
The Structural Characterization of a Polysaccharide from the Dried Root of Salvia miltiorrhiza and Its Use as a Vaccine Adjuvant to Induce Humoral and Cellular Immune Responses

Abstract

1. Introduction
2. Results
2.1. Characterization of SMPs
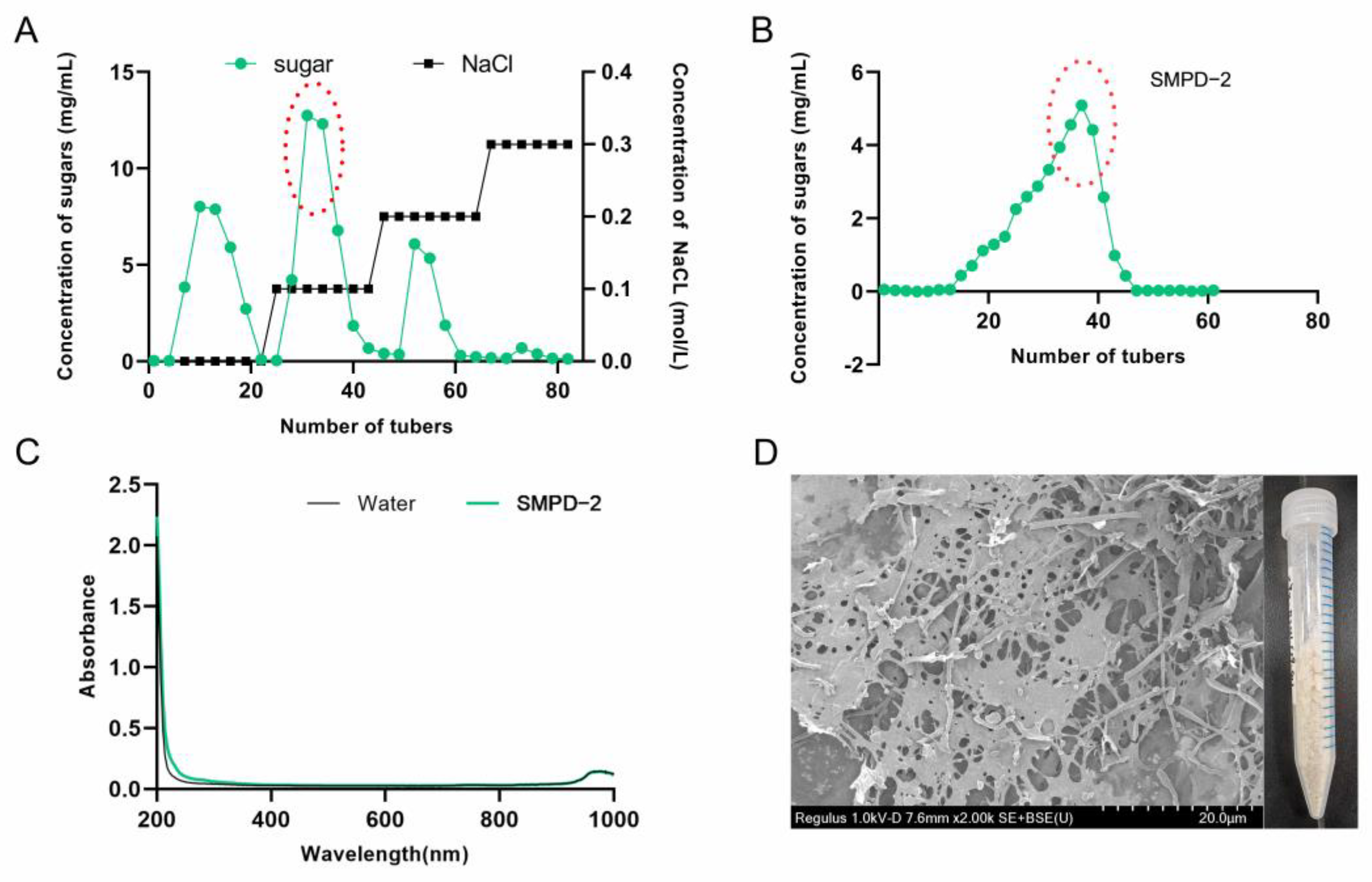
2.1.1. Purification and Morphology
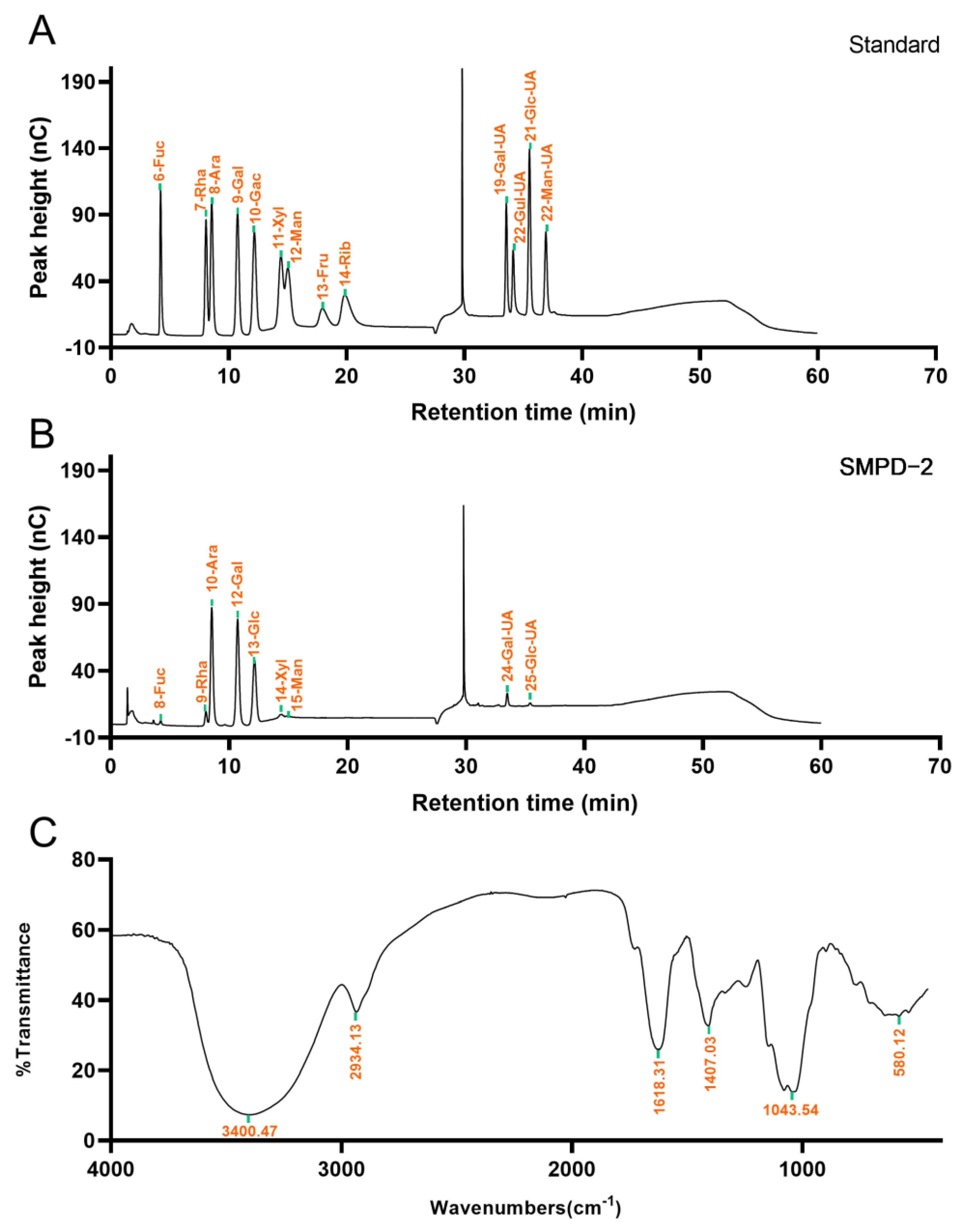
2.1.2. Determination of Monosaccharide Composition and Weight Average Molecular Weight
2.1.3. FT-IR Analysis
2.1.4. Glycosidic Linkage Pattern of SMPD–2
2.1.5. NMR Elucidation of SMPD–2
α-L-Araf-(1→ (Residue A)
→4)-α-D-Galp-(1→ (Residue B)
→3,6)-β-D-Glcp-(1→ (Residue C)
The Connection Mode and Sequence of Sugar Residues
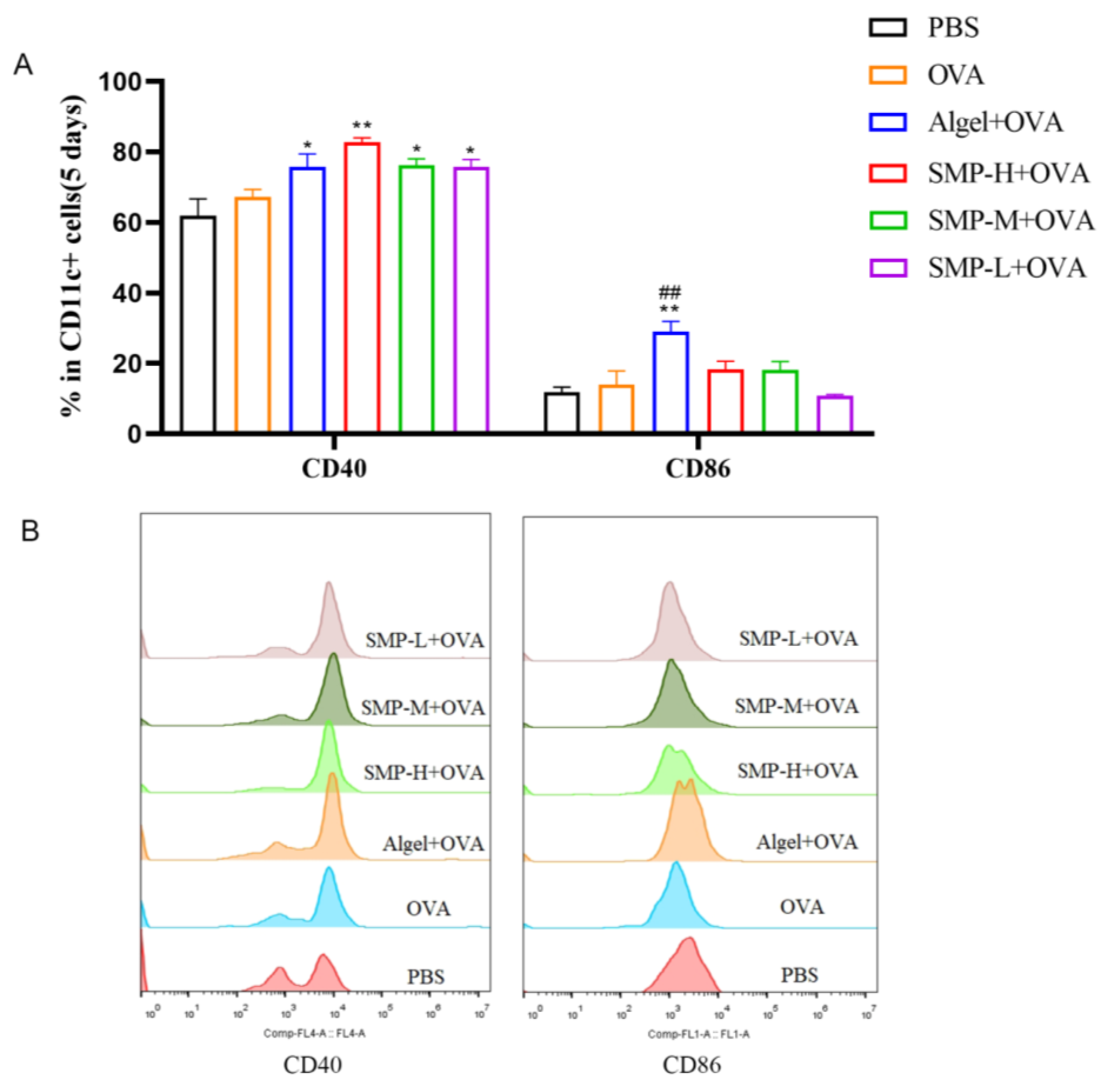
2.2. Activation of Dendritic Cells in Lymph Nodes
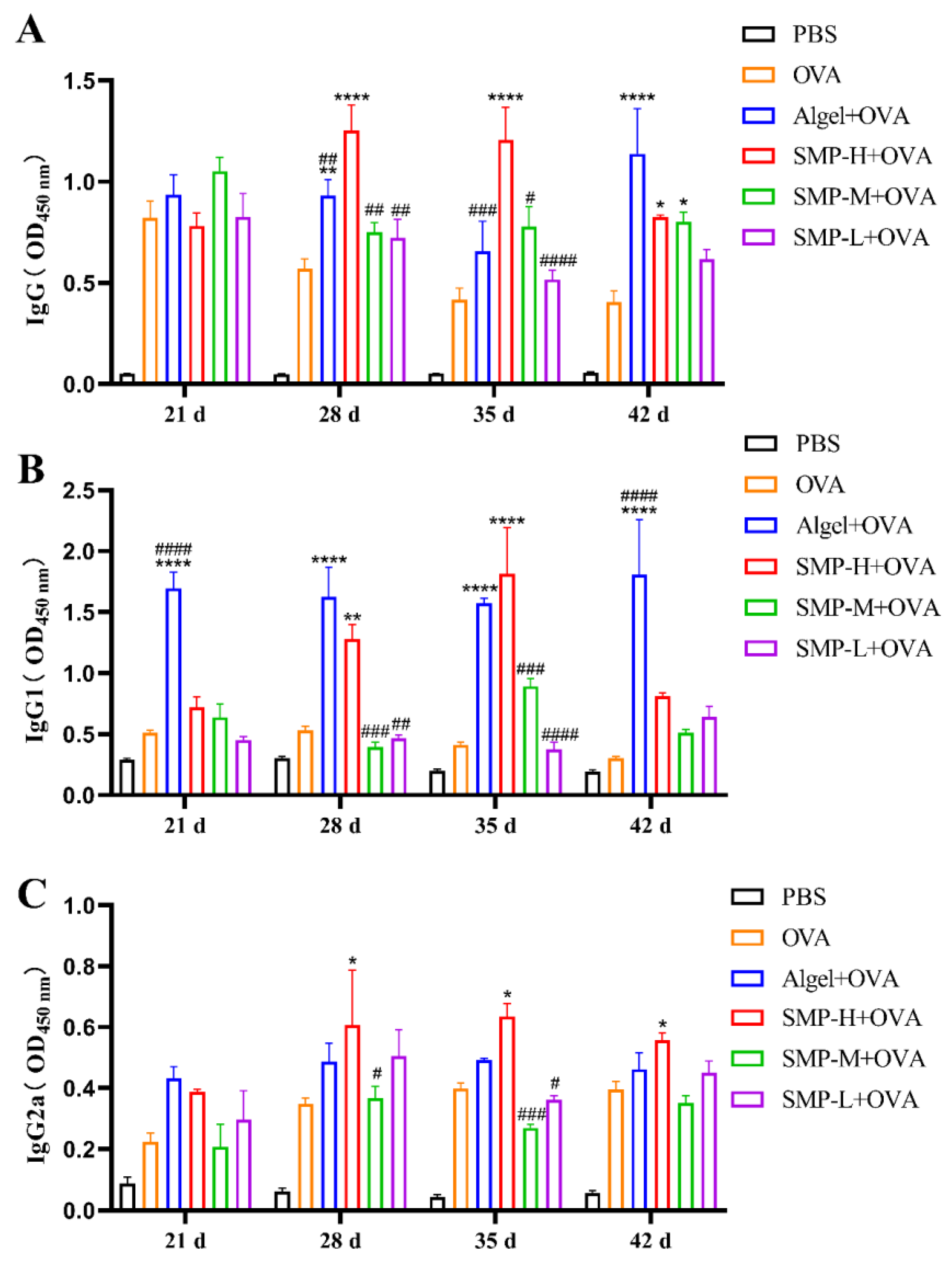
2.3. OVA-Specific IgG and Its Subtypes
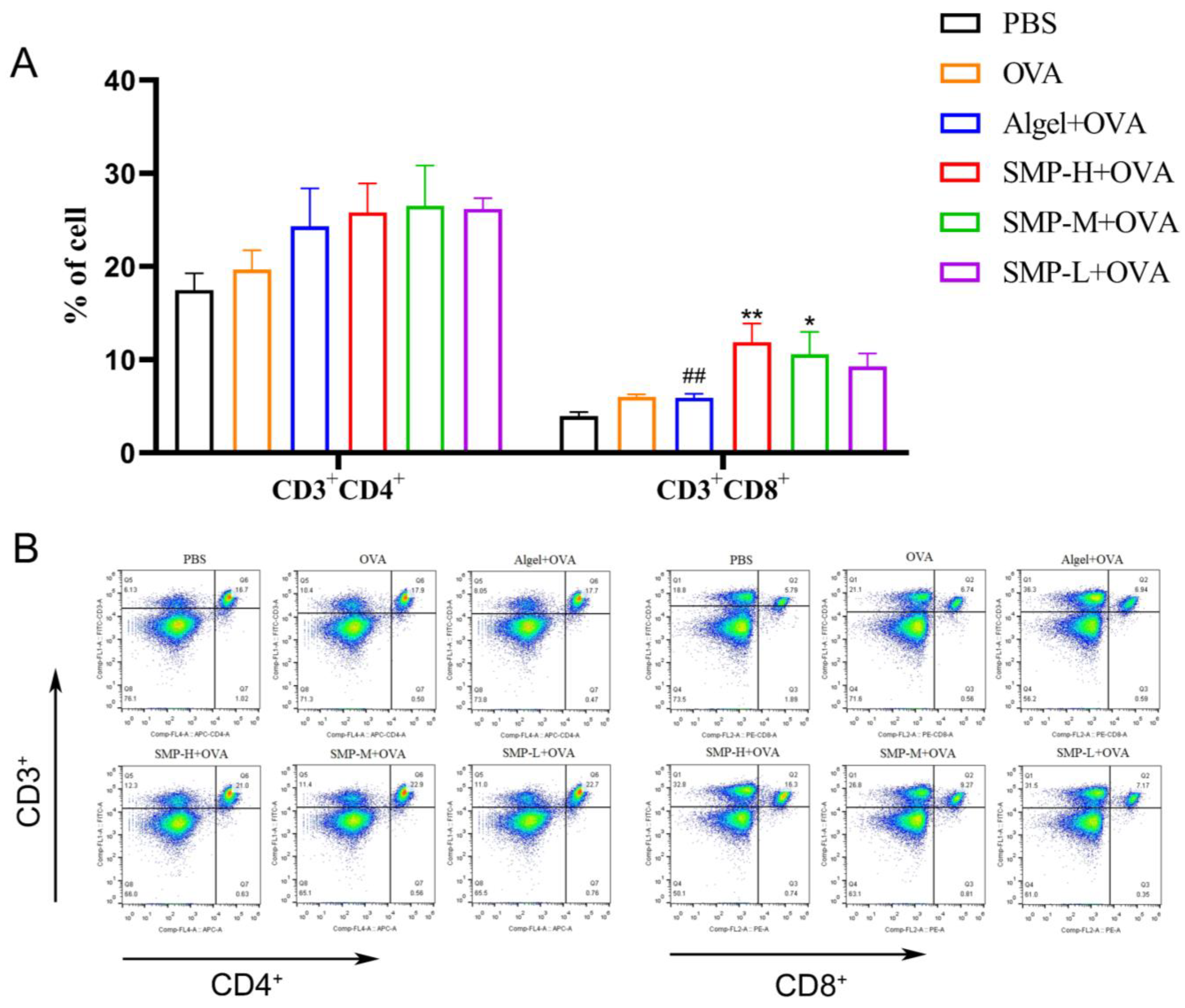
2.4. Activation of Spleen T Cells after Vaccination
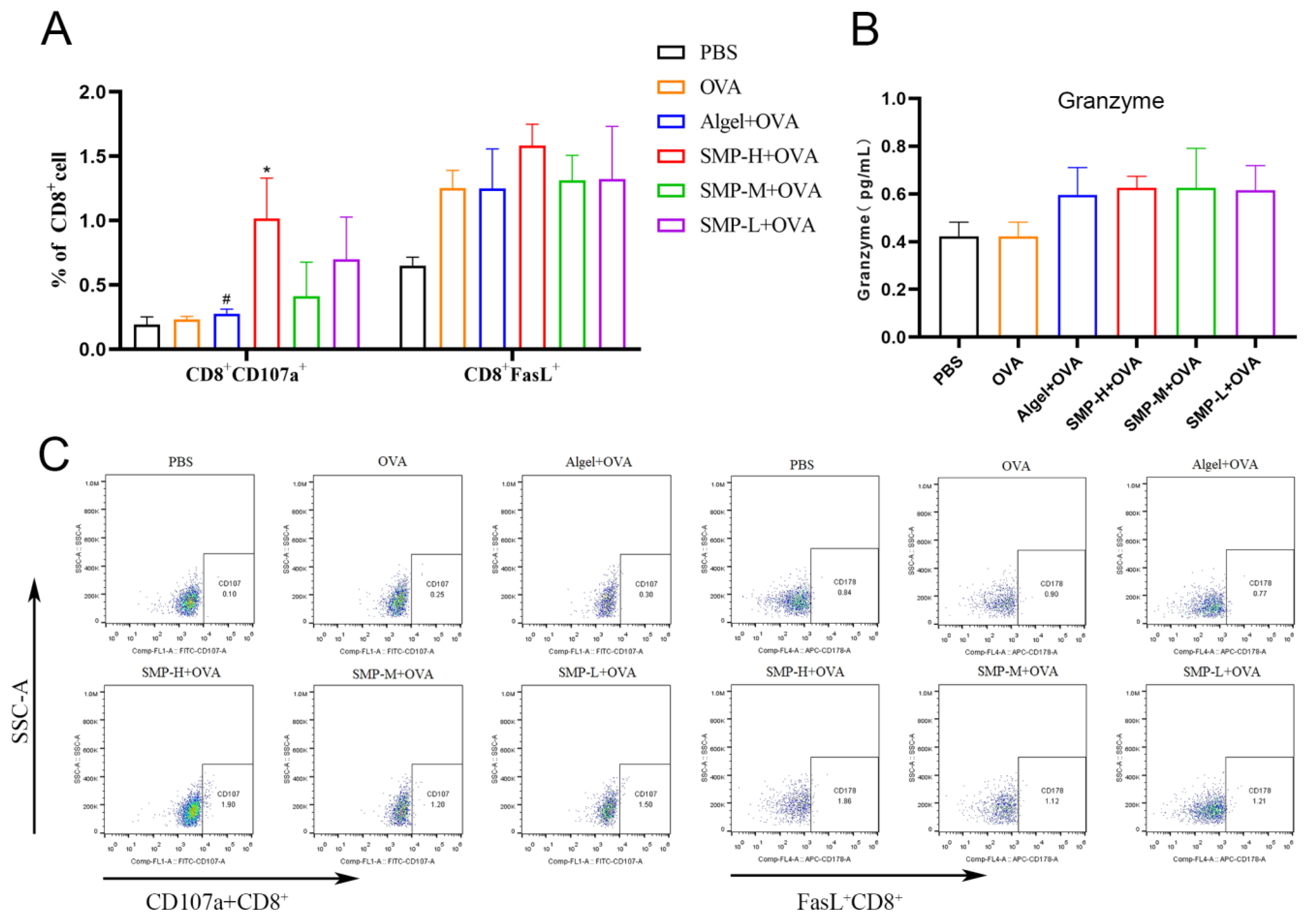
2.5. Activation of Cytotoxic T lymphocytes
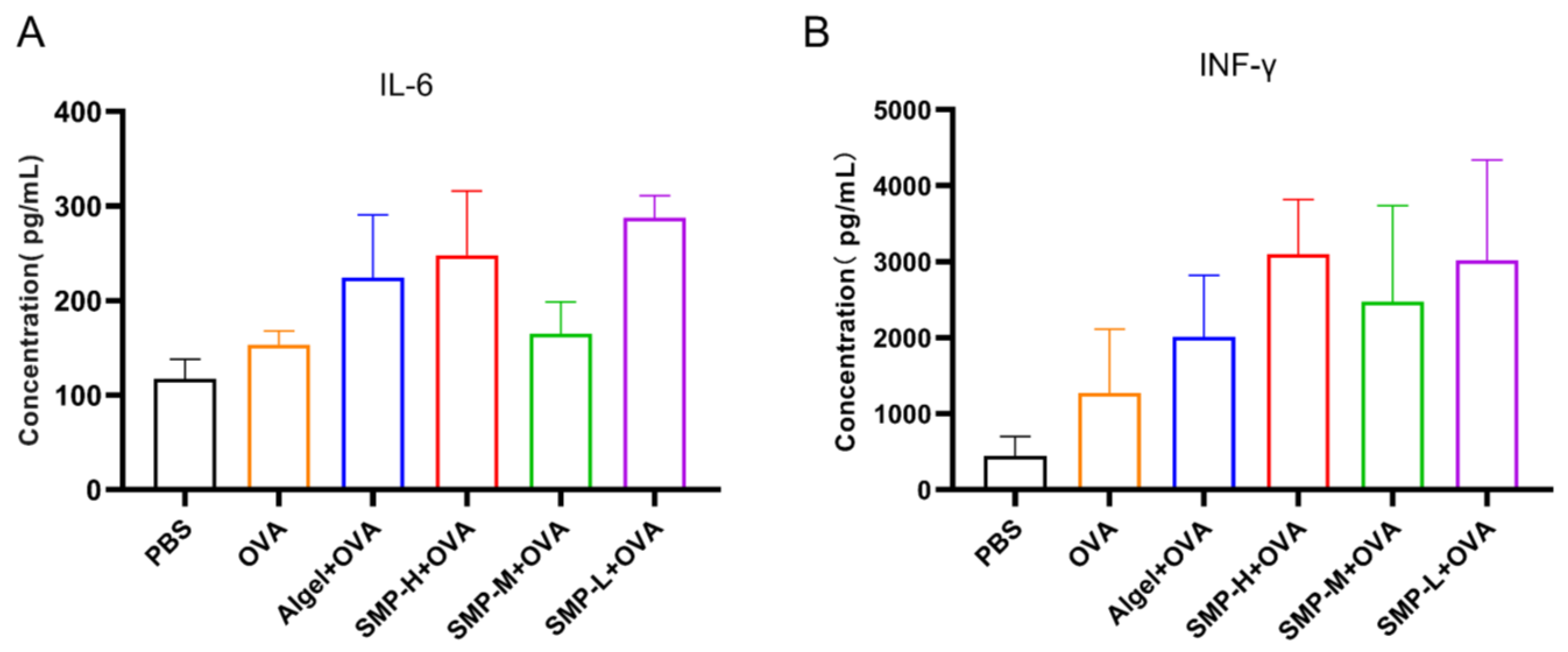
2.6. Cytokine Levels in Cellular Supernatant
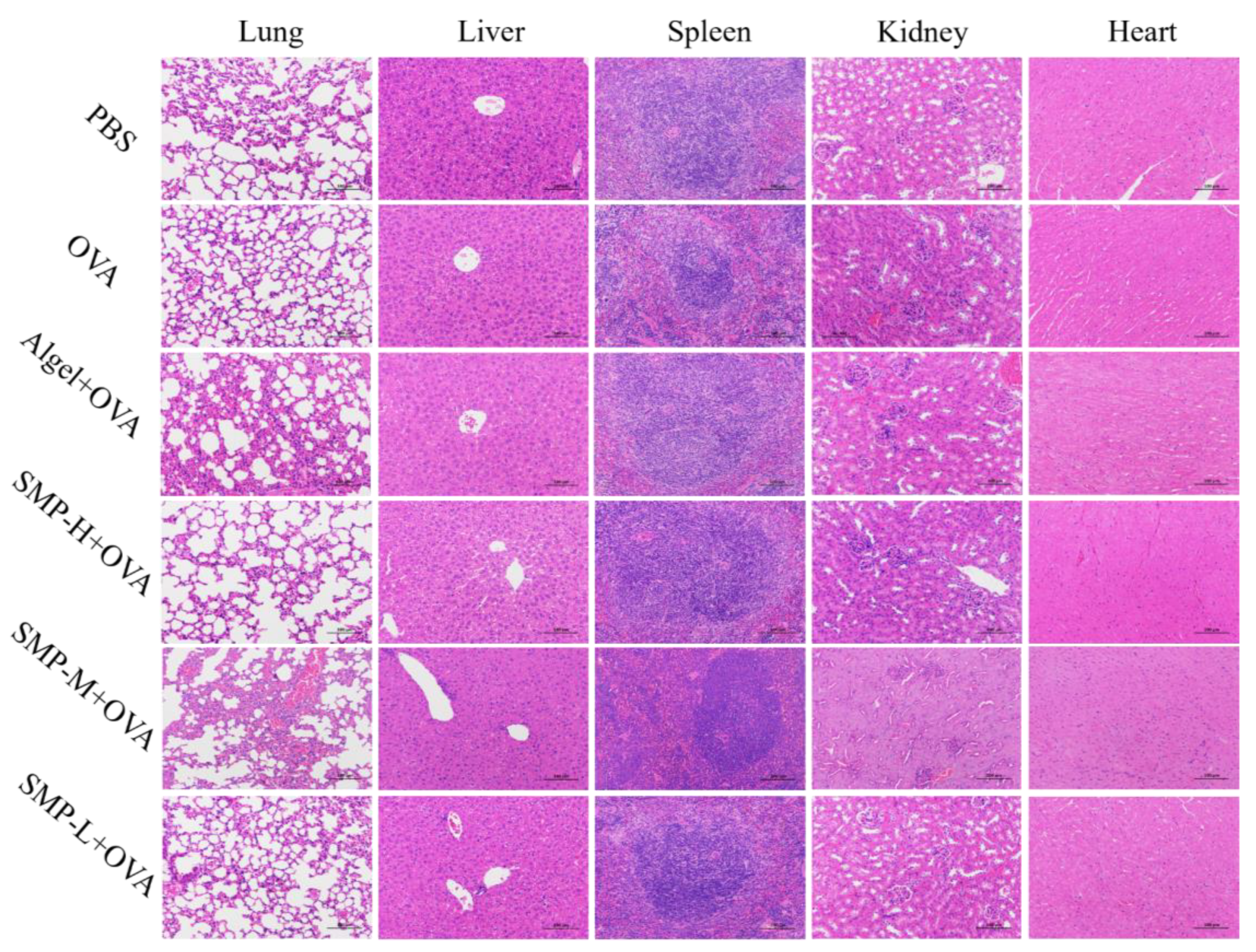
2.7. Biosafety Analysis of SMPD–2
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction and Purification of SMPs
4.3. Structural Characterization of SMPs
4.4. Animal and Vaccinations
4.5. Immunophenotypic Analysis of Lymph Node Cells
4.6. Determination of Antibody Responses
4.7. Splenic Lymphocyte Immunophenotype Analysis
4.8. Biochemistry Index
4.9. Histological Analysis of Organs
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Qiao, Z.; Ma, J.; Liu, H. Evaluation of the antioxidant potential of Salvia miltiorrhiza ethanol extract in a rat model of ischemia-reperfusion injury. Molecules 2011, 16, 10002–10012. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zuo, Z.; Chow, M.S.S. Danshen: An overview of its chemistry, pharmacology, pharmacokinetics, and clinical use. J. Clin. Pharmacol. 2005, 45, 1345–1359. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Wang, M.; Pang, Y.; Hu, X.; Sun, C.; Zhou, H.; Deng, Y.; Lu, S. The Smi-miR858a-SmMYB module regulates tanshinone and phenolic acid biosynthesis in Salvia miltiorrhiza. Hortic. Res. 2024, 11, uhae047. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-S.; Peng, W.-H.; Shih, M.-F.; Cherng, J.-Y. Anxiolytic effect of an extract of Salvia miltiorrhiza Bunge (Danshen) in mice. J. Ethnopharmacol. 2021, 264, 113285. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-Y.; Ming, Q.-L.; Rahman, K.; Han, T.; Qin, L.-P. Salvia miltiorrhiza: Traditional medicinal uses, chemistry, and pharmacology. Chin. J. Nat. Med. 2015, 13, 163–182. [Google Scholar] [CrossRef]
- Fu, Y.-P.; Peng, X.; Zhang, C.-W.; Jiang, Q.-X.; Li, C.-Y.; Paulsen, B.S.; Rise, F.; Huang, C.; Feng, B.; Li, L.-X.; et al. Salvia miltiorrhiza polysaccharide and its related metabolite 5-methoxyindole-3-carboxaldehyde ameliorate experimental colitis by regulating Nrf2/Keap1 signaling pathway. Carbohydr. Polym. 2023, 306, 120626. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, J.; Bao, J.; Lu, J.; Wang, Y. The anticancer properties of Salvia miltiorrhiza Bunge (Danshen): A systematic review. Med. Res. Rev. 2014, 34, 768–794. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Zhang, D.; Lou, H.; Sun, L.; Ji, J. Evaluation of the anti-inflammatory activities of tanshinones isolated from Salvia miltiorrhiza var. alba roots in THP-1 macrophages. J. Ethnopharmacol. 2016, 188, 193–199. [Google Scholar] [CrossRef]
- Wang, X.; Gao, A.; Jiao, Y.; Zhao, Y.; Yang, X. Antitumor effect and molecular mechanism of antioxidant polysaccharides from Salvia miltiorrhiza Bunge in human colorectal carcinoma LoVo cells. Int. J. Biol. Macromol. 2018, 108, 625–634. [Google Scholar] [CrossRef]
- Facciolà, A.; Visalli, G.; Laganà, A.; Di Pietro, A. An Overview of Vaccine Adjuvants: Current Evidence and Future Perspectives. Vaccines 2022, 10, 819. [Google Scholar] [CrossRef]
- Pogostin, B.H.; McHugh, K.J. Novel Vaccine Adjuvants as Key Tools for Improving Pandemic Preparedness. Bioengineering 2021, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Burakova, Y.; Madera, R.; McVey, S.; Schlup, J.R.; Shi, J. Adjuvants for Animal Vaccines. Viral Immunol. 2018, 31, 11–22. [Google Scholar] [CrossRef]
- Mbow, M.L.; De Gregorio, E.; Valiante, N.M.; Rappuoli, R. New adjuvants for human vaccines. Curr. Opin. Immunol. 2010, 22, 411–416. [Google Scholar] [CrossRef]
