yellow rectangles, and black lines indicate UTRs (non-coding regions), CDSs (coding sequences or exons), and introns, respectively. (C) The amino acid motifs (numbered 1–10) in CsFBA proteins are displayed in ten colored boxes, and black lines indicate amino acid length.
2.4. Chromosomal Distribution and Gene Duplication Analysis of CsFBA Genes
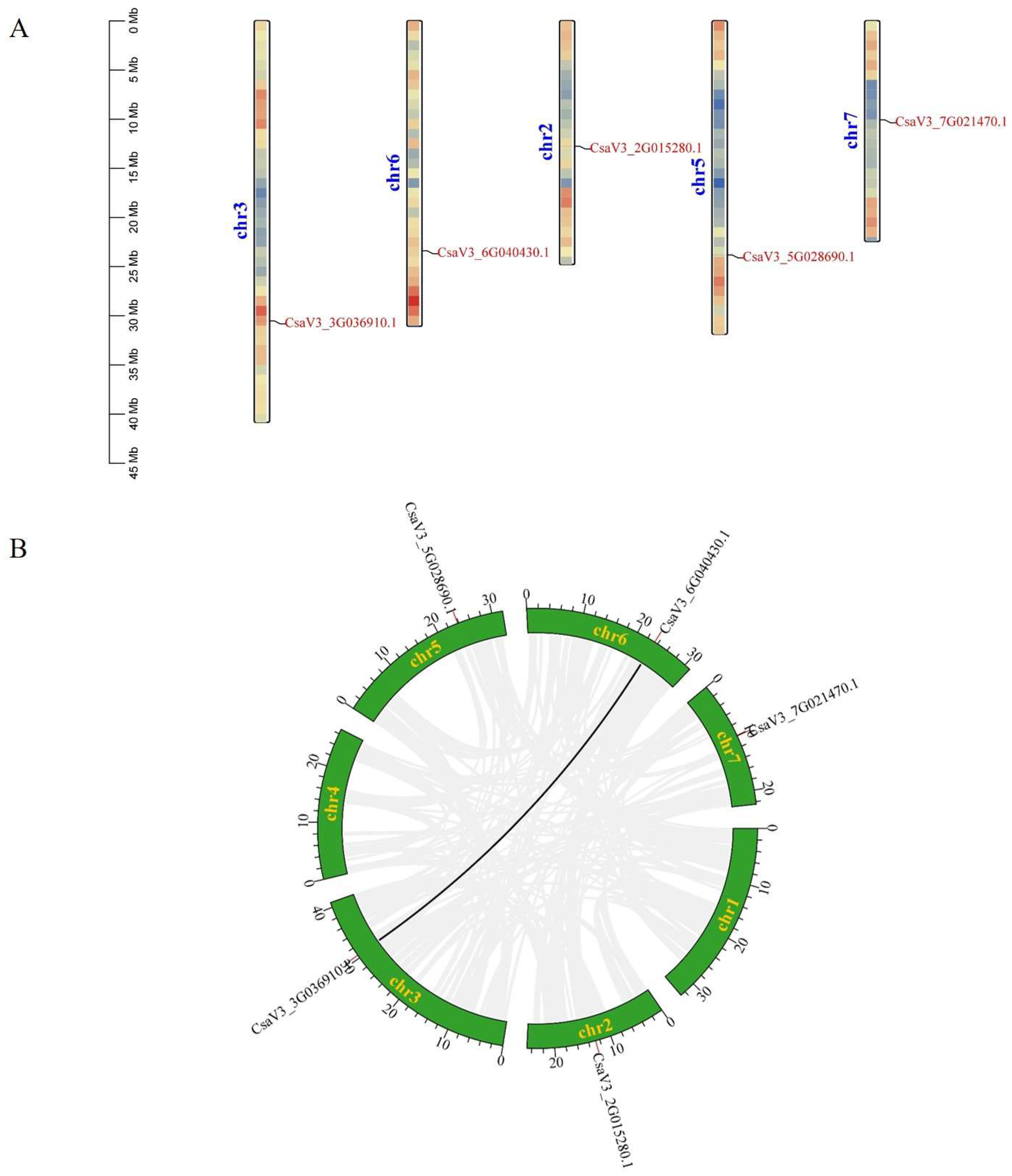
The five CsFBA genes were located on five chromosomes, with one gene per chromosome; CsFBA1 was on chromosome 2, CsFBA2 on chromosome 3, CsFBA3 on chromosome 5, CsFBA4 on chromosome 6, and CsFBA5 on chromosome 7 (Figure 3A). Gene duplication events, which include tandem and segmental duplications, were analyzed to understand the mechanisms underlying the expansion or contraction of the CsFBA gene family. A tandem duplication event is defined as the presence of two or more genes in a chromosomal region within 200 kb [49]. Our analysis revealed no tandem duplication events within the CsFBA family. However, one segmental duplication event was identified, involving a pair of genes (CsFBA2 and CsFBA4) within the cucumber genome (Figure 3B).
