


Comparative Analysis of Acetylated Flavonoids’ Chemopreventive Effects in Different Cancer Cell Lines



Abstract
:1. Introduction
2. Results
2.1. Synthesis and Structural Characterization of Acetyl Flavonoids
2.2. Acetylated Flavonoids Inhibit the Proliferation of Three Types of Cancer
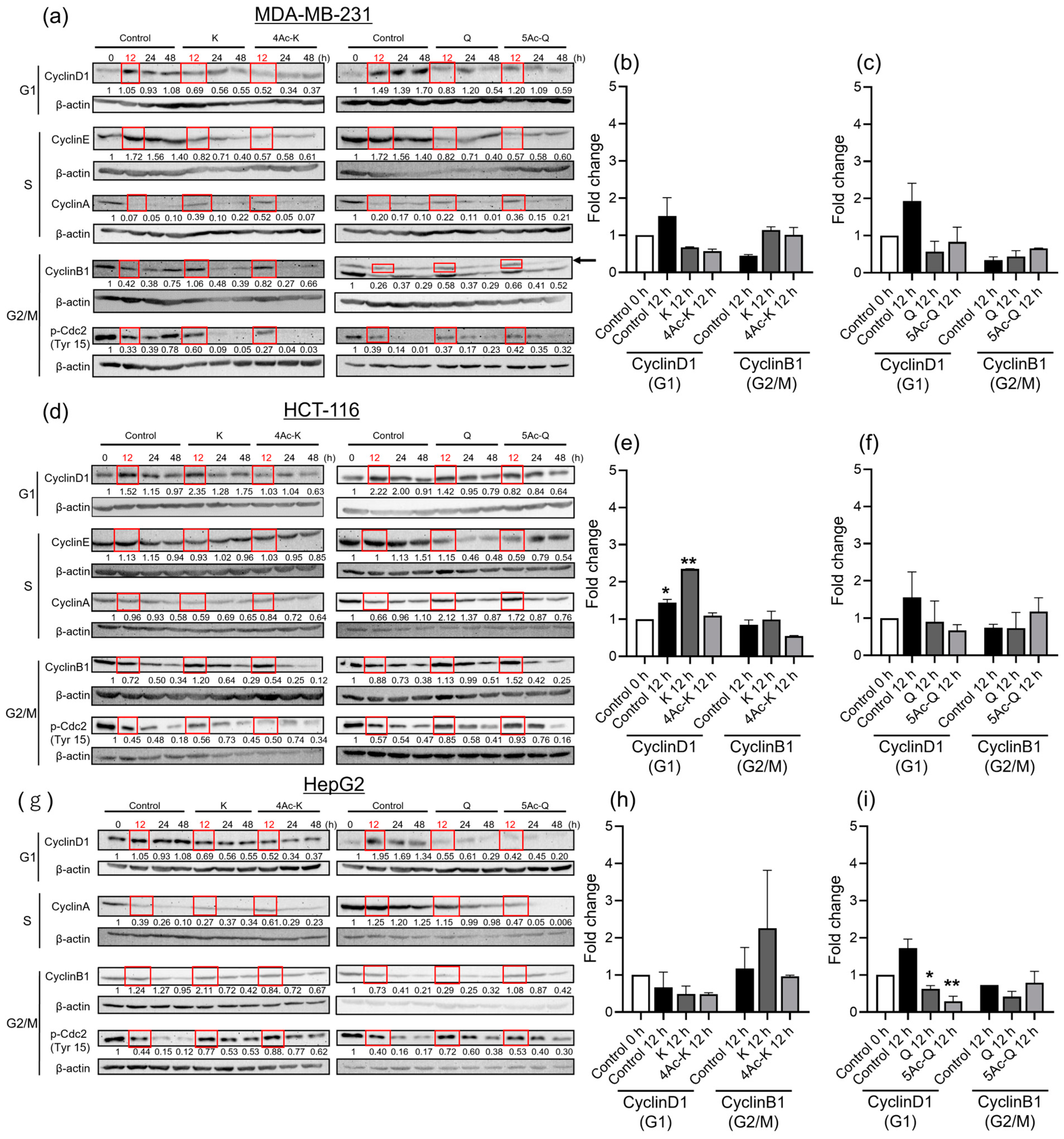
2.3. Effects of Flavonoid Acetylation on Cell Cycle Progression-Related Proteins
2.4. Screening of Acetylated Flavonoids for Anti-Migration Properties
2.5. Synthesis of Acetylated Compounds with Retained 5-Hydroxy Group
2.6. Effect of Position 5 Hydroxyl Group on Flavonoids’ Anti-Proliferative Activity
2.7. Effects of 5-Position Hydroxyl Group Acetylation on Anti-Migration Activity
2.8. Acetylated Flavonoids Induce Apoptosis
3. Discussion
3.1. Cell Proliferation Inhibitory Ability and Structure–Activity Relationships
3.2. Effects of Acetylation of Quercetin and Kaempferol on the Cell Cycle
3.3. Anti-Migration and Apoptosis-Inducing Activities of Acetylated Flavonoids
3.4. Anti-Migration Activity and Structure–Activity Relationships
4. Materials and Methods
4.1. Chemicals, Antibodies, and Reagents
4.2. Synthesis of Acetylated Derivatives of Flavonoids
4.2.1. 3,5,7,4′-. O-tetraacetate kaempferol (4Ac-K) (Compound 2)
4.2.2. 3,5,7,3′,4-. O-pentaacetate quercetin (5Ac-Q) (Compound 4)
4.2.3. 3,5,7,3′,4′,5′-. O-hexaacetate myricetin (6Ac-M) (Compound 6)
4.2.4. 5,7-. O-diacetate chrysin (2Ac-C) (Compound 8)
4.2.5. 5,7,4′-. O-triacetate apigenin (3Ac-A) (Compound 10)
4.2.6. 5,7,3’,4′-. O-tetraacetate luteolin (4Ac-L) (Compound 12)
4.2.7. 5,7,4′-. O-triacetate naringenin (3Ac-N) (Compound 14)
4.2.8. 3,5,7,3,’4′-. O-pentaacetate taxifolin (5Ac-T) (Compound 16)
4.2.9. 3,7,4′-. O-triacetate kaempferol (3Ac-K) (Compound 17)
4.2.10. 3,7,3′,4′-. O-tetraacetate quercetin (4A-Q) (Compound 18)
4.2.11. 3,7,3′,4′,5′-. O-pentaacetate myricetin (5Ac-M) (Compound 19)
4.2.12. 7,4′-. O-diacetate apigenin (2Ac-A) (Compound 20)
4.3. Structure Identification of Acetylated Flavonoids
4.4. Cell Culture
4.5. Cell Viability Assay
4.6. Western Blot Analysis
4.7. Detection of Apoptosis Using Flow Cytometry
4.8. Wound Healing Assay
4.9. Cell Migration Assay
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Aherne, S.A.; O’Brien, N.M. Dietary flavonols: Chemistry, food content, and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Bei, R.; Masuelli, L.; Turriziani, M.; Volti, L.G.; Malaguarnera, M.; Galvano, F. Impaired Expression and Function of Signaling Pathway Enzymes by Anthocyanins: Role on Cancer Prevention and Progression. Curr. Enzym. Inhib. 2009, 5, 184–197. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: A mechanism including AMPKα1/SIRT1. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, R.; Shi, Y.; Zhang, X.; Tian, C.; Xia, D. Antioxidant and Anti-Inflammatory Activities of Six Flavonoids from Smilax glabra Roxb. Molecules 2020, 25, 5295. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros, M.; Rinaldi, D.; Marder, M.; Espelt, M.V.; Mangialavori, I.; Vigil, M.; Rossi, J.P.; Ferreira-Gomes, M. Natural flavonoids inhibit the plasma membrane Ca2+-ATPase. Biochem. Pharmacol. 