

The GLP-1 Receptor Agonist Liraglutide Decreases Primary Bile Acids and Serotonin in the Colon Independently of Feeding in Mice



Abstract
:1. Introduction
2. Results
2.1. Effects of Liraglutide on Body Weight, Blood Glucose, Plasma Insulin, Serotonin, and FGF21 Levels in Mice Deprived of Food after Intraperitoneal Administration of the Drug
2.2. Effects of Liraglutide on Tryptophan Metabolites in the Colon of Mice
2.3. Effects of Liraglutide on Bile Acids in the Feces and Liver of Mice
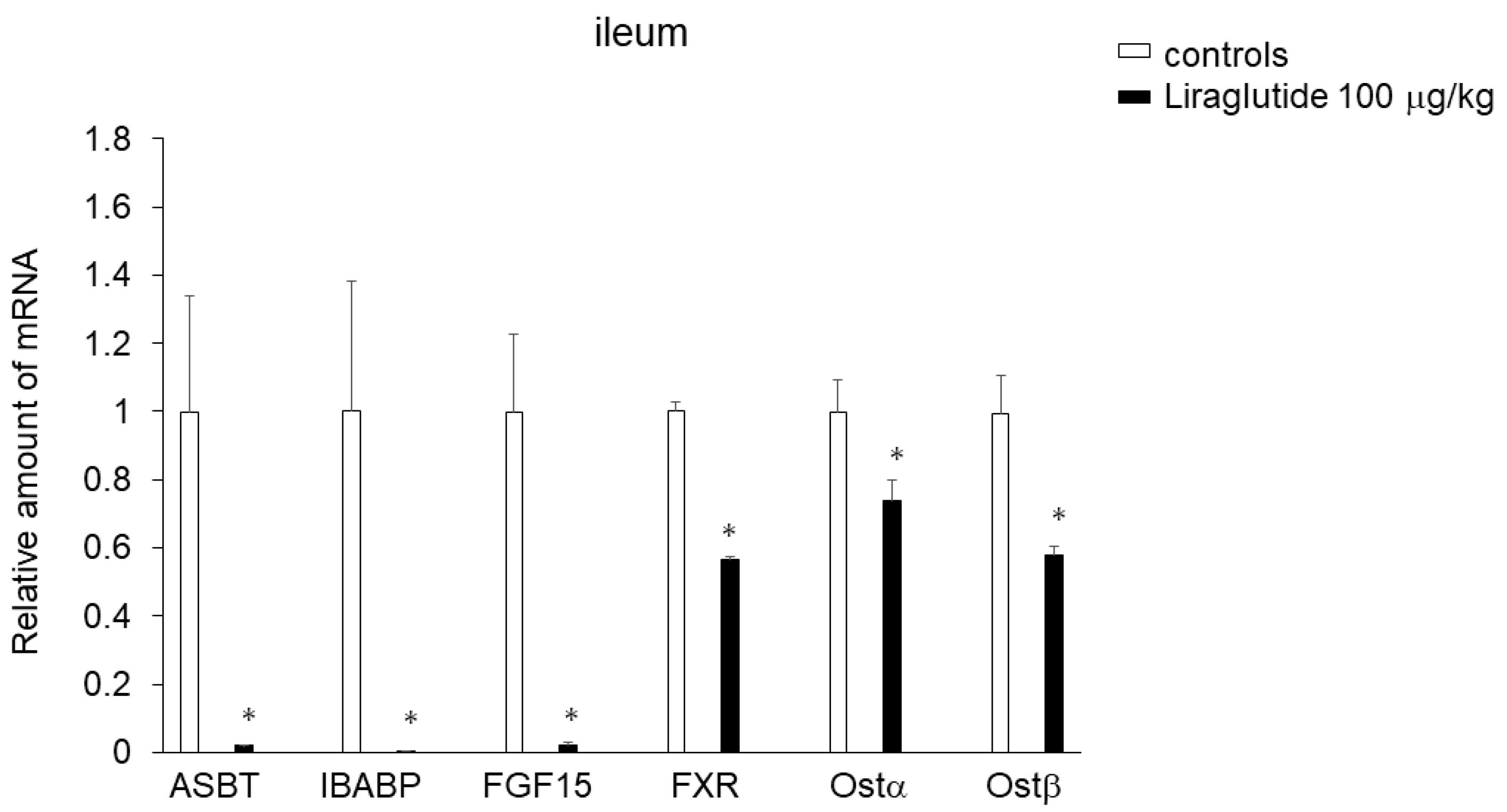
2.4. Effects of Liraglutide on Expression of Genes Involved in BA Transport in the Ileum
3. Discussion
4. Materials and Methods
4.1. General Procedures
4.2. Blood Chemistry
4.3. BA Analysis
4.4. Tryptophan and Its Metabolites Analysis
4.5. Real-Time Quantitative Reverse Transcription–Polymerase Chain Reaction (RT–PCR)
4.6. Statistical Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Campbell, J.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of incretin hormone action. Cell Metab. 2013, 17, 819–837. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes—state-of-the-art. Mol. Metab. 2021, 46, 101102. [Google Scholar] [CrossRef] [PubMed]
- Popoviciu, M.S.; Păduraru, L.; Yahya, G.; Metwally, K.; Cavalu, S. Emerging Role of GLP-1 Agonists in Obesity: A Comprehensive Review of Randomised Controlled Trials. Int. J. Mol. Sci. 2023, 24, 10449. [Google Scholar] [CrossRef] [PubMed]
- Kårhus, M.L.; Brønden, A.; Forman, J.L.; Haaber, A.; Knudsen, E.; Langholz, E.; Dragsted, L.O.; Hansen, S.H.; Krakauer, M.; Vilsbøll, T.; et al. Safety and efficacy of liraglutide versus colesevelam for the treatment of bile acid diarrhoea: A randomised, double-blind, active-comparator, non-inferiority clinical trial. Lancet Gastroenterol. Hepatol. 2022, 7, 922–931. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K. The Regulatory Role of the Central and Peripheral Serotonin Network on Feeding Signals in Metabolic Diseases. Int. J. Mol. Sci. 2022, 23, 1600. [Google Scholar] [CrossRef]
- Koopman, N.; Katsavelis, D.; Hove, A.S.T.; Brul, S.; Jonge, W.J.; Seppen, J. The Multifaceted Role of Serotonin in Intestinal Homeostasis. Int. J. Mol. Sci. 2021, 22, 9487. [Google Scholar] [CrossRef] [PubMed]
- Chojnacki, J.; Konrad, P.; Mędrek-Socha, M.; Kaczka, A.; Błońska, A.; Zajdel, R.; Chojnacki, C.; Gasiorowska, A. The Variability of Tryptophan Metabolism in Patients with Mixed Type of Irritable Bowel Syndrome. Int. J. Mol. Sci. 2024, 25, 2550. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, J.; Wu, X.; Jing, H.; Zhang, S.; Hu, Z.; Rao, L.; Chang, Q.; Wang, L.; Zhang, Z. Gut microbiota alteration after cholecystectomy contributes to post-cholecystectomy diarrhea via bile acids stimulating colonic serotonin. Gut Microbes 2023, 15, 2168101. [Google Scholar] [CrossRef]