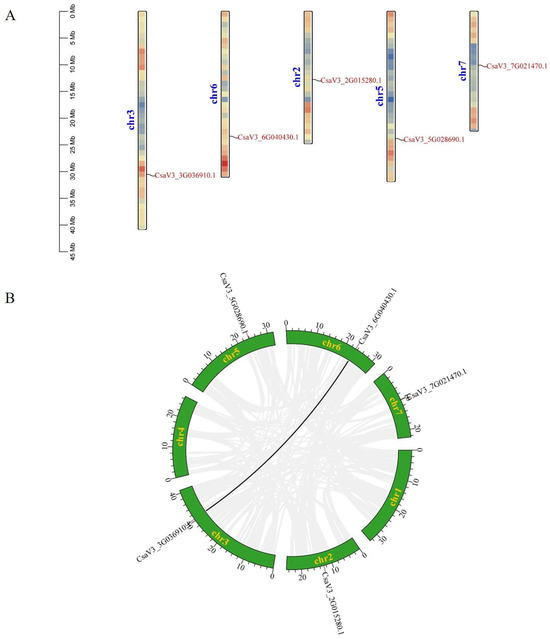
Figure 3.
(A) Chromosomal location of CsFBAs. The colored rectangular bars represent the chromosomes of cucumber, and the 0–45 Mb scale represents chromosome length. (B) Synteny analysis of FBA genes on cucumber chromosomes. Seven cucumber chromosomes are colored in green with their names. The black line in the figure denotes the syntenic region of the cucumber genome.
2.5. Analysis of Collinearity and Evolutionary Relationships between CsFBA and FBA Proteins in Arabidopsis and Tomato Genomes
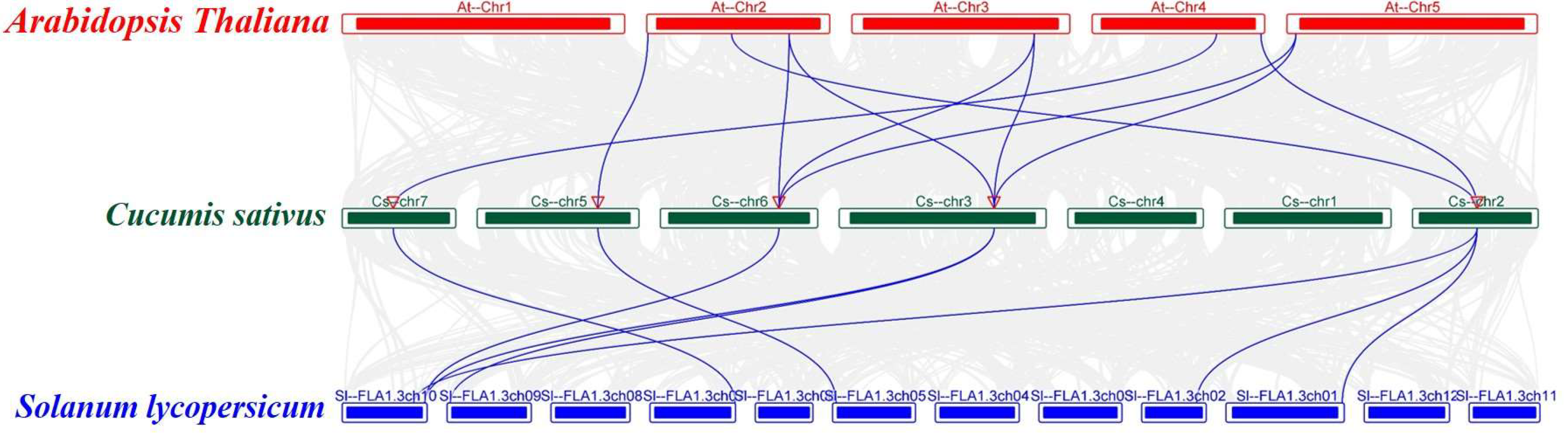
To investigate the evolutionary relationships between the CsFBA gene family in cucumber and FBA genes in other species, a collinear map of cucumber with Arabidopsis and tomato was constructed (Figure 4). This analysis revealed a strong collinear relationship between the CsFBA gene family and the FBA genes of Arabidopsis and tomato. Specifically, the five CsFBA genes showed collinearity with seven AtFBA genes from Arabidopsis and with seven out of the eight SlFBA genes from tomato.
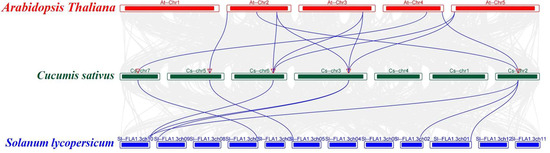
Figure 4.
Collinear analyses of FBA genes between cucumber, Arabidopsis, and tomato. The gray lines between cucumber and other plants represent collinear blocks in wide regions of the genomes, while blue lines show the orthologous relationship of FBA genes.