2019, 166, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.-J.; Daikonya, A.; Ohkawara, M.; Nemoto, T.; Noritake, R.; Takamiya, T.; Kitanaka, S.; Iijima, H. Structure-activity relationship of the inhibitory effects of flavonoids on nitric oxide production in RAW264.7 cells. Bioorgan. Med. Chem. 2017, 25, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Desmet, S.; Morreel, K.; Dauwe, R. Origin and Function of Structural Diversity in the Plant Specialized Metabolome. Plants 2021, 10, 2393. [Google Scholar] [CrossRef] [PubMed]
- Alseekh, S.; Perez de Souza, L.; Benina, M.; Fernie, A.R. The style and substance of plant flavonoid decoration; towards defining both structure and function. Phytochemistry 2020, 174, 112347. [Google Scholar] [CrossRef]
- Wang, S.; Alseekh, S.; Fernie, A.R.; Luo, J. The Structure and Function of Major Plant Metabolite Modifications. Mol. Plant 2019, 12, 899–919. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Tsai, P.-H.; Lin, C.-Y.; Cheng, C.-H.; Lin, T.-H.; Lee, K.P.H.; Huang, K.-Y.; Chen, S.-H.; Hwang, J.-J.; Kandaswami, C.C.; et al. Impact of Flavonoids on Matrix Metalloproteinase Secretion and Invadopodia Formation in Highly Invasive A431-III Cancer Cells. PLoS ONE 2013, 8, e71903. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.-H.; Chen, L.-C.; Ho, Y.-S. An apple a day to prevent cancer formation: Reducing cancer risk with flavonoids. J. Food Drug Anal. 2017, 25, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Khan, H.; Ullah, H.; Hassan, S.T.S.; Šmejkal, K.; Efferth, T.; Mahomoodally, M.F.; Xu, S.; Habtemariam, S.; Filosa, R.; et al. MicroRNA targeting by quercetin in cancer treatment and chemoprotection. Pharmacol. Res. 2019, 147, 104346. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.-S.; Jiang, F.-S.; Zhang, K.; Zhu, X.-X.; Jin, B.; Lu, J.-J.; Ding, Z.-S. Flavonoids from the leaves of Carya cathayensis Sarg. inhibit vascular endothelial growth factor-induced angiogenesis. Fitoterapia 2014, 92, 34–40. [Google Scholar] [CrossRef]
- Fatima, N.; Baqri, S.S.R.; Bhattacharya, A.; Koney, N.K.; Husain, K.; Abbas, A.; Ansari, R.A. Role of Flavonoids as Epigenetic Modulators in Cancer Prevention and Therapy. Front. Genet. 2021, 12, 758733. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.; Burkard, M.; Leischner, C.; Lauer, U.M.; Frank, J.; Venturelli, S. Epigenetic activities of flavonoids in the prevention and treatment of cancer. Clin. Epigenetics 2015, 7, 64. [Google Scholar] [CrossRef]
- Fink, B.N.; Steck, S.E.; Wolff, M.S.; Britton, J.A.; Kabat, G.C.; Gaudet, M.M.; Abrahamson, P.E.; Bell, P.; Schroeder, J.C.; Teitelbaum, S.L.; et al. Dietary Flavonoid Intake and Breast Cancer Survival among Women on Long Island. Cancer Epidemiol. Biomark. Prev. 2007, 16, 2285–2292. [Google Scholar] [CrossRef]
- Amawi, H.; Ashby, C.R., Jr.; Tiwari, A.K. Cancer chemoprevention through dietary flavonoids: What’s limiting? Chin. J. Cancer 2017, 36, 50. [Google Scholar] [CrossRef]
