
Saturation Mutagenesis and Molecular Modeling: The Impact of Methionine 182 Substitutions on the Stability of β-Lactamase TEM-1

 , ,
, ,
Abstract
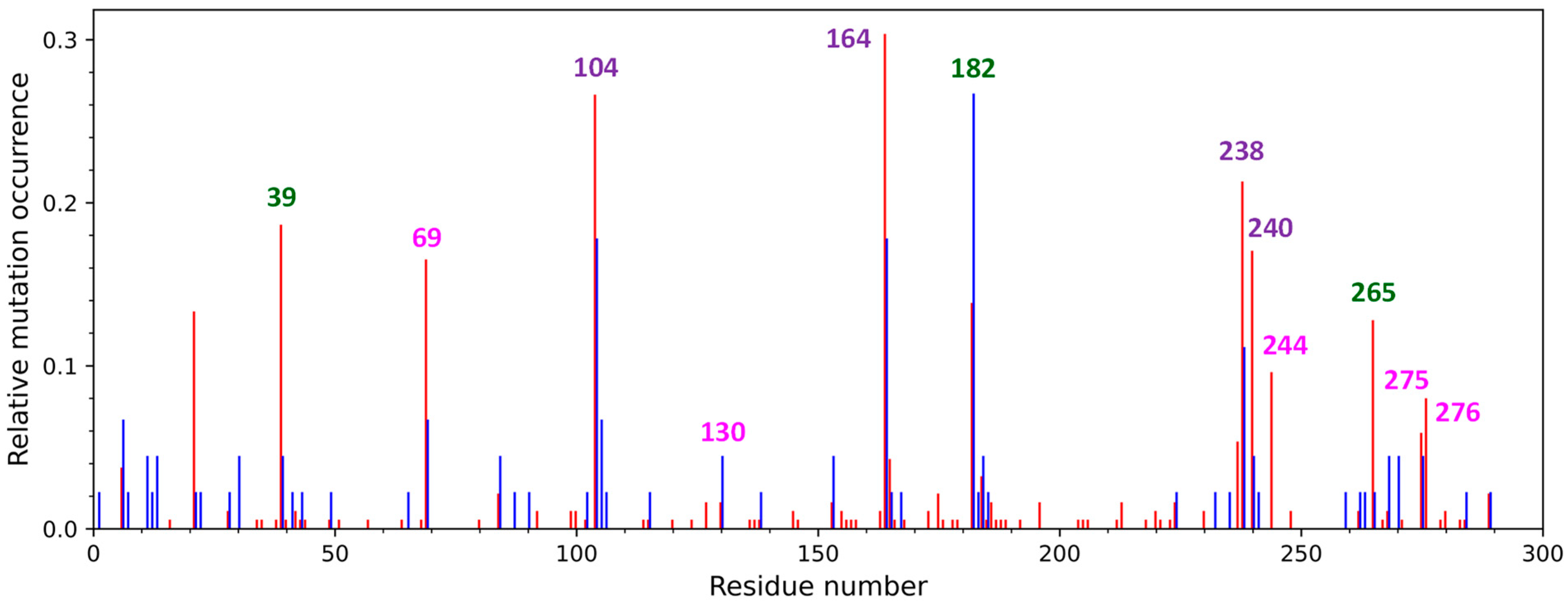
1. Introduction
2. Results
2.1. Periplasmic Expression of TEM Type β-Lactamase Genes in E. coli Cells
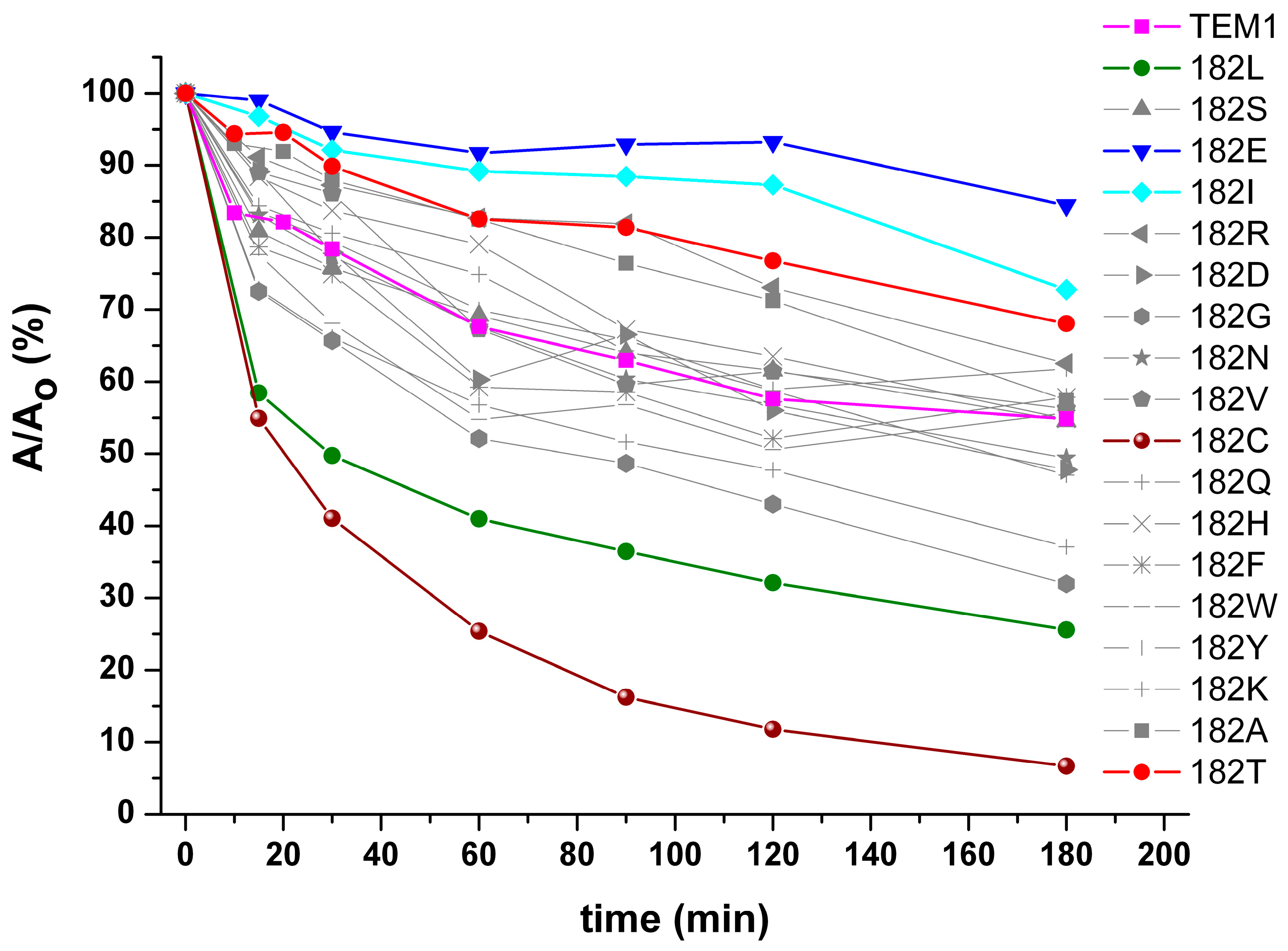
2.2. Determination of Catalytic Parameters and Thermostability of Recombinant TEM-Type β-Lactamases with M182 Substitutions
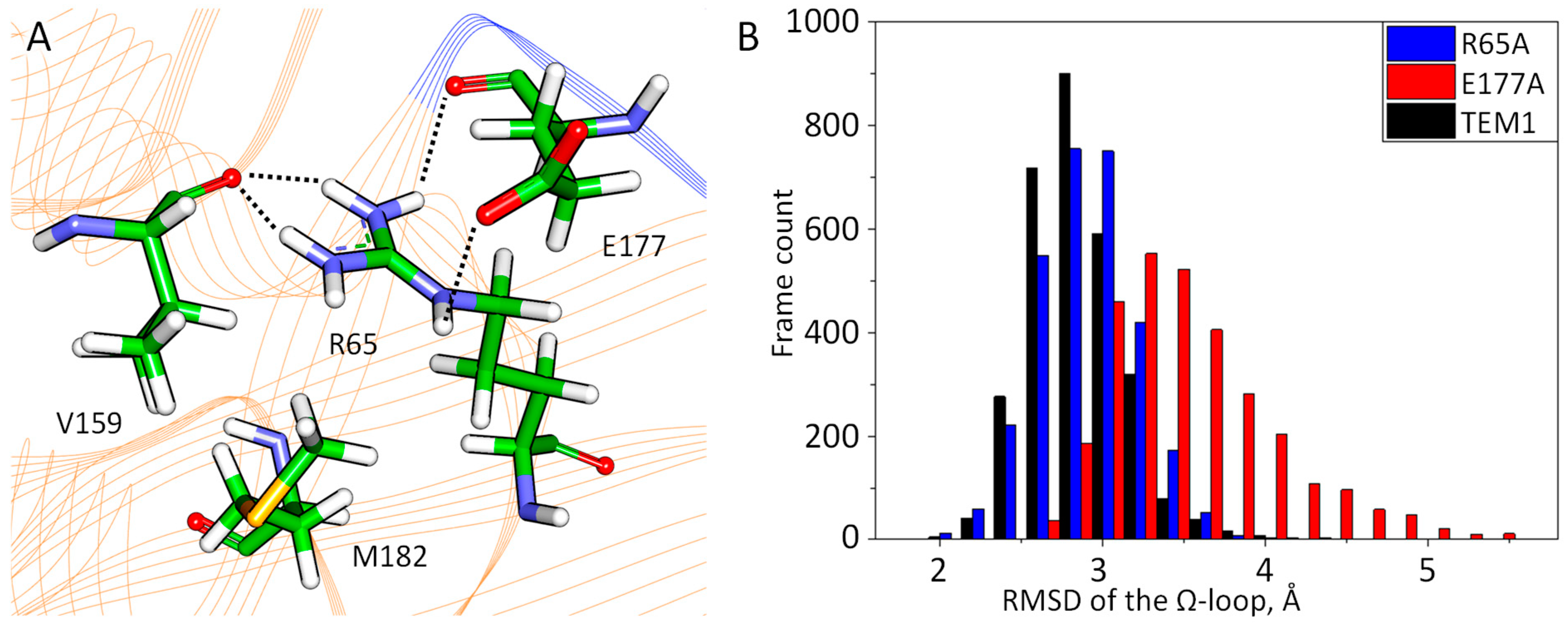
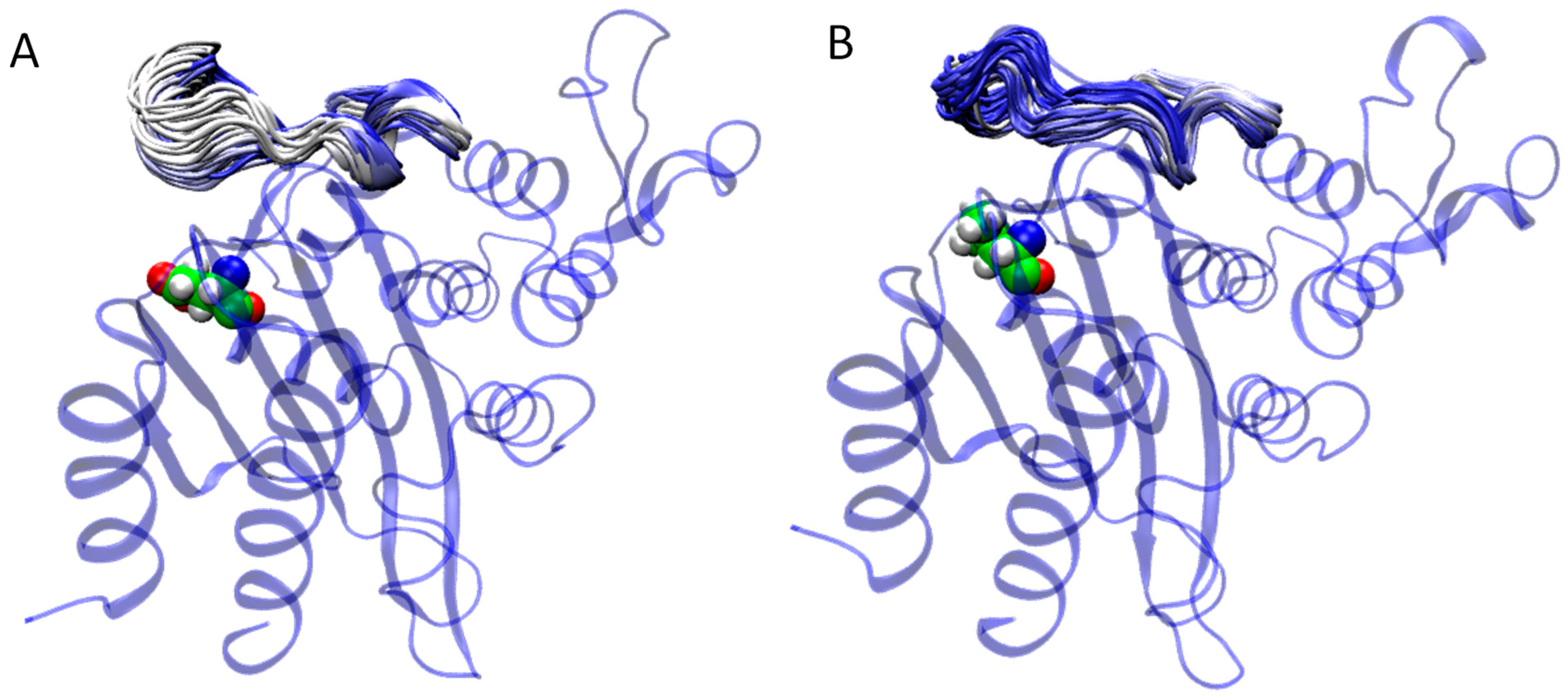
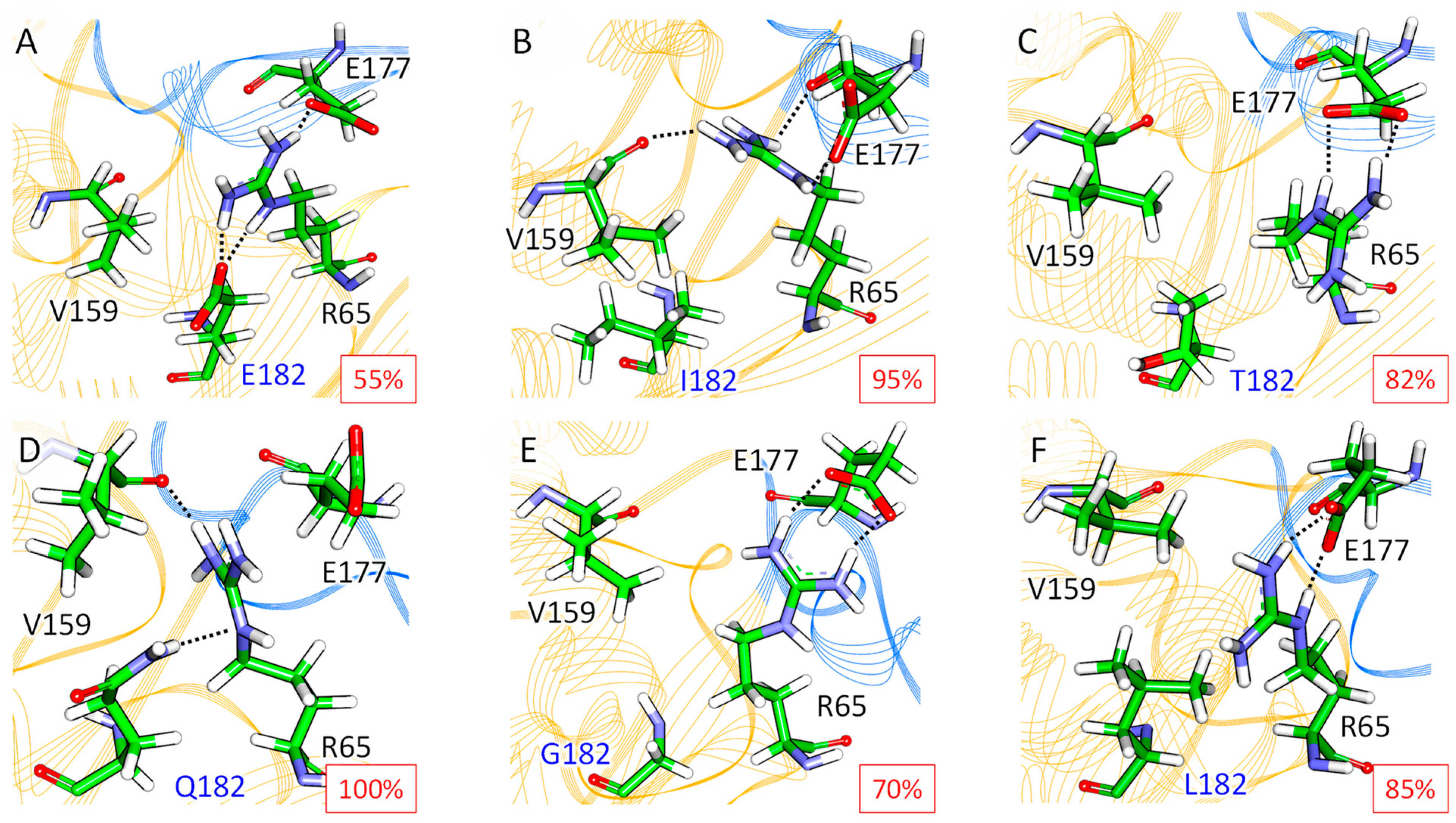
2.3. Molecular Modeling of the Influence of Substitutions at M182 Residue
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Microorganisms, Media, Plasmids, and Oligonucleotides
4.3. Introduction of Mutations into blaTEM-1 Using the Quick Change Method
4.4. Cultivation of E. coli Cells
4.5. Isolation and Purification of Recombinant Mutant Forms of TEM-Type β-Lactamases
4.6. Determination of Catalytic Parameters of Recombinant ß-Lactamases
4.7. The Study of the Thermal Inactivation of Recombinant ß-Lactamases
4.8. Thermal Shift Analysis of Recombinant ß-Lactamases
4.9. Molecular Modeling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eichenberger, E.M.; Thaden, J.T. Epidemiology and Mechanisms of Resistance of Extensively Drug Resistant Gram-Negative Bacteria. Antibiotics 2019, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.P.; Chain, E. An Enzyme from Bacteria able to Destroy Penicillin. Nature 1940, 146, 837. [Google Scholar] [CrossRef]