2.6. Expression Pattern of CsFBAs in Different Conditions
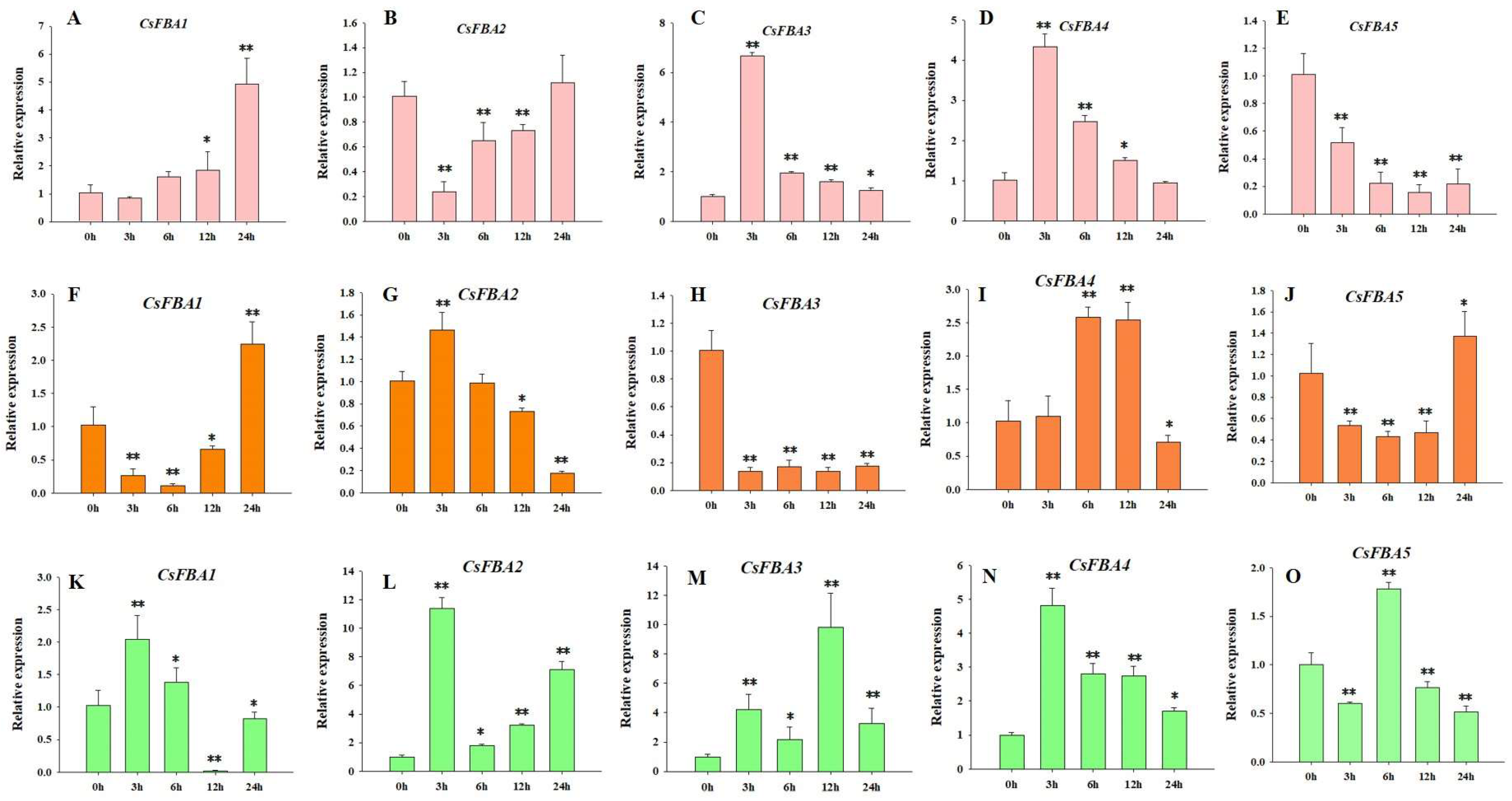
FBA genes play a crucial role in responding to abiotic stresses and affect plant growth and development [29]. To further explore the role of the CsFBA genes in response to various stresses, cucumber seedlings were subjected to low-temperature, heat, and salt stresses. The samples were collected at intervals of 3, 6, 12, and 24 h post treatment, with 0 h serving as the control. The relative expressions of the five CsFBA genes were measured. As shown in Figure 5, all five CsFBA genes responded to low-temperature, heat, and salt treatments. The expression of CsFBA1, CsFBA3, and CsFBA4 was particularly induced by low-temperature stress, with CsFBA3 and CsFBA4 showing a substantial increase within the first 3 h. In contrast, the expression levels of CsFBA2 and CsFBA5 were suppressed under low-temperature conditions. Under heat stress, the expression of CsFBA2 and CsFBA4 increased, whereas the expression levels of CsFBA1, CsFBA3, and CsFBA5 decreased. Moreover, 24 h after heat treatment, the expression levels of CsFBA1 and CsFBA5 recovered to their original levels or even exceeded those at the 0 h mark, suggesting a possible self-regulation mechanism in plants. All five CsFBA genes were also induced at various times after exposure to salt stress, indicating a broad spectrum of stress responsiveness.