- Lewandowska, U.; Fichna, J.; Gorlach, S. Enhancement of anticancer potential of polyphenols by covalent modifications. Biochem. Pharmacol. 2016, 109, 1–13. [Google Scholar] [CrossRef]
- Fernández, C.; Nieto, O.; Rivas, E.; Montenegro, G.; Fontenla, J.A.; Fernández-Mayoralas, A. Synthesis and biological studies of glycosyl dopamine derivatives as potential antiparkinsonian agents. Carbohydr. Res. 2000, 327, 353–365. [Google Scholar] [CrossRef]
- Koirala, N.; Pandey, R.P.; Van Thang, D.; Jung, H.J.; Sohng, J.K. Glycosylation and subsequent malonylation of isoflavonoids in E. coli: Strain development, production and insights into future metabolic perspectives. J. Ind. Microbiol. Biotechnol. 2014, 41, 1647–1658. [Google Scholar] [CrossRef] [PubMed]
- Needs, P.W.; Williamson, G. Syntheses of daidzein-7-yl β-d-glucopyranosiduronic acid and daidzein-4′,7-yl di-β-d-glucopyranosiduronic acid. Carbohydr. Res. 2001, 330, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Sakao, K.; Hamamoto, S.; Urakawa, D.; He, Z.; Hou, D.X. Anticancer Activity and Molecular Mechanisms of Acetylated and Methylated Quercetin in Human Breast Cancer Cells. Molecules 2024, 29, 2408. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Jiang, Y.; Yang, J.; Zhao, Y.; Tian, M.; Yang, B. Structure, bioactivity, and synthesis of methylated flavonoids. Ann. N. Y. Acad. Sci. 2017, 1398, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K.; Mitsunaga, T.; Afroze, S.H.; Uddin, M.N. Structure—Activity Relationships of Methylquercetin on Anti-migration and Anti-proliferation Activity in B16 Melanoma Cells. Anticancer. Res. 2017, 37, 1575–1579. [Google Scholar] [CrossRef]
- Ninomiya, M.; Nishida, K.; Tanaka, K.; Watanabe, K.; Koketsu, M. Structure-activity relationship studies of 5,7-dihydroxyflavones as naturally occurring inhibitors of cell proliferation in human leukemia HL-60 cells. J. Nat. Med. 2013, 67, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Koirala, N.; Thuan, N.H.; Ghimire, G.P.; Thang, D.V.; Sohng, J.K. Methylation of flavonoids: Chemical structures, bioactivities, progress and perspectives for biotechnological production. Enzym. Microb. Technol. 2016, 86, 103–116. [Google Scholar] [CrossRef]
- Chen, L.; Teng, H.; Xie, Z.; Cao, H.; Cheang, W.S.; Skalicka-Woniak, K.; Georgiev, M.I.; Xiao, J. Modifications of dietary flavonoids towards improved bioactivity: An update on structure–activity relationship. Crit. Rev. Food Sci. Nutr. 2018, 58, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; He, Y.; Zhu, S.; Zhong, X.; Chen, D.; Liu, Z. New Acylglycosides Flavones from Fuzhuan Brick Tea and Simulation Analysis of Their Bioactive Effects. Int. J. Mol. Sci. 2019, 20, 494. [Google Scholar] [CrossRef]
- El-Kersh, D.M.; Abou El-Ezz, R.F.; Fouad, M.; Farag, M.A. Unveiling Natural and Semisynthetic Acylated Flavonoids: Chemistry and Biological Actions in the Context of Molecular Docking. Molecules 2022, 27, 5501. [Google Scholar] [CrossRef]