- Exley, C. Aluminium adjuvants and adverse events in sub-cutaneous allergy immunotherapy. Allergy Asthma Clin. Immunol. 2014, 10, 4. [Google Scholar] [CrossRef]
- Liu, Z.; Xing, J.; Huang, Y.; Bo, R.; Zheng, S.; Luo, L.; Niu, Y.; Zhang, Y.; Hu, Y.; Liu, J.; et al. Activation effect of Ganoderma lucidum polysaccharides liposomes on murine peritoneal macrophages. Int. J. Biol. Macromol. 2016, 82, 973–978. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Chen, G. Extraction, structural analysis, derivatization and antioxidant activity of polysaccharide from Chinese yam. Food Chem. 2021, 361, 130089. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yin, X.; Zhang, D.; Lu, J.; Wang, X. Isolation, Structural Characterization and Macrophage Activation Activity of an Acidic Polysaccharide from Raspberry Pulp. Molecules 2022, 27, 1674. [Google Scholar] [CrossRef]
- Wang, F.; Wang, W.; Huang, Y.; Liu, Z.; Zhang, J. Characterization of a novel polysaccharide purified from a herb of Cynomorium songaricum Rupr. Food Hydrocoll. 2015, 47, 79–86. [Google Scholar] [CrossRef]
- Zhang, X.; Cai, Z.; Mao, H.; Hu, P.; Li, X. Isolation and structure elucidation of polysaccharides from fruiting bodies of mushroom Coriolus versicolor and evaluation of their immunomodulatory effects. Int. J. Biol. Macromol. 2021, 166, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qiu, Z.; Li, L.; Vidyarthi, S.K.; Zheng, Z.; Zhang, R. Structural characterization and antioxidant activities of one neutral polysaccharide and three acid polysaccharides from Ziziphus jujuba cv. Hamidazao: A comparison. Carbohydr. Polym. 2021, 261, 117879. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, G.; Zhao, X.; Cao, X.; Wang, L.; Mu, J.; Qi, F.; Liu, L.; Zhang, H. Structural Characterization, Antioxidant and Antitumor Activities of the Two Novel Exopolysaccharides Produced by Debaryomyces hansenii DH-1. Int. J. Mol. Sci. 2022, 24, 335. [Google Scholar] [CrossRef] [PubMed]
- He, B.-L.; Zheng, Q.-W.; Guo, L.-Q.; Huang, J.-Y.; Yun, F.; Huang, S.-S.; Lin, J.-F. Structural characterization and immune-enhancing activity of a novel high-molecular-weight polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2020, 145, 11–20. [Google Scholar] [CrossRef]