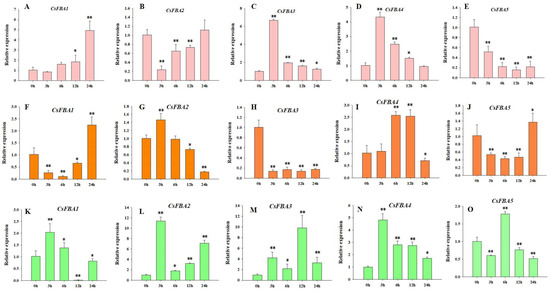
Figure 5.
Expression patterns of the five CsFBA genes in cucumber seedlings after low-temperature treatment (A–E), heat treatment (F–J), and salt treatment (K–O). * Significant differences according to t-test (* p < 0.05, ** p < 0.01).
3. Discussion
Many gene families play essential roles in metabolic and development processes [50]. In recent years, the focus on understanding regulatory mechanisms has led to the identification of many gene families. FBAs are widely distributed in various higher plant species and are essential for regulating plant resistance to biotic and abiotic stresses [49]. To date, information on the FBA gene family has been reported in Arabidopsis [28], tomato [29], and rice genomes [40], with particular attention to the structure, kinetic parameters, and potential therapeutic applications of these genes. However, reports on the FBA gene family in the cucumber genome are lacking, and there has been limited study on the functions of this gene family. Are they regulatory factors in plant growth and tolerance to abiotic stresses? Are they candidate genes for the breeding of new varieties of vegetables with low-temperature tolerance? In this study, we used various bioinformatics methods to identify the FBA gene family in cucumber. Our objectives were to (1) identify the FBA gene family in the cucumber genome, (2) predict the responses of genes to abiotic stresses, and (3) select some candidate CsFBA genes for further functional research on regulating growth or metabolic pathways in plants.
In this study, we identified five CsFBA genes in cucumber, which contains fewer FBA members than Arabidopsis, tomato, and rice. This suggests that the gene family may have undergone contraction during evolution, potentially leading to the loss of some functionalities. We also analyzed the physicochemical properties of these proteins, including amino acid count (length), protein MW, pI, instability index, aliphatic index, and grand average of hydropathicity (Table 1). The variations among the five CsFBA genes indicate divergence in the evolutionary paths due to changing environmental conditions. Phylogenetic and subcellular localization prediction analyses suggested that CsFBA1 and CsFBA3 are homologous with chloroplast/plastid FBAs, such as AtFBA1-3 and SlFBA1-4, whereas CsFBA2, CsFBA4, and CsFBA5 are similar to cytosolic FBAs, such as AtFBA4-8 and SlFBA5-8 (Figure 1A) [28,29]. Chloroplast is the organelle for photosynthesis, and the FBA genes in this clade play a crucial part in both photosynthetic and non-photosynthetic metabolism pathways and, therefore, in regulating development and tolerance to low-temperature stress [51]. A previous study showed that over-expression of SlFBA7 in tomato led to an enhanced net photosynthetic rate and activity of other enzymes in the Calvin cycle. However, a study of mutants in Arabidopsis showed that there is functional redundancy of AtFBA1, AtFBA2, and AtFBA3 in regulating plant growth. Because the fba1 mutants showed no phenotype distinguishable from wild type, while both the fba2 and fba3 mutants showed reduced growth, double mutants fba2/fba3 were lethal [51]. This provides a bright insight in the study of gene family functions. The cytolist-located FBA gene AtFBA6 not only functions in metabolic process but also works as a transcription factor [52]. It has a relatively higher expression level in shoot-apical meristem, root-apical meristem, and the vascular bundle, and the protein can interact with WUS, WOX4, and WOX5 [52]. For protein function studies, SDS-PAGE and in vitro enzymatic synthesis of biochemicals can provide more bright ideas [53]. This differentiation in localization suggests varied functions among the members, with plastid-located proteins CsFBA1 and CsFBA3 likely playing crucial roles in the Calvin cycle, particularly in response to low-temperature stress. The conserved gene structure and motifs across these genes imply functional conservation, with orthologous genes typically sharing similar functions and grouped within the same phylogenetic clades. The collinear relationship of CsFBA genes with those of Arabidopsis and tomato provides insights into the potential functions of cucumber FBA genes. The results of collinearity analysis (Figure 4) align with previous studies suggesting that all AtFBA genes play a role in response to abnormal temperature stresses. Phylogenetic studies suggest that CsFBA2, CsFBA3, and CsFBA4 share a high homology with SlFBA7, further indicating the specific roles of these genes in responding to low-temperature stress, warranting further investigation. In addition, we found that all five CsFBA genes respond to low-temperature, heat, and salt stresses to different degrees. Notably, CsFBA1, CsFBA3, and CsFBA4 were induced by low-temperature and salt stresses (Figure 5), indicating their similar roles in response to low-temperature and salt stresses. However, under heat stress, the expression of CsFBA1, CsFBA3, and CsFBA5 was suppressed, while the remaining genes were induced. These results indicate that these genes regulate stress tolerance, possibly through different mechanisms. The plastid, a key site for photosynthesis and biosynthesis of starch and sugar, plays a significant role. In Arabidopsis, the fba2 mutant displayed growth retardation under short-day conditions, unlike the fba1 mutant, which grew normally, suggesting a more pivotal role of AtFBA2 in regulating plant growth [51]. Similarly, the fba8 mutant exhibited a distinct phenotype compared with the wide type. Based on these observations, we hypothesize that CsFBA genes may be involved in regulating plant growth and development, warranting further explorations.