- Bush, K. Classification for β-lactamases: Historical perspectives. Expert Rev. Anti. Infect. Ther. 2023, 21, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, R.A. β-Lactamases: A Focus on Current Challenges. Cold Spring Harb. Perspect. Med. 2017, 7, a025239. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, G.; Del Giacomo, P.; Posteraro, B.; Sanguinetti, M.; Tumbarello, M. Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae. Int. J. Mol. Sci. 2020, 21, 5090. [Google Scholar] [CrossRef] [PubMed]
- Ogawara, H. Comparison of Antibiotic Resistance Mechanisms in Antibiotic-Producing and Pathogenic Bacteria. Molecules 2019, 24, 3430. [Google Scholar] [CrossRef]
- Årdal, C.; Balasegaram, M.; Laxminarayan, R.; McAdams, D.; Outterson, K.; Rex, J.H.; Sumpradit, N. Antibiotic development—Economic, regulatory and societal challenges. Nat. Rev. Microbiol. 2020, 18, 267–274. [Google Scholar] [CrossRef]
- Datta, N.; Kontomichalou, P. Penicillinase Synthesis Controlled By Infectious R Factors In Enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef]
- Jacquier, H.; Birgy, A.; Le Nagard, H.; Mechulam, Y.; Schmitt, E.; Glodt, J.; Bercot, B.; Petit, E.; Poulain, J.; Barnaud, G.; et al. Capturing the mutational landscape of the beta-lactamase TEM-1. Proc. Natl. Acad. Sci. USA 2013, 110, 13067–13072. [Google Scholar] [CrossRef]