- Martin, A.M.; Yabut, J.M.; Choo, J.M.; Page, A.J.; Sun, E.W.; Jessup, C.F.; Wesselingh, S.L.; Khan, W.I.; Rogers, G.B.; Steinberg, G.R.; et al. The gut microbiome regulates host glucose homeostasis via peripheral serotonin. Proc. Natl. Acad. Sci. USA 2019, 116, 19802–19804. [Google Scholar] [CrossRef]
- Nonogaki, K.; Kaji, T. The acute anorexic effect of liraglutide, a GLP-1 receptor agonist, does not require functional leptin receptor, serotonin, and hypothalamic POMC and CART activities in mice. Diabetes Res. Clin. Pract. 2016, 120, 186–189. [Google Scholar] [CrossRef]
- Nonogaki, K.; Kaji, T. Liraglutide, a GLP-1 Receptor Agonist, Which Decreases Hypothalamic 5-HT2A Receptor Expression, Reduces Appetite and Body Weight Independently of Serotonin Synthesis in Mice. J. Diabetes Res. 2018, 2018, 6482958. [Google Scholar] [CrossRef] [PubMed]
- Borup, C.; Vinter-Jensen, L.; Jørgensen, S.P.G.; Wildt, S.; Graff, J.; Gregersen, T.; Zaremba, A.; Borup Andersen, T.; Nøjgaard, C.; Timm, H.B.; et al. Efficacy and safety of colesevelam for the treatment of bile acid diarrhoea: A double-blind, randomised, placebo-controlled, phase 4 clinical trial. Lancet Gastroenterol. Hepatol. 2023, 8, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Sagar, N.M.; Duboc, H.; Kay, G.L.; Alam, M.T.; Wicaksono, A.N.; Covington, J.A.; Quince, C.; Kokkorou, M.; Svolos, V.; Palmieri, L.J.; et al. The pathophysiology of bile acid diarrhoea: Differences in the colonic microbiome, metabolome and bile acids. Sci. Rep. 2020, 10, 20436. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.L.; Egerod, K.L.; Engelstoft, M.S.; Dmytriyeva, O.; Theodorsson, E.; Patel, B.A.; Schwartz, T.W. Enterochromaffin 5-HT cells—A major target for GLP-1 and gut microbial metabolites. Mol. Metab. 2018, 11, 70–83. [Google Scholar] [CrossRef]
- Rao, A.; Haywood, J.; Craddock, A.L.; Belinsky, M.G.; Kruh, G.D.; Dawson, P.A. The organic solute transporter alpha-beta, Ostalpha-Ostbeta, is essential for intestinal bile acid transport and homeostasis. Proc. Natl. Acad. Sci. USA 2008, 105, 3891–3896. [Google Scholar] [CrossRef]
- Lundåsen, T.; Andersson, E.-M.; Snaith, M.; Lindmark, H.; Lundberg, J.; Östlund-Lindqvist, A.-M. Inhibition of intestinal bile acid transporter Slc10a2 improves triglyceride metabolism and normalizes elevated plasma glucose levels in mice. PLoS ONE 2012, 7, e37787. [Google Scholar] [CrossRef]