- Suzuki, H.; Nakayama, T.; Yonekura-Sakakibara, K.; Fukui, Y.; Nakamura, N.; Yamaguchi, M.-a.; Tanaka, Y.; Kusumi, T.; Nishino, T. cDNA Cloning, Heterologous Expressions, and Functional Characterization of Malonyl-Coenzyme A:Anthocyanidin 3-O-Glucoside-6″-O-Malonyltransferase from Dahlia Flowers. Plant Physiol. 2002, 130, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Sendri, N.; Bhandari, P. Anthocyanins: A comprehensive review on biosynthesis, structural diversity, and industrial applications. Phytochem. Rev. 2024, 23, 1–62. [Google Scholar] [CrossRef]
- Liu, X.; Shi, Y.; Hu, H.; Li, J.; Li, C.; Yang, L.; Wang, Z. Three new C-glycosyflavones with acetyl substitutions from Swertia mileensis. J. Nat. Med. 2018, 72, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Le, J.; Lu, W.; Xiong, X.; Wu, Z.; Chen, W. Anti-Inflammatory Constituents from Bidens frondosa. Molecules 2015, 20, 18496–18510. [Google Scholar] [CrossRef] [PubMed]
- Sakao, K.; Hou, D.-X. Effects and Mechanisms of Acylated Flavonoid on Cancer Chemopreventive Activity. Curr. Pharmacol. Rep. 2020, 6, 286–305. [Google Scholar] [CrossRef]
- Massi, A.; Bortolini, O.; Ragno, D.; Bernardi, T.; Sacchetti, G.; Tacchini, M.; De Risi, C. Research Progress in the Modification of Quercetin Leading to Anticancer Agents. Molecules 2017, 22, 1270. [Google Scholar] [CrossRef] [PubMed]
- Sadilova, E.; Stintzing, F.C.; Carle, R. Thermal Degradation of Acylated and Nonacylated Anthocyanins. J. Food Sci. 2006, 71, C504–C512. [Google Scholar] [CrossRef]
- Jokioja, J.; Yang, B.; Linderborg, K.M. Acylated anthocyanins: A review on their bioavailability and effects on postprandial carbohydrate metabolism and inflammation. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5570–5615. [Google Scholar] [CrossRef] [PubMed]
- Tu, F.; Xie, C.; Li, H.; Lei, S.; Li, J.; Huang, X.; Yang, F. Effect of in vitro digestion on chestnut outer-skin and inner-skin bioaccessibility: The relationship between biotransformation and antioxidant activity of polyphenols by metabolomics. Food Chem. 2021, 363, 130277. [Google Scholar] [CrossRef]
- Cho, S.; Kong, B.; Jung, Y.; Shin, J.; Park, M.; Chung, W.J.; Ban, C.; Kweon, D.H. Synthesis and physicochemical characterization of acyl myricetins as potential anti-neuroexocytotic agents. Sci. Rep. 2023, 13, 5136. [Google Scholar] [CrossRef]
- Sakao, K.; Saruwatari, H.; Minami, S.; Hou, D.X. Hydroxyl Group Acetylation of Quercetin Enhances Intracellular Absorption and Persistence to Upregulate Anticancer Activity in HepG2 Cells. Int. J. Mol. Sci. 2023, 24, 16652. [Google Scholar] [CrossRef] [PubMed]