- Salverda, M.L.M.; De Visser, J.A.G.M.; Barlow, M. Natural evolution of TEM-1 β-lactamase: Experimental reconstruction and clinical relevance. FEMS Microbiol. Rev. 2010, 34, 1015–1036. [Google Scholar] [CrossRef] [PubMed]
- Birgy, A.; Magnan, M.; Hobson, C.A.; Figliuzzi, M.; Panigoni, K.; Codde, C.; Tenaillon, O.; Jacquier, H. Local and Global Protein Interactions Contribute to Residue Entrenchment in Beta-Lactamase TEM-1. Antibiotics 2022, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, V.G.; Rubtsova, M.Y.; Uporov, I.V.; Ishtubaev, I.V.; Andreeva, I.P.; Shcherbinin, D.S.; Veselovsky, A.V.; Egorov, A.M. Bacterial TEM-Type Serine Beta-Lactamases: Structure and Analysis of Mutations. Biochem. Suppl. Ser. B Biomed. Chem. 2018, 12, 87–95. [Google Scholar] [CrossRef]
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-spectrum β -lactamases: An update on their characteristics, epidemiology and detection. JAC-Antimicrob. Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef] [PubMed]
- Lepe, J.A.; Martínez-Martínez, L. Resistance mechanisms in Gram-negative bacteria. Med. Intensiv. 2022, 46, 392–402. [Google Scholar] [CrossRef]