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Treatments
We used cucumber cultivar ‘Jinyou 35’, a prickly Chinese long cucumber. The seeds were soaked in distilled water for 4–6 h, then sown on moist filter paper, allowing them to germinate in the dark at 28 °C. After three days of germination, the seeds were transferred to nutrition pots filled with culture substrate and placed in a solar greenhouse. Here, they received natural sunlight during the day (with a peak photosynthetic photon flux density of 800–1000 μmol/m²/s) and were maintained at a day/night temperature of 25 °C–31 °C/13 °C–21 °C under a 13 h photoperiod [54]. Once the seedlings developed three true leaves, they were subjected to different stress treatments. For low-temperature and heat stress treatments, the seedlings were moved to growth chambers set at 8 °C/5 °C (day/night) and 42 °C/32 °C (day/night), respectively. The second leaves were harvested 0, 3, 6, 12, and 24 h after initiating treatment; immediately frozen in liquid nitrogen; and stored at −80 °C for subsequent analysis. For salt stress, the seedlings were treated with Okazaki nutrient solution adjusted to a salinity concentration of 150 mmol/L. The second leaves were harvested at different time intervals similar to those in the low-temperature and heat stress treatments, quickly frozen in liquid nitrogen, and stored at −80 °C for analysis.
4.2. Identification of CsFBA Genes in the Cucumber Genome
Arabidopsis FBA protein sequences were used as a query and searched against the cucumber genome database (http://cucurbitgenomics.org/; accessed on 8 April 2024). The GFF3 file of the cucumber genome database was downloaded from the Cucurbit Genomics Database. Sequence analysis was conducted using the Local BlastP program through TBtools. Pfam (http://pfam.janelia.org; accessed on 8 April 2024) and SMART (http://smart.embl-heidel-berg.de/; accessed on 8 April 2024) tools were used to confirm the glycolytic domain (PF00274) and verify the reliability of each candidate CsFBA protein as a cucumber FBA family member.
4.3. Phylogenetic Analysis
Protein sequences for Arabidopsis, tomato, and cucumber were prepared in FASTA format. Multiple sequence alignments were performed using ClustalX 1.8, with default parameters. The phylogenetic tree was constructed using MEGA 7.0 [48]. The neighbor-joining method [46] was used with the following parameters: p-distance substitution model [47], pairwise deletion, and bootstrap (1000 replicates; random seed) [55].
4.4. Gene Structure and Conserved Motif Analysis
For the analysis of the cucumber FBA gene family and multiple sequence comparisons, we performed structural mapping of CsFBA genes and visualized conserved motifs by using TBtools-II v2.096 software, utilizing a previously prepared GFF3 file. The search for maximum conserved motifs was set to 10, with all other parameters left at their default settings.
4.5. Chromosomal Distribution and Synteny Analysis of CsFBA Genes
Genomic FASTA files and genome annotation GFF3 files were prepared to analyze the chromosomal distribution of the genes. These files were then input into the One Step MSCanX program within TBtools-II v2.096. This process highlighted target genes on the visual map, providing a clear representation of their chromosomal locations.
4.6. Total RNA Extraction, Reverse Transcription, and qRT-PCR Analysis
Samples from Section 2.1 (weighing 0.1 g) were thoroughly ground in liquid nitrogen, and total RNA was extracted using TRIzol reagent as per the manufacturer’s instructions. The integrity of RNA was assessed using 1% agarose gel electrophoresis, whereas its purity and concentration were measured using a spectrophotometer. For cDNA synthesis, 1 μg of high-quality total RNA was used. Specific primers for quantitative real-time PCR (qRT-PCR) were designed using Primer Premier software (version 5.0; Premier, Nanaimo BC, Canada). These primers were selected to prevent the amplification of conserved regions and exclusively amplify products ranging from 150 to 300 bp in length. β-actin served as an internal control for the experiments. Quantitative real-time PCR was conducted using PerfecStart Green qPCR Super Mix, following the manufacturer’s protocol. The amplification was conducted on a BIO-RAD CFX96 real-time PCR detection system (Bio-Rad, Hercules, CA, USA). Each sample type was analyzed in triplicate. The relative mRNA expression levels of the target genes were calculated using the 2−∆∆Ct method [56].
4.7. Statistical Analysis
For this study, data entry and statistical analyses were conducted using Microsoft Excel 2010. Statistical differences were determined using DPS 9.01 software, employing Student’s t-test. Bar graphs were generated using Origin 2020. Statistical significance was determined at two levels, namely p < 0.05 and p < 0.01, which are denoted by asterisks (*) and double asterisks (**), respectively, in the figures.
5. Conclusions
Our study systematically identified five CsFBA genes from the whole genome of cucumber. The physicochemical properties of the CsFBA genes are similar, and their structures are conserved. The five CsFBA genes are distributed across five chromosomes, with only one pair of genes derived from segmental duplication. Moreover, the CsFBA gene family responds to temperature and salt stresses, indicating their potential involvement in environmental stress responses. This study not only provides a theoretical basis but also lays the basis for future research into the functions and mechanisms of cucumber FBA genes in plant development and responses to abiotic stresses.