- Chen, D.; Yang, L.; Yang, F.; Pei, Q.; Lu, L.; Huang, X.; Ouyang, P.; Geng, Y.; Li, Z.; Zhang, X.; et al. Salvia miltiorrhiza polysaccharide activated macrophages and improved the disease resistance of sturgeon against Aeromonas hydrophila. Fish. Shellfish Immunol. 2022, 127, 594–603. [Google Scholar] [CrossRef]
- Chen, Y.; Li, H.; Li, M.; Niu, S.; Wang, J.; Shao, H.; Li, T.; Wang, H. Salvia miltiorrhiza polysaccharide activates T Lymphocytes of cancer patients through activation of TLRs mediated -MAPK and -NF-κB signaling pathways. J. Ethnopharmacol. 2017, 200, 165–173. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Li, Y.; Yang, P.; Zhang, Z.; Wu, H.; Zhu, L.; Liu, Y. Preparation methods, structural characteristics, and biological activity of polysaccharides from Salvia miltiorrhiza: A review. J. Ethnopharmacol. 2023, 305, 116090. [Google Scholar] [CrossRef]
- Li, Y.Q.; Fang, L.; Zhang, K.C. Structure and bioactivities of a galactose rich extracellular polysaccharide from submergedly cultured Ganoderma lucidum. Carbohydr. Polym. 2007, 68, 323–328. [Google Scholar] [CrossRef]
- Zhang, J.; Song, Z.; Li, Y.; Zhang, S.; Bao, J.; Wang, H.; Dong, C.; Ohizumi, Y.; Xu, J.; Guo, Y. Structural analysis and biological effects of a neutral polysaccharide from the fruits of Rosa laevigata. Carbohydr. Polym. 2021, 265, 118080. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Wang, Y.; Niu, H.; Ma, Y.; Song, J. Isolation, purification, and physicochemical characterization of Polygonatum polysaccharide and its protective effect against CCl4-induced liver injury via Nrf2 and NF-κB signaling pathways. Int. J. Biol. Macromol. 2024, 261, 129863. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Fang, S.; Wang, Y.; Zhang, T.; Hu, S. Molecular mechanisms associated with macrophage activation by Rhizoma Atractylodis Macrocephalae polysaccharides. Int. J. Biol. Macromol. 2020, 147, 616–628. [Google Scholar] [CrossRef]
- Hu, F.; Yue, H.; Lu, T.; Ma, G. Cytosolic delivery of HBsAg and enhanced cellular immunity by pH-responsive liposome. J. Control. Release 2020, 324, 460–470. [Google Scholar] [CrossRef]