- Egorov, A.; Rubtsova, M.; Grigorenko, V.; Uporov, I.; Veselovsky, A. The Role of the Ω-Loop in Regulation of the Catalytic Activity of TEM-Type β-Lactamases. Biomolecules 2019, 9, 854. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Okoye, C.N.; Biesbrock, D.; Harris, E.C.; Miyasaki, K.F.; Rilinger, R.G.; Tso, M.; Hart, K.M. Natural and Synthetic Suppressor Mutations Defy Stability–Activity Tradeoffs. Biochemistry 2022, 61, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Minasov, G.; Shoichet, B.K. Evolution of an Antibiotic Resistance Enzyme Constrained by Stability and Activity Trade-offs. J. Mol. Biol. 2002, 320, 85–95. [Google Scholar] [CrossRef]
- Sideraki, V.; Huang, W.; Palzkill, T.; Gilbert, H.F. A secondary drug resistance mutation of TEM-1 β-lactamase that suppresses misfolding and aggregation. Proc. Natl. Acad. Sci. USA 2001, 98, 283–288. [Google Scholar] [CrossRef]
- Docquier, J.-D.; Benvenuti, M.; Calderone, V.; Rossolini, G.-M.; Mangani, S. Structure of the extended-spectrum β-lactamase TEM-72 inhibited by citrate. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 303–306. [Google Scholar] [CrossRef]
- Farzaneh, S.; Chaibi, E.B.; Peduzzi, J.; Barthelemy, M.; Labia, R.; Blazquez, J.; Baquero, F. Implication of Ile-69 and Thr-182 residues in kinetic characteristics of IRT-3 (TEM-32) beta-lactamase. Antimicrob. Agents Chemother. 1996, 40, 2434–2436. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, V.G.; Andreeva, I.P.; Serova, O.V.; Rubtsova, M.Y.; Egorov, A.M. Impact of Residue R65 on the Stabilization of TEM-Type β-Lactamases with Substitution of M182T. Moscow Univ. Chem. Bull. 2020, 75, 179–185. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-Lactamase Data Base (BLDB)—Structure and function. J. Enzyme Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef] [PubMed]
- Shcherbinin, D.; Veselovsky, A.; Rubtsova, M.; Grigorenko, V.; Egorov, A. The impact of long-distance mutations on the Ω-loop conformation in TEM type β-lactamases. J. Biomol. Struct. Dyn. 2020, 38, 2369–2376. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, M.; Raquet, X.; Palzkill, T.; Pain, R.H.; Frère, J.-M. The rate-limiting step in the folding of the cis -Pro167Thr mutant of TEM-1 β-lactamase is the trans to cis isomerization of a non-proline peptide bond. Proteins Struct. Funct. Bioinforma. 1996, 25, 104–111. [Google Scholar] [CrossRef]
- Egorov, A.M.; Ulyashova, M.M.; Rubtsova, M.Y. Bacterial Enzymes and Antibiotic Resistance. Acta Naturae 2018, 10, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, M.; Lejeune, A.; Pain, R.H. β-Lactamases as models for protein-folding studies. Cell. Mol. Life Sci. CMLS 1998, 54, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, M.; Raquet, X.; Frère, J. Investigation of the folding pathway of the TEM-1 β-lactamase. Proteins Struct. Funct. Bioinforma. 