Author Contributions
J.Z., Z.Z. and L.Y. gathered the data and wrote the original manuscript. Y.L. generated all the figures in the revised manuscript. Z.X., Q.L. and B.C. edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the China Agriculture Research System of MOF and MARA (grant number CARS-24-G-03), an earmarked fund for the Modern Agro-industrial Technology Research System of Hebei (grant number HBCT2023100212), and the Hebei Province Youth Science Fund project (grant number C2023204123).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
| FBA | Fructose-1, 6-bisphosphate aldolase |
| ROS | Reactive oxygen species |
| FBPase | Fructose-1,6-diphosphatase |
| SBPase | Isoheptanone-1,7-diphosphatase |
| IAA | Indoleacetic acid |
| ABA | Abscisic acid |
| SA | Salicylic acid |
| JA | Jasmonic acid |
| ET | Ethylene |
| DREB1 | Dehydration-responsive element binding factor 1 |
| CBF | C-repeat binding factor |
| COR | Cold response |
| qRT-PCR | Quantitative real-time PCR |
| MW | Molecular weight |
| pI | Isoelectric point |
References
- Wang, X.; Mi, S.; Miao, H. Transcriptomic responses to chilling reveal potential chilling tolerance mechanisms in cucumber. Int. J. Mol. Sci. 2022, 23, 12834. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Zhang, Y.; Guo, L.; Li, D.; Liu, A.; Bilal, M.; Xie, C.; Yang, R.; Gu, Z.; Jiang, D.; et al. Antifreeze polysaccharides from wheat bran: The structural characterization and antifreeze mechanism. Biomacromolecules 2024, 10, 3c00958. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, A.H.; Li, X.M.; Lu, C.M. The role of chloroplast gene expression in plant responses to environmental stress. Int. J. Mol. Sci. 2020, 21, 6082. [Google Scholar] [CrossRef] [PubMed]
- Littlejohn, G.R.; Breen, S.; Smirnoff, N.; Grant, M. Chloroplast immunity illuminated. New Phytol. 2021, 229, 3088–3107. [Google Scholar] [CrossRef] [PubMed]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Crawford, T.; Lehotai, N.; Strand, Å. The role of retrograde signals during plant stress responses. J. Exp. Bot. 2018, 69, 2783–2795. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.Z.; Meyer, E.H.; Wu, S.; Bock, R. Extensive posttranscriptional regulation of nuclear gene expression by plastid retrograde signals. Plant Physiol. 2019, 180, 2034–2048. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.C.; Burch-Smith, T.M. Chloroplasts as mediators of plant biotic interactions over short and long distances. Curr. Opin. Plant Biol. 2019, 50, 148–155. [Google Scholar] [CrossRef]
- Gan, P.; Liu, F.; Li, R.B.; Wang, S.K.; Luo, J.J. Chloroplasts-beyond energy capture and carbon fixation: Tuning of photosynthesis in response to chilling stress. Int. J. Mol. Sci. 2019, 20, 5046. [Google Scholar] [CrossRef]
- KingstonSmith, A.H.; Harbinson, J.; Williams, J.; Foyer, C.H. Effect of chilling on carbon assimilation, enzyme activation, and photosynthetic electron transport in the absence of photoinhibition in maize leaves. Plant Physiol. 1997, 114, 1039–1046. [Google Scholar] [CrossRef]
- Zhang, X.W.; Liu, F.J.; Zhai, J.; Li, F.D.; Bi, H.G.; Ai, X.Z. Auxin acts as a downstream signaling molecule involved in hydrogen sulfide-induced chilling tolerance in cucumber. Planta 2020, 251, 69. [Google Scholar] [CrossRef]
- Chen, J.X.; Mao, L.C.; Mi, H.B.; Lu, W.J.; Ying, T.J.; Luo, Z.S. Involvement of three annexin genes in the ripening of strawberry fruit regulated by phytohormone and calcium signal transduction. Plant Cell Rep. 2016, 35, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Yang, S.; Wang, Y.; Lu, L.; Sun, M.; He, C.; Wang, J.; Li, Y.; Yu, X.; Li, Q.; et al. Physiological and molecular mechanisms of ABA and CaCl(2) regulating chilling tolerance of cucumber seedlings. Plants 2021, 10, 2746. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Li, F.; Ai, X.; Bi, H. H2O2 participates in ABA regulation of grafting-induced chilling tolerance in cucumber. Plant Cell Rep. 2022, 41, 1115–1130. [Google Scholar] [CrossRef]
- Dong, C.J.; Li, L.; Shang, Q.M.; Liu, X.Y.; Zhang, Z.G. Endogenous salicylic acid accumulation is required for chilling tolerance in cucumber (Cucumis sativus L.) seedlings. Planta 2014, 240, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Feng, Y.Q.; Zhang, X.W.; Zhang, Y.Y.; Bi, H.G.; Ai, X.Z. Salicylic acid is involved in rootstock-scion communication in improving the chilling tolerance of grafted cucumber. Front. Plant Sci. 2021, 12, 693344. [Google Scholar] [CrossRef]
