
Testosterone Nanoemulsion Prevents Prostate Cancer: PC-3 and LNCaP Cell Viability In Vitro


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
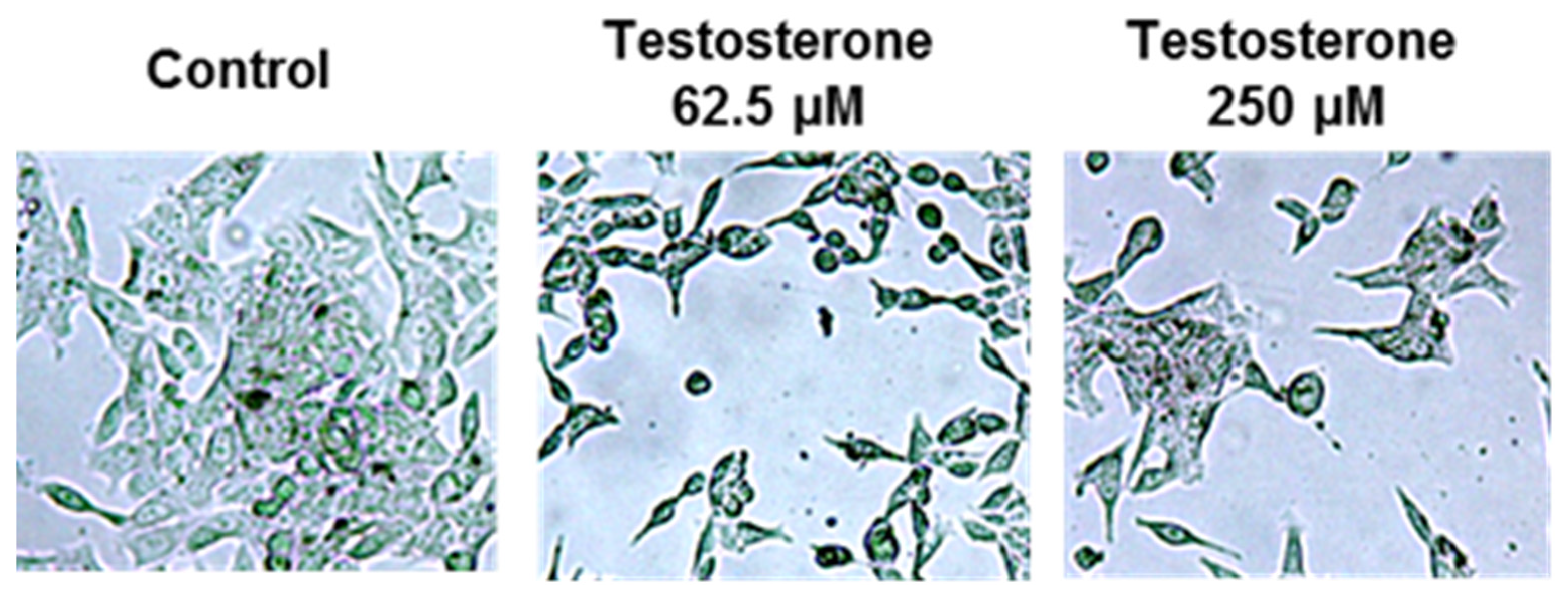
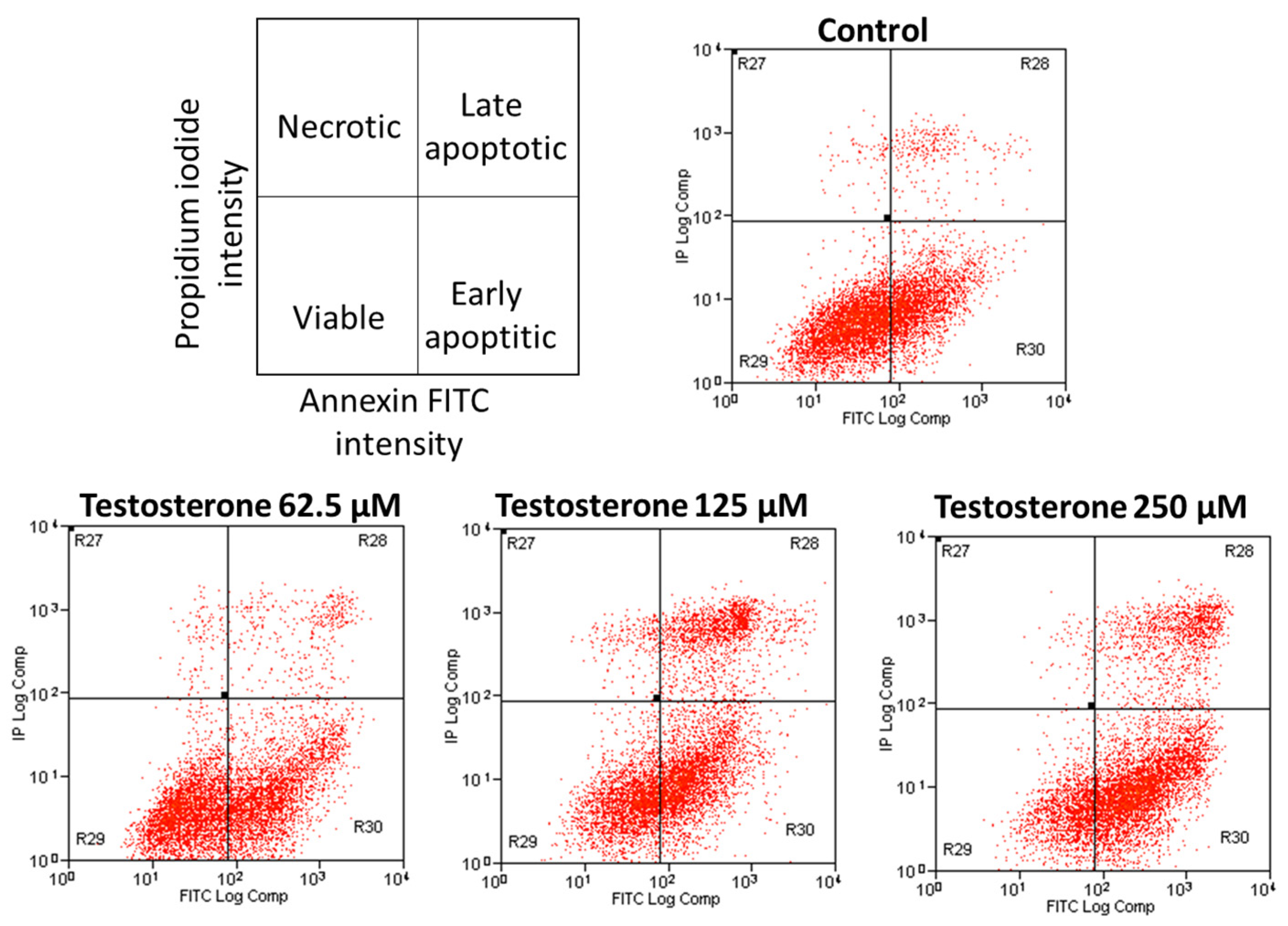
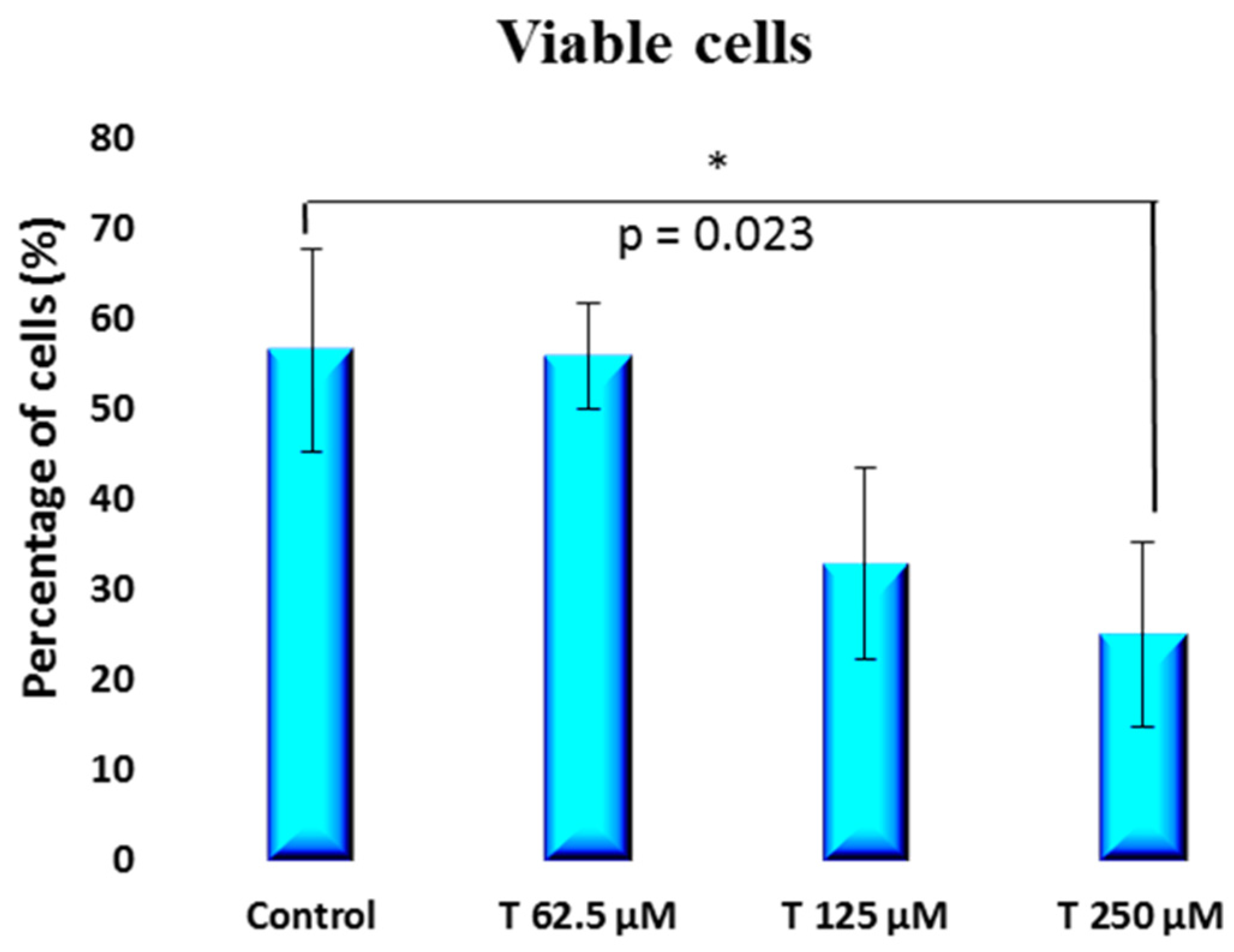
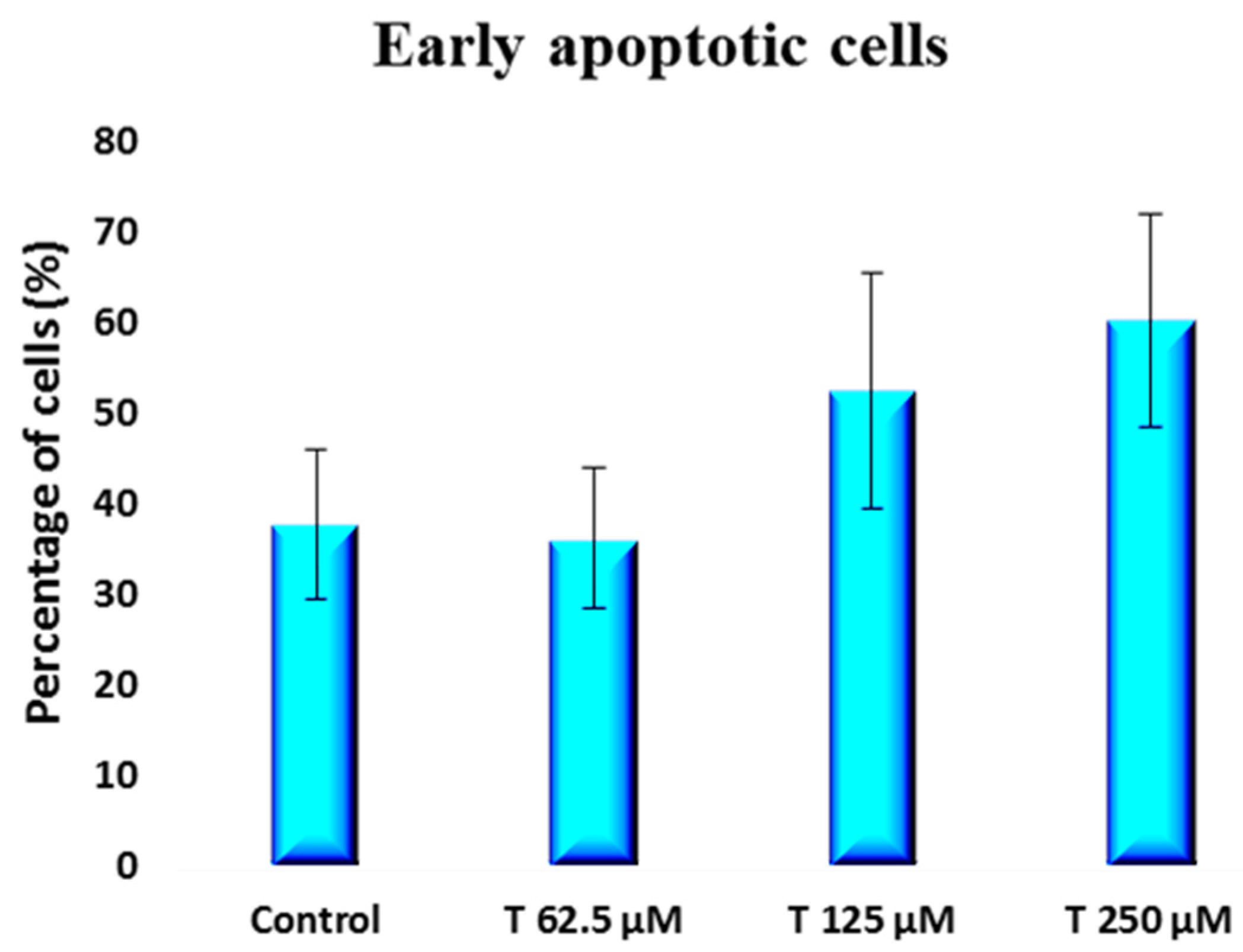
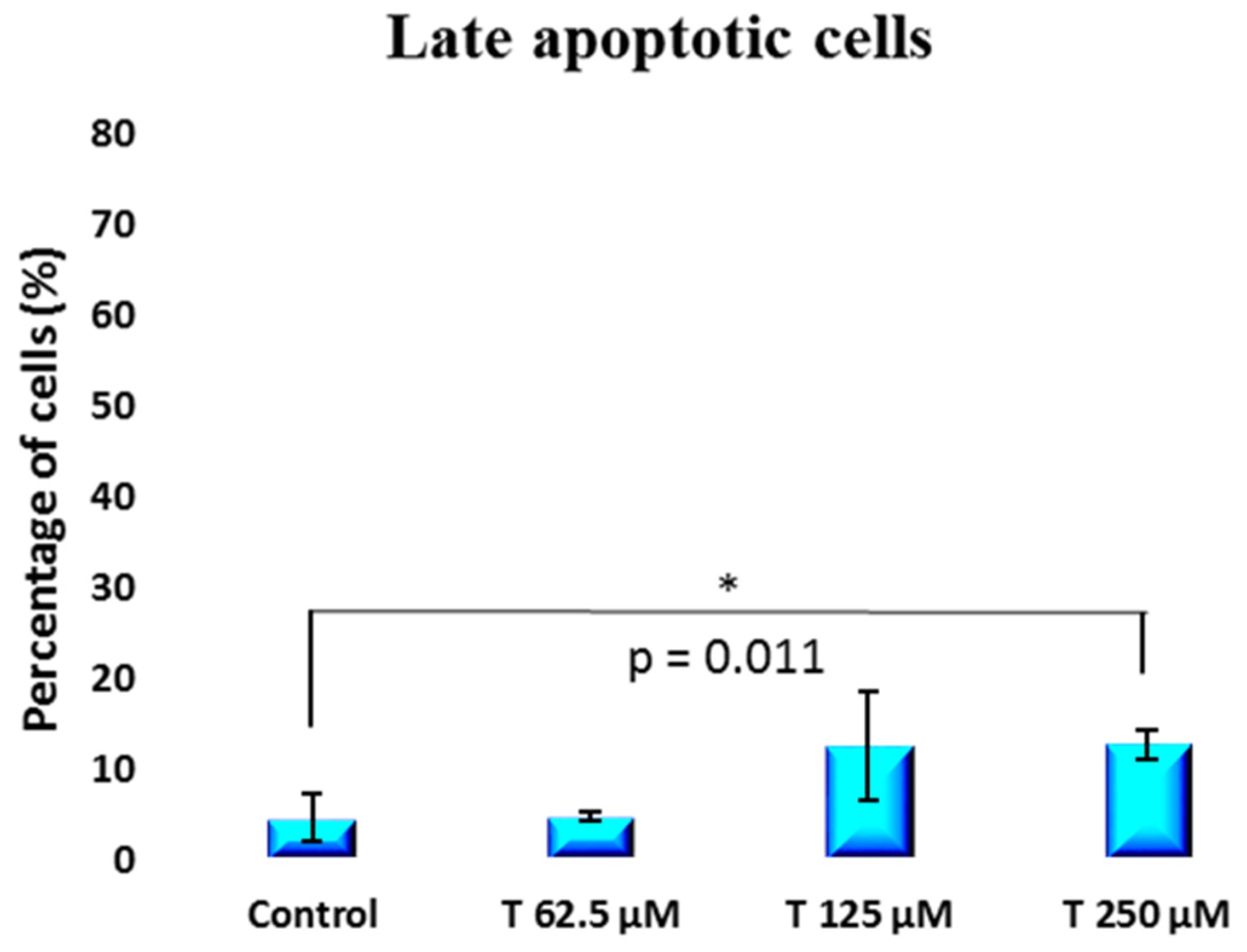
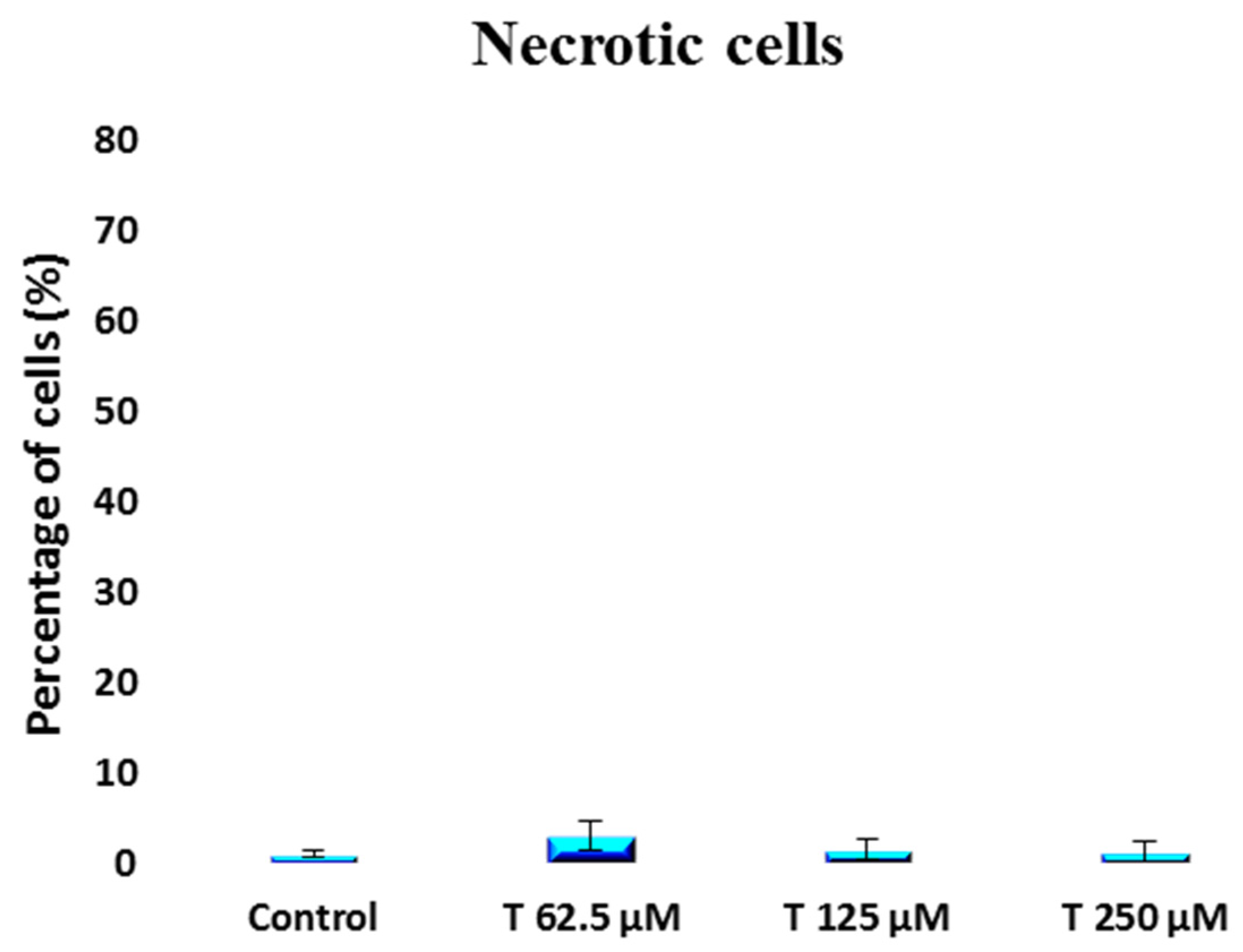
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Metabolic Activity Evaluation—Alamar Blue Test
4.3. Cell Viability Evaluation—Flow Cytometry
4.4. Particle Size Z-Average and Physical Stability
4.5. Nanoemulsion Preparation
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Cancer Society. Cancer Statistics Center. Available online: https://cancerstatisticscenter.cancer.org/#/cancer-site/Prostate (accessed on 25 August 2017).
- Stepan, J.J.; Lachman, M.; Zverina, J.; Pacovsky, V.; Baylink, D.J. Castrated men exhibit bone loss: Effect of calcitonin treatment on biochemical indices of bone remodeling. J. Clin. Endocrinol. Metab. 