- Sakao, K.; Fujii, M.; Hou, D.X. Acetyl derivate of quercetin increases the sensitivity of human leukemia cells toward apoptosis. Biofactors 2009, 35, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Meng, W.-D.; Xu, Y.-Y.; Cao, J.-G.; Qing, F.-L. Synthesis and anticancer effect of chrysin derivatives. Bioorgan. Med. Chem. Lett. 2003, 13, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.K.-D.; Huynh, T.K.-C.; Nguyen, T.-D. Synthesis, characterization, anti-inflammatory and anti-proliferative activity against MCF-7 cells of O-alkyl and O-acyl flavonoid derivatives. Bioorgan. Chem. 2015, 63, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Calderaro, A.; Patanè, G.T.; Navarra, M.; Barreca, D.; Cirmi, S.; Felice, M.R. Targets Involved in the Anti-Cancer Activity of Quercetin in Breast, Colorectal and Liver Neoplasms. Int. J. Mol. Sci. 2023, 24, 2952. [Google Scholar] [CrossRef] [PubMed]
- Adinew, G.M.; Taka, E.; Mendonca, P.; Messeha, S.S.; Soliman, K.F.A. The Anticancer Effects of Flavonoids through miRNAs Modulations in Triple-Negative Breast Cancer. Nutrients 2021, 13, 1212. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yan, T.; Deng, R.; Jiang, X.; Xiong, H.; Wang, Y.; Yu, Q.; Wang, X.; Chen, C.; Zhu, Y. Low dose of kaempferol suppresses the migration and invasion of triple-negative breast cancer cells by downregulating the activities of RhoA and Rac1. OncoTargets Ther. 2017, 10, 4809–4819. [Google Scholar] [CrossRef] [PubMed]
- Shahi Thakuri, P.; Gupta, M.; Singh, S.; Joshi, R.; Glasgow, E.; Lekan, A.; Agarwal, S.; Luker, G.D.; Tavana, H. Phytochemicals inhibit migration of triple negative breast cancer cells by targeting kinase signaling. BMC Cancer 2020, 20, 4. [Google Scholar] [CrossRef]
- Sharma, P.; Khan, M.A.; Najmi, A.K.; Chaturvedi, S.; Akhtar, M. Myricetin-induced apoptosis in triple-negative breast cancer cells through inhibition of the PI3K/Akt/mTOR pathway. Med. Oncol. 2022, 39, 248. [Google Scholar] [CrossRef]
- Cao, D.; Zhu, G.-Y.; Lu, Y.; Yang, A.; Chen, D.; Huang, H.-J.; Peng, S.-X.; Chen, L.-W.; Li, Y.-W. Luteolin suppresses epithelial-mesenchymal transition and migration of triple-negative breast cancer cells by inhibiting YAP/TAZ activity. Biomed. Pharmacother. 2020, 129, 110462. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wei, Y.X.; Shen, M.C.; Tu, Y.H.; Wang, C.C.; Huang, H.C. Chrysin, Abundant in Morinda citrifolia Fruit Water-EtOAc Extracts, Combined with Apigenin Synergistically Induced Apoptosis and Inhibited Migration in Human Breast and Liver Cancer Cells. J. Agric. Food Chem. 2016, 64, 4235–4245. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, A.H.; Alsahli, M.A.; Almatroudi, A.; Almogbel, M.A.; Khan, A.A.; Anwar, S.; Almatroodi, S.A. The Potential Role of Apigenin in Cancer Prevention and Treatment. Molecules 2022, 27, 6051. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Kong, S.; Zhao, S.; Tang, Q. Naringenin inhibits human breast cancer cells (MDA-MB-231) by inducing programmed cell death, caspase stimulation, G2/M phase cell cycle arrest and suppresses cancer metastasis. Cell. Mol. Biol. 2021, 67, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, L.; Zhou, T.; Gong, X.; Jiang, R.; Li, H.; Kuang, G.; Wan, J.; Li, H. Taxifolin inhibits breast cancer cells proliferation, migration and invasion by promoting mesenchymal to epithelial transition via β-catenin signaling. Life Sci. 2019, 232, 116617. [Google Scholar] [CrossRef]