- Liu, Z.; Xing, J.; Zheng, S.; Bo, R.; Luo, L.; Huang, Y.; Niu, Y.; Li, Z.; Wang, D.; Hu, Y.; et al. Ganoderma lucidum polysaccharides encapsulated in liposome as an adjuvant to promote Th1-bias immune response. Carbohydr. Polym. 2016, 142, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Pang, G.; Chen, C.; Qin, J.; Yu, H.; Liu, Y.; Zhang, X.; Song, Z.; Zhao, J.; Wang, F.; et al. Effective cancer immunotherapy by Ganoderma lucidum polysaccharide-gold nanocomposites through dendritic cell activation and memory T cell response. Carbohydr. Polym. 2019, 205, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Courant, T.; Bayon, E.; Reynaud-Dougier, H.L.; Villiers, C.; Menneteau, M.; Marche, P.N.; Navarro, F.P. Tailoring nanostructured lipid carriers for the delivery of protein antigens: Physicochemical properties versus immunogenicity studies. Biomaterials 2017, 136, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kesharwani, S.S.; Kuppast, B.; Bakkari, M.A.; Tummala, H. Pathogen-mimicking vaccine delivery system designed with a bioactive polymer (inulin acetate) for robust humoral and cellular immune responses. J. Control. Release 2017, 261, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Trapani, J.A.; Smyth, M.J. Functional significance of the perforin/granzyme cell death pathway. Nat. Rev. Immunol. 2002, 2, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Chae, D.W.; Kim, S.M.; Nam, E.S.; Jang, M.K.; Lee, J.H.; Kim, H.Y.; Yoo, J.Y. Expression of FasL and perforin/granzyme B mRNA in chronic hepatitis B virus infection. J. Viral Hepat. 2004, 11, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Hu, F.; Yue, H.; Yang, T.; Ma, G. The incorporation of cationic property and immunopotentiator in poly (lactic acid) microparticles promoted the immune response against chronic hepatitis B. J. Control. Release 2020, 321, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, J.; Wang, Z. Outer membrane vesicles for vaccination and targeted drug delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1523. [Google Scholar] [CrossRef] [PubMed]