- Qi, C.D.; Dong, D.H.; Li, Y.F.; Wang, X.W.; Guo, L.Q.; Liu, L.; Dong, X.N.; Li, X.S.; Yuan, X.W.; Ren, S.X.; et al. Heat shock-induced cold acclimation in cucumber through CsHSFA1d-activated JA biosynthesis and signaling. Plant J. 2022, 111, 85–102. [Google Scholar] [CrossRef]
- Catalá, R.; López-Cobollo, R.; Castellano, M.M.; Angosto, T.; Alonso, J.M.; Ecker, J.R.; Salinas, J. The Arabidopsis 14-3-3 protein RARE COLD NDUCIBLE 1A links low-temperature response and ethylene biosynthesis to regulate freezing tolerance and cold acclimation. Plant Cell 2014, 26, 3326–3342. [Google Scholar] [CrossRef]
- Dong, Y.F.; Tang, M.J.; Huang, Z.L.; Song, J.N.; Xu, J.; Ahammed, G.J.; Yu, J.Q.; Zhou, Y.H. The miR164a-NAM3 module confers cold tolerance by inducing ethylene production in tomato. Plant J. 2022, 111, 440–456. [Google Scholar] [CrossRef]
- Shi, Y.T.; Tian, S.W.; Hou, L.Y.; Huang, X.Z.; Zhang, X.Y.; Guo, H.W.; Yang, S.H. Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and type-A ARR genes in Arabidopsis. Plant Cell 2012, 24, 2578–2595. [Google Scholar] [CrossRef]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Stockinger, E.J.; Gilmour, S.J.; Thomashow, M.F. Arabidopsis thaliana CBF1 encodes an AP2 domain-containing transcriptional activator that binds to the C-repeat/DRE, a cis-acting DNA regulatory element that stimulates transcription in response to low temperature and water deficit. Proc. Natl. Acad. Sci. USA 1997, 94, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.B.; Ding, Y.L.; Shi, Y.T.; Ma, L.; Wang, Y.; Song, C.P.; Wilkins, K.A.; Davies, J.M.; Knight, H.; Knight, M.R.; et al. The calcium transporter ANNEXIN1 mediates cold-induced calcium signaling and freezing tolerance in plants. Embo J. 2021, 40, e104559. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Yang, H.; Wu, S.; Fu, D.; Li, M.; Gong, Z.; Yang, S. CPK28-NLP7 module integrates cold-induced Ca(2+) signal and transcriptional reprogramming in Arabidopsis. Sci. Adv. 2022, 8, eabn7901. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.Q.; Liu, Y.M.; Mu, Y.; Anwar, A.; He, C.X.; Yan, Y.; Li, Y.S.; Yu, X.C. Heterotrimeric G-protein γ subunit CsGG3.2 positively regulates the expression of genes and chilling tolerance in Cucumber. Front. Plant Sci. 2018, 9, 488. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Li, H.M.; Quan, X.Y.; Shan, Q.L.; Wang, W.B.; Yin, N.; Wang, S.Q.; Wang, Z.H.; He, W.X. Comprehensive analysis of cucumber C-repeat/dehydration-responsive element binding factor family genes and their potential roles in cold tolerance of cucumber. BMC Plant Biol. 2022, 22, 270. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Tang, X.L.; Huo, Y.Q.; Xu, R.; Qi, S.D.; Huang, J.G.; Zheng, C.C.; Wu, C.A. Identification and characterization of fructose 1,6-bisphosphate aldolase genes in Arabidopsis reveal a gene family with diverse responses to abiotic stresses. Gene 2012, 503, 65–74. [Google Scholar] [CrossRef]
- Cai, B.B.; Li, Q.; Xu, Y.C.; Yang, L.; Bi, H.G.; Ai, X.Z. Genome-wide analysis of the fructose 1,6-bisphosphate aldolase (FBA) gene family and functional characterization of FBA7 in tomato. Plant Physiol. Biochem. 2016, 108, 251–265. [Google Scholar] [CrossRef]
- Zhang, Y. Functional Analysis of a Fructose-1, 6-Diphosphatase Aldolase Gene ALDY in Rice. Master’s Dissertation, Huazhong Agricultural University, Wuhan, China, 2014; pp. 34–37. [Google Scholar]
- Kelley, P.M.; Freeling, M. Anaerobic expression of maize fructose-1,6-diphosphate aldolase. J. Biol. Chem. 1984, 259, 14180–14183. [Google Scholar] [CrossRef]