1995, 22, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, V.G.; Andreeva, I.P.; Rubtsova, M.Y.; Deygen, I.M.; Antipin, R.L.; Majouga, A.G.; Egorov, A.M.; Beshnova, D.A.; Kallio, J.; Hackenberg, C.; et al. Novel non-β-lactam inhibitor of β-lactamase TEM-171 based on acylated phenoxyaniline. Biochimie 2017, 132, 45–53. [Google Scholar] [CrossRef]
- Bebrone, C.; Moali, C.; Mahy, F.; Rival, S.; Docquier, J.D.; Rossolini, G.M.; Fastrez, J.; Pratt, R.F.; Frère, J.-M.; Galleni, M. CENTA as a Chromogenic Substrate for Studying β-Lactamases. Antimicrob. Agents Chemother. 2001, 45, 1868–1871. [Google Scholar] [CrossRef]
- Cantu, C.; Huang, W.; Palzkill, T. Cephalosporin Substrate Specificity Determinants of TEM-1 β-Lactamase. J. Biol. Chem. 1997, 272, 29144–29150. [Google Scholar] [CrossRef] [PubMed]
- Stojanoski, V.; Chow, D.-C.; Hu, L.; Sankaran, B.; Gilbert, H.F.; Prasad, B.V.V.; Palzkill, T. A Triple Mutant in the Ω-loop of TEM-1 β-Lactamase Changes the Substrate Profile via a Large Conformational Change and an Altered General Base for Catalysis. J. Biol. Chem. 2015, 290, 10382–10394. [Google Scholar] [CrossRef]
- Minasov, G.; Wang, X.; Shoichet, B.K. An Ultrahigh Resolution Structure of TEM-1 β-Lactamase Suggests a Role for Glu166 as the General Base in Acylation. J. Am. Chem. Soc. 2002, 124, 5333–5340. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Denning, E.J.; Priyakumar, U.D.; Nilsson, L.; Mackerell, A.D. Impact of 2′-hydroxyl sampling on the conformational properties of RNA: Update of the CHARMM all-atom additive force field for RNA. J. Comput. Chem. 2011, 32, 1929–1943. [Google Scholar] [CrossRef] [PubMed]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; MacKerell, A.D. Optimization of the Additive CHARMM All-Atom Protein Force Field Targeting Improved Sampling of the Backbone ϕ, ψ and Side-Chain χ1 and χ2 Dihedral Angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M182 Substitution in β-Lactamase TEM-1 | Tm, °C | ∆T, °C | Relative Residual Activity, % |
|---|---|---|---|
| M182 (wild-type) | 48.9 ± 0.2 | 0 | 55 |
| M182T | 56.2 ± 0.2 | 7.3 | 68 |
| M182S | 53.6 ± 0.2 | 4.7 | 55 |
| M182A | 50.5 ± 0.1 | 1.6 | 57 |
| M182C | 50.6 ± 0.1 | 1.7 | 7 |
| M182Q | 49.1 ± 0.1 | 0.2 | 47 |
| M182N | 49.1 ± 0.2 | 0.2 | 49 |
| M182H | 48.6 ± 0.1 | −0.3 | 55 |
| M182V | 48.1 ± 0.2 | −0.8 | 56 |
| M182E | 47.0 ± 0.1 | −1.9 | 84 |
| M182K | 46.6 ± 0.1 | −2.3 | 37 |
| M182W | 46.2 ± 0.1 | −2.7 | 56 |
| M182Y | 45.7 ± 0.2 | −3.2 | 62 |
| M182I | 45.6 ± 0.1 | −3.3 | 73 |
| M182G | 45.0 ± 0.1 | −3.9 | 32 |
| M182R | 44.2 ± 0.1 | −4.7 | 63 |
| M182F | 44.1 ± 0.1 | −4.8 | 58 |
| M182D | 42.3 ± 0.1 | −6.6 | 48 |