- Wusiman, A.; Xu, S.; Ni, H.; Gu, P.; Liu, Z.; Zhang, Y.; Qiu, T.; Hu, Y.; Liu, J.; Wu, Y.; et al. Immunomodulatory effects of Alhagi honey polysaccharides encapsulated into PLGA nanoparticles. Carbohydr. Polym. 2019, 211, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Wang, Y.; Kim, E.; Zhang, C.; Zhang, G.; Lee, Y. Structural Characteristics and Immunomodulatory Effects of a Long-Chain Polysaccharide from Laminaria japonica. Front. Nutr. 2022, 9, 762595. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, G.; Huang, H. Extraction, derivatization and antioxidant activities of onion polysaccharide. Food Chem. 2022, 388, 133000. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Huang, R.; Wen, P.; Song, Y.; He, B.; Tan, J.; Hao, H.; Wang, H. Structural characterization and immunological activity of pectin polysaccharide from kiwano (Cucumis metuliferus) peels. Carbohydr. Polym. 2021, 254, 117371. [Google Scholar] [CrossRef]
- Gu, P.; Wusiman, A.; Wang, S.; Zhang, Y.; Liu, Z.; Hu, Y.; Liu, J.; Wang, D. Polyethylenimine-coated PLGA nanoparticles-encapsulated Angelica sinensis polysaccharide as an adjuvant to enhance immune responses. Carbohydr. Polym. 2019, 223, 115128. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Chen, G.J.; Fan, C.Y.; Shen, F.; Yang, Y.; Pang, W.; Zhao, Z.N.; Guan, H.X.; Wu, H.; Lu, Y.; et al. An Adjustable Ad-juvant STINGsome for Tailoring the Potent and Broad Immunity Against SARS-CoV-2 and Monkeypox Virus via STING and Necroptosis. Adv. Funct. Mater. 2023, 33, 2306010. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, S.; Li, H.; An, J.; Li, C.; Zhou, R.; Teng, L.; Zhu, Y.; Liao, S.; Yang, Y.; et al. Structural characterization of Alpiniae oxyphyllae fructus polysaccharide 2 and its activation effects on RAW264.7 macrophages. Int. Immunopharmacol. 2021, 97, 107708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ma, W.; Zhang, J.; Song, X.; Sun, W.; Fan, Y. The immunoregulatory activities of astragalus polysaccharide liposome on macrophages and dendritic cells. Int. J. Biol. Macromol. 2017, 105, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ma, P.; Wang, H.; Zhang, C.; Sun, H.; Wang, C.; Song, C.; Leng, X.; Kong, D.; Ma, G. Immune responses to vaccines delivered by encapsulation into and/or adsorption onto cationic lipid-PLGA hybrid nanoparticles. J. Control. Release 2016, 225, 230–239. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Wang, L.; Liu, Y.; Zhang, W.; Fan, B.; Ma, X.; Yuan, Q.; Ma, G.; Su, Z. Enhanced Humoral and Cell-Mediated Immune Responses Generated by Cationic Polymer-Coated PLA Microspheres with Adsorbed HBsAg. Mol. Pharm. 2014, 11, 1772–1784. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycosidic Linkages | Derivative Name | MW | mol, (%) |
|---|---|---|---|
| t-Ara(f) | 1,4-di-O-acetyl-2,3,5-tri-O-methyl arabinitol | 279 | 25.78 |
| t-Xyl(p) | 1,5-di-O-acetyl-2,3,4-tri-O-methyl xylitol | 279 | 0.81 |
| t-Glc(p) | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl glucitol | 323 | 3.53 |
| t-Gal(p) | 1,5-di-O-acetyl-2,3,4,6-tetra-O-methyl galactitol | 323 | 4.46 |
| 5-Ara(f) | 1,4,5-tri-O-acetyl-2,3-di-O-methyl arabinitol | 307 | 8.05 |
| 2-Xyl(p) | 1,2,5-tri-O-acetyl-3,4-di-O-methyl xylitol | 307 | 0.77 |
| 3-Glc(p) | 1,3,5-tri-O-acetyl-2,4,6-tri-O-methyl glucitol | 351 | 6.36 |
| 4-Gal(p) | 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl galactitol | 351 | 25.71 |
| 6-Gal(p) | 1,5,6-tri-O-acetyl-2,3,4-tri-O-methyl galactitol | 351 | 4.14 |
| 2,4-Rha(p) | 1,2,4,5-tetra-O-acetyl-6-deoxy-3-O-methyl rhamnitol | 349 | 2.10 |
| 3,4-Glc(p) | 1,3,4,5-tetra-O-acetyl-2,6-di-O-methyl glucitol | 379 | 1.14 |
| 4,6-Glc(p) | 1,4,5,6-tetra-O-acetyl-2,3-di-O-methyl glucitol | 379 | 2.24 |
| 3,6-Glc(p) | 1,3,5,6-tetra-O-acetyl-2,4-di-O-methyl glucitol | 379 | 14.92 |
| Code | Glycosyl Residues | Chemical Shifts (ppm) | |||||
|---|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | ||
| A α-L-Araf-(1→ | 5.02/107.44 | 4.07/80.91 | 3.88/76.55 | 4.03/83.99 | 3.76, 3.66/61.23 | n.d/n.d | |
| B →4)-α-D-Galp-(1→ | 5.19/109.33 | 4.16/68.46 | 3.9/73.36 | 3.59/76.73 | 3.49/70.74 | 3.76, 3.62/61.1 | |
| C →3,6)-β-D-Glcp-(1→ | 4.43/103.15 | 3.48/75.11 | 3.66/80.12 | 3.78/71.2 | 3.87/73.36 | 3.82, 3.74/66.69 | |
| D →5)-α-L-Araf-(1→ | 5.09/106.89 | 4.08/81.27 | 4.23/79.23 | 4.16/81.35 | 3.98, 3.85/69.32 | n.d/n.d | |
| E →3)-α-D-Glcp-(1→ | 5.34/99.63 | 3.56/71.55 | 3.89/83.99 | 3.79/71.2 | 3.83/73.7 | 3.67/59.34 | |
| F →6)-α-D-Galp-(1→ | 5.19/106.41 | 3.91/72.95 | 3.65/75.81 | 3.79/71.17 | 3.9/73.75 | 3.83, 3.69/66.52 | |
| Group | ALT (U/L) | AST (U/L) | ALP (U/L) | UREA (mmol/L) | LDH (U/L) |