1989, 69, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Scimeca, M.; Bonfiglio, R.; Varone, F.; Ciuffa, S.; Mauriello, A.; Bonanno, E. Calcifications in prostate cancer: An active phenomenon mediated by epithelial cells with osteoblast-phenotype. Microsc. Res. Tech. 2018, 81, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Szulc, P.; Hofbauer, L.C.; Heufelder, A.E.; Roth, S.; Delmas, P.D. Osteoprotegerin serum levels in men: Correlation with age, estrogen, and testosterone status. J. Clin. Endocrinol. Metab. 2001, 86, 3162–3165. [Google Scholar] [CrossRef] [PubMed]
- Mearini, L.; Zucchi, A.; Nunzi, E.; Villirillo, T.; Bini, V.; Porena, M. Low serum testosterone levels are predictive of prostate cancer. World J. Urol. 2013, 31, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Morgentaler, A. Testosterone replacement therapy and prostate cancer. Urol. Clin. N. Am. 2007, 34, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Debras, C.; Chazelas, E.; Srour, B.; Kesse-Guyot, E.; Julia, C.; Zelek, L.; Agaësse, C.; Druesne-Pecollo, N.; Galan, P.; Hercberg, S.; et al. Total and added sugar intakes, sugar types, and cancer risk: Results from the prospective NutriNet-Santé cohort. Am. J. Clin. Nutr. 2020, 112, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.D.; Chen, Y.H.; Xu, S.; Zhang, C.; Wang, D.M.; Wang, H.; Chen, L.; Zhang, Z.H.; Xia, M.Z.; Xu, D.X.; et al. Low vitamin D status is associated with inflammation in patients with prostate cancer. Oncotarget 2017, 8, 22076–22085. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Yang, X.; Chen, H.; Zhang, S.; Chen, X.; Sheng, Y.; Pang, J. Association of cigarette smoking habits with the risk of prostate cancer: A systematic review and meta-analysis. BMC Public Health 2023, 23, 1150. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bandini, M.; Gandaglia, G.; Briganti, A. Obesity and prostate cancer. Curr. Opin. Urol. 2017, 27, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Flores, I.E.; Sierra-Fonseca, J.A.; Davalos, O.; Saenz, L.A.; Castellanos, M.M.; Zavala, J.K.; Gosselink, K.L. Stress alters the expression of cancer-related genes in the prostate. BMC Cancer 2017, 17, 621. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Morales, A.; Siemens, D.R. Testosterone Therapy and Prostate Cancer: Incorporating Low-Level Evidence into Practical Recommendations. Urol. Clin. N. Am. 2022, 49, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Morgentaler, A.; Caliber, M. Safety of testosterone therapy in men with prostate cancer. Expert Opin. Drug Saf. 2019, 18, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Bosland, M.C.; Schlicht, M.J.; Horton, L.; McCormick, D.L. The MNU Plus Testosterone Rat Model of Prostate Carcinogenesis. Toxicol. Pathol. 2022, 50, 478–496. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Botelho, M.A.; Martins, J.G.; Ruela, R.S. Nanotechnology in ligature-induced periodontitis: Protective effect of a doxycycline gel with nanoparticules. J. Appl. Oral Sci. 2010, 18, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Gonzaga, L.W.; Botelho, M.A.; Queiroz, D.B. Nanotechnology in Hormone Replacement Therapy: Safe and Efficacy of Transdermal Estriol and Estradiol Nanoparticles after 5 Years Follow-Up Study. Lat. Am. J. Pharm. 2012, 31, 442–450. [Google Scholar]
- Botelho, M.A.; Barros, G.; Queiroz, D.B. Nanotechnology in Phytotherapy: Antiinflammatory Effect of a Nanostructured Thymol Gel from Lippia sidoides in Acute Periodontitis in Rats. Phytother. Res. 2016, 30, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.A.; Queiroz, D.B.; Barros, G. Nanostructured transdermal hormone replacement therapy for relieving menopausal symptoms: A confocal Raman spectroscopy study. Clinics 2014, 69, 75–82. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Botelho, M.A.; Queiroz, D.B.; Freitas, A.; Guerreiro, S.; Umbelino, S.; Barros, G. Effects of a new testosterone transdermal delivery system, Biolipid B2-testosterone in healthy middle aged men: A Confocal Raman Spectroscopy Study. J. Pharm. Sci. Innov. 2013, 2, 1–7. [Google Scholar] [CrossRef]