| M182L | 41.1 ± 0.1 | −7.8 | 26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grigorenko, V.G.; Krivitskaya, A.V.; Khrenova, M.G.; Rubtsova, M.Y.; Presnova, G.V.; Andreeva, I.P.; Serova, O.V.; Egorov, A.M. Saturation Mutagenesis and Molecular Modeling: The Impact of Methionine 182 Substitutions on the Stability of β-Lactamase TEM-1. Int. J. Mol. Sci. 2024, 25, 7691. https://doi.org/10.3390/ijms25147691
Grigorenko VG, Krivitskaya AV, Khrenova MG, Rubtsova MY, Presnova GV, Andreeva IP, Serova OV, Egorov AM. Saturation Mutagenesis and Molecular Modeling: The Impact of Methionine 182 Substitutions on the Stability of β-Lactamase TEM-1. International Journal of Molecular Sciences. 2024; 25(14):7691. https://doi.org/10.3390/ijms25147691
Chicago/Turabian StyleGrigorenko, Vitaly G., Alexandra V. Krivitskaya, Maria G. Khrenova, Maya Yu. Rubtsova, Galina V. Presnova, Irina P. Andreeva, Oxana V. Serova, and Alexey M. Egorov. 2024. "Saturation Mutagenesis and Molecular Modeling: The Impact of Methionine 182 Substitutions on the Stability of β-Lactamase TEM-1" International Journal of Molecular Sciences 25, no. 14: 7691. https://doi.org/10.3390/ijms25147691
APA StyleGrigorenko, V. G., Krivitskaya, A. V., Khrenova, M. G., Rubtsova, M. Y., Presnova, G. V., Andreeva, I. P., Serova, O. V., & Egorov, A. M. (2024). Saturation Mutagenesis and Molecular Modeling: The Impact of Methionine 182 Substitutions on the Stability of β-Lactamase TEM-1. International Journal of Molecular Sciences, 25(14), 7691. https://doi.org/10.3390/ijms25147691