- Lebherz, H.G.; Leadbetter, M.M.; Bradshaw, R.A. Isolation and characterization of the cytosolic and chloroplast forms of spinach leaf fructose diphosphate aldolase. J. Biol. Chem. 1984, 259, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.A.; Wong, D.M.; Sachs, M.M. The anaerobic response of soybean. Plant Physiol. 1990, 92, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Haake, V.; Zrenner, R.; Sonnewald, U.; Stitt, M. A moderate decrease of plastid aldolase activity inhibits photosynthesis, alters the levels of sugars and starch, and inhibits growth of potato plants. Plant J. 1998, 14, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Michelis, R.; Gepstein, S. Identification and characterization of a heat-induced isoform of aldolase in oat chloroplast. Plant Mol. Biol. 2000, 44, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Komori, T.; Hashimoto, A.; Kuwata, S.; Imaseki, H.; Kubo, T. Differential expression of plastidic aldolase genes in plants under salt stress. Plant Sci. 2000, 154, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Zhang, Z.; Zhang, Y. Cloning and molecular characterization of fructose-1,6-bisphosphate aldolase gene regulated by high-salinity and drought in Sesuvium portulacastrum. Plant Cell Rep. 2009, 28, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Lv, K. The Primary Study of the Wheat Fructose-1,6-Bisphoshate Aldolase from Albinism Line. Master’s Dissertation, Northwest Agriculture & Forestry University, Yangling, China, 2011; pp. 35–37. [Google Scholar]
- Lao, X.T.; Azuma, J.; Sakamoto, M. Two cytosolic aldolases show different expression patterns during shoot elongation in Moso bamboo, Phyllostachys pubescens Mazel. Physiol. Plant. 2013, 149, 422–431. [Google Scholar] [CrossRef]
- Zeng, Y.L.; Tan, X.F.; Zhang, L.; Jiang, N.; Cao, H.P. Identification and expression of fructose-1,6-bisphosphate aldolase genes and their relations to oil content in developing seeds of tea oil tree (Camellia oleifera). PLoS ONE 2014, 9, e107422. [Google Scholar] [CrossRef] [PubMed]
- Khanna, S.M.; Taxak, P.C.; Jain, P.K.; Saini, R.; Srinivasan, R. Glycolytic enzyme activities and gene expression in cicer arietinum exposed to water-deficit stress. Appl. Biochem. Biotechnol. 2014, 173, 2241–2253. [Google Scholar] [CrossRef]
- Cai, B.B.; Li, Q.; Liu, F.J.; Bi, H.A.; Ai, X.Z. Decreasing fructose-1,6-bisphosphate aldolase activity reduces plant growth and tolerance to chilling stress in tomato seedlings. Physiol. Plant. 2018, 163, 247–258. [Google Scholar] [CrossRef]
- Cai, B.B.; Ning, Y.; Li, Q.; Li, Q.Y.; Ai, X.Z. Effects of the Chloroplast Fructose-1,6-Bisphosphate Aldolase Gene on Growth and Low-Temperature Tolerance of Tomato. Int. J. Mol. Sci. 2022, 23, 728. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, M.T.; Liu, Q.Y.; Niu, B. Hydrological cycling optimization-based multiobjective feature-selection method for customer segmentation. Int. J. Intell. Syst. 2021, 36, 2347–2366. [Google Scholar] [CrossRef]
- Huang, H.H.; Wu, N.Q.; Liang, Y.; Peng, X.D.; Jun, S. SLNL: A novel method for gene selection and phenotype classification. Int. J. Intell. Syst. 2022, 37, 6283–6304. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford; UK; New York, NY, USA, 2000. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.N.; Liu, Y.K.; Xue, Z.J.; Li, Q.Y.; Cai, B.B. Structures, characteristics and functions of fructose-1,6-bisphosphate aldolase in various tissues. Acta Soc. Bot. Pol. 2023, 92, 174253. [Google Scholar] [CrossRef]
- Xing, H.Y.; Pudake, R.N.; Guo, G.G.; Xing, G.F.; Hu, Z.R.; Zhang, Y.R.; Sun, Q.X.; Ni, Z.F. Genome-wide identification and expression profiling of auxin response factor (ARF) gene family in maize. BMC Genom. 2011, 12, 178. [Google Scholar] [CrossRef] [PubMed]
- Carrera, D.A.; George, G.M.; Fischer-Stettler, M.; Galbier, F.; Eicke, S.; Truernit, E.; Streb, S.; Zeeman, S.C. Distinct plastid fructose bisphosphate aldolases function in photosynthetic and non-photosynthetic metabolism in Arabidopsis. J. Exp. Bot. 2021, 72, 3739–3755. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.M. Fructose Biphosphate Aldolase Regulates the Stem Cell Function in Arabidopsis Thaliana. Master’s Dissertation, Xiamen University, Xiamen, China, 2020. [Google Scholar]
- Liu, W.; Deng, Y.; Li, Y.; Yang, L.; Zhu, L.; Jiang, L. Coupling protein scaffold and biosilicification: A sustainable and recyclable approach for d-mannitol production via one-step purification and immobilization of multienzymes. Int. J. Biol. Macromol. 2024, 269, 132196. [Google Scholar] [CrossRef]
- Liu, F.J.; Zhang, X.W.; Cai, B.B.; Pan, D.Y.; Fu, X.; Bi, H.G.; Ai, X.Z. Physiological response and transcription profiling analysis reveal the role of glutathione in HS-induced chilling stress tolerance of cucumber seedlings. Plant Sci. 2020, 291, 110363. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).