- Gleave, M.E.; Hsieh, J.-T.; Wu, H.-C.; von Eschenbach, A.C.; Chung, L.W. Serum prostate specific antigen levels in mice bearing human prostate LNCaP tumors are determined by tumor volume and endocrine and growth factors. Cancer Res. 1992, 52, 1598–1605. [Google Scholar]
- Kaighn, M.; Narayan, K.S.; Ohnuki, Y.; Lechner, J.; Jones, L. Establishment and characterization of a human prostatic carcinoma cell line [PC-3]. Investig. Urol. 1979, 17, 16–23. [Google Scholar]
- Nunez, R. Flow cytometry: Principles and instrumentation. Curr. Issues Mol. Biol. 2001, 3, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, R.; Kutzner, B.; Truong, M.; Sanchez-Dardon, J.; McLean, J. Analysis of radiation-induced apoptosis in human lymphocytes: Flow cytometry using Annexin V and propidium iodide versus the neutral comet assay. Cytom. Part A 2002, 48, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Del Buono, C.; Baldracchini, F.; Talesa, V.; Cottini, E.; Brancadoro, C.; Zucchi, A. Alteration of glyoxalases gene expression in response to testosterone in LNCaP and PC3 human prostate cancer cells. Cancer Biol. Ther. 2007, 6, 1880–1888. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botelho, M.A.; Queiroz, D.B. Testosterone Nanoemulsion Prevents Prostate Cancer: PC-3 and LNCaP Cell Viability In Vitro. Int. J. Mol. Sci. 2024, 25, 7729. https://doi.org/10.3390/ijms25147729
Botelho MA, Queiroz DB. Testosterone Nanoemulsion Prevents Prostate Cancer: PC-3 and LNCaP Cell Viability In Vitro. International Journal of Molecular Sciences. 2024; 25(14):7729. https://doi.org/10.3390/ijms25147729
Chicago/Turabian StyleBotelho, Marco Antonio, and Dinalva Brito Queiroz. 2024. "Testosterone Nanoemulsion Prevents Prostate Cancer: PC-3 and LNCaP Cell Viability In Vitro" International Journal of Molecular Sciences 25, no. 14: 7729. https://doi.org/10.3390/ijms25147729
APA StyleBotelho, M. A., & Queiroz, D. B. (2024). Testosterone Nanoemulsion Prevents Prostate Cancer: PC-3 and LNCaP Cell Viability In Vitro. International Journal of Molecular Sciences, 25(14), 7729. https://doi.org/10.3390/ijms25147729