- Sudan, S.; Rupasinghe, H.V. Antiproliferative activity of long chain acylated esters of quercetin-3-O-glucoside in hepatocellular carcinoma HepG2 cells. Exp. Biol. Med. 2015, 240, 1452–1464. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.D.; Sang, S.; Hong, J.; Kwon, S.-J.; Lee, M.-J.; Ho, C.-T.; Yang, C.S. Peracetylation as a Means of Enhancing in Vitro Bioactivity and Bioavailability of Epigallocatechin-3-Gallate. Drug Metab. Dispos. 2006, 34, 2111. [Google Scholar] [CrossRef] [PubMed]
- Tavsan, Z.; Kayali, H.A. Flavonoids showed anticancer effects on the ovarian cancer cells: Involvement of reactive oxygen species, apoptosis, cell cycle and invasion. Biomed. Pharmacother. 2019, 116, 109004. [Google Scholar] [CrossRef] [PubMed]
- Plochmann, K.; Korte, G.; Koutsilieri, E.; Richling, E.; Riederer, P.; Rethwilm, A.; Schreier, P.; Scheller, C. Structure–activity relationships of flavonoid-induced cytotoxicity on human leukemia cells. Arch. Biochem. Biophys. 2007, 460, 1–9. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Identification of Six Flavonoids as Novel Cellular Antioxidants and Their Structure-Activity Relationship. Oxidative Med. Cell. Longev. 2020, 2020, 4150897. [Google Scholar] [CrossRef]
- Karancsi, Z.; Kovács, D.; Palkovicsné Pézsa, N.; Gálfi, P.; Jerzsele, Á.; Farkas, O. The Impact of Quercetin and Its Methylated Derivatives 3-o-Methylquercetin and Rhamnazin in Lipopolysaccharide-Induced Inflammation in Porcine Intestinal Cells. Antioxidants 2022, 11, 1265. [Google Scholar] [CrossRef] [PubMed]
- Rusak, G.; Gutzeit, H.O.; Müller, J.L. Structurally related flavonoids with antioxidative properties differentially affect cell cycle progression and apoptosis of human acute leukemia cells. Nutr. Res. 2005, 25, 143–155. [Google Scholar] [CrossRef]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Antiproliferative activity of flavonoids on several cancer cell lines. Biosci. Biotechnol. Biochem. 1999, 63, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Baidya, R.; Chakraborty, T.; Samanta, A.K.; Roy, S. Pharmacological basis and new insights of taxifolin: A comprehensive review. Biomed. Pharmacother. 2021, 142, 112004. [Google Scholar] [CrossRef]
- Shi, M.D.; Shiao, C.K.; Lee, Y.C.; Shih, Y.W. Apigenin, a dietary flavonoid, inhibits proliferation of human bladder cancer T-24 cells via blocking cell cycle progression and inducing apoptosis. Cancer Cell Int. 2015, 15, 33. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Meng, X.; Zheng, H.; Zeng, Q.; Chen, T.; Wang, W.; Zhang, X.; Su, J. Kaempferol Attenuates ROS-Induced Hemolysis and the Molecular Mechanism of Its Induction of Apoptosis on Bladder Cancer. Molecules 2018, 23, 2592. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.H.; Chien, M.H.; Lin, W.L.; Wen, Y.C.; Chow, J.M.; Chen, C.K.; Kuo, T.C.; Lee, W.J. Inhibition of MDA-MB-231 breast cancer cell proliferation and tumor growth by apigenin through induction of G2/M arrest and histone H3 acetylation-mediated p21(WAF1/CIP1) expression. Environ. Toxicol. 