|---|---|---|---|---|---|
| PBS | 21.15 ± 5.972 | 67.89 ± 19.002 | 79.96 ± 7.216 | 3.40 ± 0.146 | 499.58 ± 64.260 |
| OVA | 26.61 ± 2.725 | 99.53 ± 1.056 | 171.24 ± 31.824 | 3.86 ± 0.528 | 806.21 ± 174.741 |
| Algel+OVA | 29.53 ± 4.626 | 94.46 ± 12.682 | 107.13 ± 21.800 | 3.79 ± 0.474 | 787.40 ± 56.323 |
| SMP-H+OVA | 35.36 ± 2.449 | 82.72 ± 26.902 | 80.11 ± 13.021 | 4.23 ± 0.287 | 672.34 ± 142.625 |
| SMP-M+OVA | 23.00 ± 7.500 | 100.34 ± 31.49 | 139.90 ± 34.009 | 3.30 ± 0.093 | 647.88 ± 100.273 |
| SMP-L+OVA | 27.77 ± 1.134 | 96.78 ± 53.936 | 92.37 ± 7.251 | 3.96 ± 0.243 | 737.83 ± 351.49 |
| Dyeing Group | Target for Testing | Color Matching [44] |
|---|---|---|
| 1 | T lymphocyte | anti-CD3-FITC, anti-CD4-APC, anti-CD8a-PE |
| 2 | Cytotoxic T Lymphocyte | anti-CD8a-PE, anti-CD107-FITC, anti-FASL-APC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Yang, X.; Gu, P.; Wang, X.; Bao, Y.; Shi, W. The Structural Characterization of a Polysaccharide from the Dried Root of Salvia miltiorrhiza and Its Use as a Vaccine Adjuvant to Induce Humoral and Cellular Immune Responses. Int. J. Mol. Sci. 2024, 25, 7765. https://doi.org/10.3390/ijms25147765
Zhu Y, Yang X, Gu P, Wang X, Bao Y, Shi W. The Structural Characterization of a Polysaccharide from the Dried Root of Salvia miltiorrhiza and Its Use as a Vaccine Adjuvant to Induce Humoral and Cellular Immune Responses. International Journal of Molecular Sciences. 2024; 25(14):7765. https://doi.org/10.3390/ijms25147765
Chicago/Turabian StyleZhu, Yixuan, Xiaochen Yang, Pengfei Gu, Xiao Wang, Yongzhan Bao, and Wanyu Shi. 2024. "The Structural Characterization of a Polysaccharide from the Dried Root of Salvia miltiorrhiza and Its Use as a Vaccine Adjuvant to Induce Humoral and Cellular Immune Responses" International Journal of Molecular Sciences 25, no. 14: 7765. https://doi.org/10.3390/ijms25147765
APA StyleZhu, Y., Yang, X., Gu, P., Wang, X., Bao, Y., & Shi, W. (2024). The Structural Characterization of a Polysaccharide from the Dried Root of Salvia miltiorrhiza and Its Use as a Vaccine Adjuvant to Induce Humoral and Cellular Immune Responses. International Journal of Molecular Sciences, 25(14), 7765. https://doi.org/10.3390/ijms25147765