2017, 32, 434–444. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Xue, L. Kaempferol Suppresses Proliferation and Induces Cell Cycle Arrest, Apoptosis, and DNA Damage in Breast Cancer Cells. Oncol. Res. 2019, 27, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Nandi, S.K.; Roychowdhury, T.; Chattopadhyay, S.; Basu, S.; Chatterjee, K.; Choudhury, P.; Banerjee, N.; Saha, P.; Mukhopadhyay, S.; Mukhopadhyay, A.; et al. Deregulation of the CD44-NANOG-MDR1 associated chemoresistance pathways of breast cancer stem cells potentiates the anti-cancer effect of Kaempferol in synergism with Verapamil. Toxicol. Appl. Pharmacol. 2022, 437, 115887. [Google Scholar] [CrossRef]
- Chekuri, S.; Vyshnava, S.S.; Somisetti, S.L.; Cheniya, S.B.K.; Gandu, C.; Anupalli, R.R. Isolation and anticancer activity of quercetin from Acalypha indica L. against breast cancer cell lines MCF-7 and MDA-MB-231. 3 Biotech 2023, 13, 289. [Google Scholar] [CrossRef]
- Leong, K.X.; Chao, S.P.; Siah, P.C.; Lim, S.K.; Khoo, B.Y. Comparative Studies on a Standardized Subfraction of Red Onion Peel Ethanolic Extract (Plant Substance), Quercetin (Pure Compound), and Their Cell Mechanism and Metabolism on MDA-MB-231. Evid.-Based Complement. Altern. Med. 2022, 2022, 9284063. [Google Scholar] [CrossRef] [PubMed]
- Budisan, L.; Gulei, D.; Jurj, A.; Braicu, C.; Zanoaga, O.; Cojocneanu, R.; Pop, L.; Raduly, L.; Barbat, A.; Moldovan, A.; et al. Inhibitory Effect of CAPE and Kaempferol in Colon Cancer Cell Lines-Possible Implications in New Therapeutic Strategies. Int. J. Mol. Sci. 2019, 20, 1199. [Google Scholar] [CrossRef] [PubMed]
- Chiou, Y.-S.; Sang, S.; Cheng, K.-H.; Ho, C.-T.; Wang, Y.-J.; Pan, M.-H. Peracetylated (−)-epigallocatechin-3-gallate (AcEGCG) potently prevents skin carcinogenesis by suppressing the PKD1-dependent signaling pathway in CD34 + skin stem cells and skin tumors. Carcinogenesis 2013, 34, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Jia, Y.; Liu, J.; Ren, X.; Yang, X.; Xia, X.; Pan, X. Naringenin and Quercetin Exert Contradictory Cytoprotective and Cytotoxic Effects on Tamoxifen-Induced Apoptosis in HepG2 Cells. Nutrients 2022, 14, 5394. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-J.; Chen, W.-K.; Wang, C.-J.; Lin, W.-L.; Tseng, T.-H. Apigenin inhibits HGF-promoted invasive growth and metastasis involving blocking PI3K/Akt pathway and β4 integrin function in MDA-MB-231 breast cancer cells. Toxicol. Appl. Pharmacol. 2008, 226, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Jung, J.; Moon, A.; Kang, H.; Cho, H. Antitumor and Anti-Invasive Effect of Apigenin on Human Breast Carcinoma through Suppression of IL-6 Expression. Int. J. Mol. Sci. 2019, 20, 3143. [Google Scholar] [CrossRef] [PubMed]
- Naponelli, V.; Rocchetti, M.T.; Mangieri, D. Apigenin: Molecular Mechanisms and Therapeutic Potential against Cancer Spreading. Int. J. Mol. Sci. 2024, 25, 5569. [Google Scholar] [CrossRef] [PubMed]
- Phromnoi, K.; Yodkeeree, S.; Anuchapreeda, S.; Limtrakul, P. Inhibition of MMP-3 activity and invasion of the MDA-MB-231 human invasive breast carcinoma cell line by bioflavonoids. Acta Pharmacol. Sin. 2009, 30, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Jeon, Y.K.; Nam, M.J. A mechanism of apigenin-induced apoptosis is potentially related to anti-angiogenesis and anti-migration in human hepatocellular carcinoma cells. Food Chem. Toxicol. 2011, 49, 1626–1632. [Google Scholar] [CrossRef]
- Sakao, K.; Desineni, S.; Hahm, E.R.; Singh, S.V. Phenethyl isothiocyanate suppresses inhibitor of apoptosis family protein expression in prostate cancer cells in culture and in vivo. Prostate 2012, 72, 1104–1116. [Google Scholar] [CrossRef]
- Tanigawa, S.; Fujii, M.; Hou, D.X. Stabilization of p53 is involved in quercetin-induced cell cycle arrest and apoptosis in HepG2 cells. Biosci. Biotechnol. Biochem. 2008, 72, 797–804. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | |||
|---|---|---|---|
| Compound | MDA-MB-231 | HCT-116 | HepG2 |
| Flavonols (IC50 μM) | |||
| K | 46.7 ± 6.8 | 34.9 ± 1.0 | 33.4 ± 2.1 |
| 4Ac-K | 33.6 ± 0.6 | 28.5 ± 0.5 | 23.2 ± 0.1 |
| Q | 24.3 ± 9.6 | 23.5 ± 2.8 | 28.2 ± 5.0 |
| 5Ac-Q | 17.4 ± 10.8 | 15.7 ± 2.6 | 15.5 ± 0.2 |
| M | 27.2 ± 0.3 | Over 160 | Over 160 |
| 6Ac-M | 50.9 ± 5.5 | 81.7 ± 36 | 76.6 ± 20.4 |
| Flavone (IC50 μM) | |||
| C | 37.5 ± 5.0 | 27.5 ± 2.9 | 25.3 ± 2.8 |
| 2Ac-C | 38.4 ± 5.4 | 32.2 ± 1.3 | 23.8 ± 9.0 |
| A | 27.1 ± 5.2 | 19.0 ± 4.2 | 25.8 ± 1.9 |
| 3Ac-A | 31.1 ± 7.2 | 21.9 ± 2.6 | 6.5 ± 1.0 |
| L | 12.9 ± 3.6 | 9.3 ± 2.2 | 10.2 ± 2.0 |
| 4Ac-L | 20.2 ± 6.3 | 12.2 ± 3.7 | 12.4 ± 3.5 |
| Flavanone (IC50 μM) | |||
| N | Over 160 | 120.4 ± 39.8 | 118.5 ± 30.8 |
| 3Ac-N | 156.4 ± 23.1 | 94.3 ± 23.1 | 97.9 ± 10.6 |
| T | Over 160 | Over 160 | Over 160 |
| 5Ac-T | 128.0 ± 23.1 | 125.8 ± 23.1 | 152.2 ± 23.1 |
| Sample | MDA-MB-231 | HCT-116 | HepG2 |
|---|---|---|---|
| Flavonols | |||
| 3Ac-K | ↑ | - | - |
| 4Ac-K | ↑ | ↑ | ↑ |
| 4Ac-Q | ↑ | - | - |
| 5Ac-Q | ↑ | ↑ | ↑ |
| 5Ac-M | ↓ | - | - |
| 6Ac-M | ↓ | ↑ | ↑ |
| Flavone | |||
| 2Ac-C | ↓ | ↓ | ↑ |
| 2Ac-A | ↑ | - | - |
| 3Ac-A | ↓ | ↓ | ↑ |
| 4Ac-L | ↓ | ↓ | ↓ |
| Flavanone | |||
| 3Ac-N | ↑ | → | → |
| 5Ac-T | ↑ | ↑ | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urakawa, D.; Shioiridani, Y.; Igata, S.; Hou, D.-X.; Sakao, K. Comparative Analysis of Acetylated Flavonoids’ Chemopreventive Effects in Different Cancer Cell Lines. Int. J. Mol. Sci. 2024, 25, 7689. https://doi.org/10.3390/ijms25147689
Urakawa D, Shioiridani Y, Igata S, Hou D-X, Sakao K. Comparative Analysis of Acetylated Flavonoids’ Chemopreventive Effects in Different Cancer Cell Lines. International Journal of Molecular Sciences. 2024; 25(14):7689. https://doi.org/10.3390/ijms25147689
Chicago/Turabian StyleUrakawa, Daigo, Yuki Shioiridani, Shinya Igata, De-Xing Hou, and Kozue Sakao. 2024. "Comparative Analysis of Acetylated Flavonoids’ Chemopreventive Effects in Different Cancer Cell Lines" International Journal of Molecular Sciences 25, no. 14: 7689. https://doi.org/10.3390/ijms25147689