- Ge, M.-X.; Niu, W.-X.; Ren, J.-F.; Cai, S.-Y.; Yu, D.-K.; Liu, H.-T.; Zhang, N.; Zhang, Y.X.; Wang, Y.C.; Shao, R.G.; et al. A novel ASBT inhibitor, IMB17-15, repressed nonalcoholic fatty liver disease development in high-fat diet-fed Syrian golden hamsters. Acta Pharmacol. Sin. 2019, 40, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Kosters, A.; Mells, J.E.; Zhang, W.; Setchell, K.D.; Amanso, A.M.; Wynn, G.M.; Xu, T.; Keller, B.T.; Yin, H.; et al. Inhibition of ileal bile acid uptake protects against nonalcoholic fatty liver disease in high-fat diet-fed mice. Sci. Transl. Med. 2016, 8, 357ra122. [Google Scholar] [CrossRef]
- Hansen, M.; Sonne, D.P.; Mikkelsen, K.H.; Gluud, L.L.; Vilsbøll, T.; Knop, F.K. Bile acid sequestrants for glycemia control in patients with type 2 diabetes: A systematic review with meta-analysis of randomized controlled trials. J. Diabetes Complicat. 2017, 31, 918–927. [Google Scholar] [CrossRef]
- Hartmann, P.; Duan, Y.; Miyamoto, Y.; Demir, M.; Lang, S.; Hasa, E.; Stern, P.; Yamashita, D.; Conrad, M.; Eckmann, L.; et al. Colesevelam ameliorates non-alcoholic steatohepatitis and obesity in mice. Hepatol. Int. 2022, 16, 359–370. [Google Scholar] [CrossRef]
- Iglesias, P.; Selgas, R.; Romero, S.; Díez, J.J. Biological role, clinical significance, and therapeutic possibilities of the recently discovered metabolic hormone fibroblastic growth factor 21. Eur. J. Endocrinol. 2012, 167, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.D.; Palanivel, R.; Mottillo, E.P.; Bujak, A.L.; Wang, H.; Ford, R.J.; Collins, A.; Blümer, R.M.; Fullerton, M.D.; Yabut, J.M.; et al. Inhibiting peripheral serotonin synthesis reduces obesity and metabolic dysfunction by promoting brown adipose tissue thermogenesis. Nat. Med. 2015, 21, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Jiang, M.; Zhao, J.; Song, Y.; Du, W.; Shi, J. The Mechanism Underlying the Influence of Indole-3-Propionic Acid: A Relevance to Metabolic Disorders. Front. Endocrinol. 2022, 13, 841703. [Google Scholar] [CrossRef] [PubMed]
- Tuomainen, M.; Lindström, J.; Lehtonen, M.; Auriola, S.; Pihlajamäki, J.; Peltonen, M.; Tuomilehto, J.; Uusitupa, M.; de Mello, V.D.; Hanhineva, K. Associations of serum indolepropionic acid, a gut microbiota metabolite, with type 2 diabetes and low-grade inflammation in high-risk individuals. Nutr. Diabetes 2018, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- de Mello, V.D.; Paananen, J.; Lindström, J.; Lankinen, M.A.; Shi, L.; Kuusisto, J.; Pihlajamäki, J.; Auriola, S.; Lehtonen, M.; Rolandsson, O.; et al. Indolepropionic acid and novel lipid metabolites are associated with a lower risk of type 2 diabetes in the Finnish Diabetes Prevention Study. Sci. Rep. 2017, 7, 46337. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, Y.; Sun, S.; Xie, Y.; Pan, C.; Li, M.; Li, C.; Liu, Y.; Xu, Z.; Liu, W.; et al. Indolepropionic acid reduces obesity-induced metabolic dysfunction through colonic barrier restoration mediated via tuft cell-derived IL-25. FEBS J. 2022, 289, 5985–6004. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K.; Kaji, T. Whey protein isolate inhibits hepatic FGF21 production, which precedes weight gain, hyperinsulinemia and hyperglycemia in mice fed a high-fat diet. Sci. Rep. 2020, 10, 15784. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, K.; Kaji, T. Ingestion of whey protein and β-conglycinin exerts opposite effects on intestinal FGF15 and serotonin secretion in mice. Front. Endocrinol. 2023, 14, 1080790. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| (ng/g) | Control | Liraglutide | p-Value |
|---|---|---|---|
| Trp | 15,620 ± 4608.09 | 3027 ± 220.4 | p < 0.05 |
| 5-HT | 1367 ± 120.7 | 385 ± 13.66 | p < 0.05 |
| 5-HIAA | 462 ± 66.0 | 266 ± 5.42 | p < 0.05 |
| KYN | 311 ± 56.93 | 46 ± 0.82 | p < 0.05 |
| XA | 417 ± 100.68 | 72.1 ± 7.72 | p < 0.05 |
| IPA | 29.9 ± 1.91 | 652.5 ± 119.94 | p < 0.05 |
| (nmol/g) | Control | Liraglutide | p-Value |
|---|---|---|---|
| CA | 291.18 ± 68.59 | 16.53 ± 1.79 | p < 0.05 |
| TCA | 70.65 ± 23.95 | 5.42 ± 2.3 | p < 0.05 |
| TCDCA | 2.55 ± 0.75 | 0.39 ± 0.12 | p < 0.05 |
| GCA | 0.57 ± 0.11 | 0.07 ± 0.01 | p < 0.05 |
| αMCA | 37.13 ± 11.25 | 21.48 ± 4.21 | NS |
| βMCA | 176.93 ± 62.51 | 70.60 ± 17.04 | p < 0.05 |
| DCA | 382.08 ± 108.4 | 407.5 ± 72.23 | NS |
| UDCA | 28.16 ± 9.10 | 31.6 ± 8.73 | NS |
| TDCA | 4.54 ± 1.50 | 1.3795 ± 0.41 | p < 0.05 |
| LCA | 11.88 ± 4.37 | 20.89 ± 3.89 | NS |
| GDCA | 0.27 ± 0.06 | 0.37 ± 0.10 | NS |
| TBA | 1008.78 ± 155.75 | 439.81 ± 87.28 | p < 0.05 |
| (nmol/g) | Control | Liraglutide | p-Value |
|---|---|---|---|
| CA | 2.72 ± 0.14 | 1.45 ± 0.41 | p < 0.05 |
| TCA | 131.57 ± 23.57 | 39.55 ± 0.47 | p < 0.05 |
| TCDCA | 2.36 ± 0.15 | 2.01 ± 0.46 | NS |
| GCA | 0.18 ± 0.03 | 0.16 ± 0.05 | NS |
| CDCA | 0.31 ± 0.02 | 0.24 ± 0.05 | NS |
| αMCA | 0.91 ± 0.12 | 0.78 ± 0.18 | NS |
| βMCA | 5.24 ± 0.26 | 3.57 ± 0.19 | p < 0.05 |
| DCA | 0.21 ± 0.004 | 0.20 ± 0.01 | NS |
| UDCA | 0.25 ± 0.02 | 0.18 ± 0.08 | p < 0.05 |
| TDCA | 10.28 ± 0.99 | 8.57 ± 1.82 | NS |
| LCA | 0.06 ± 0.02 | 0.08 ± 0.06 | NS |
| TBA | 154.1 ± 23.46 | 43.54 ± 6.57 | p < 0.05 |
| GENES | Sense | Antisense |
|---|---|---|
| FGF15 | ACGGGCTGATTCGCTACTC | TGTAGCCTAAACAGTCCATTTCCT |
| FXR | CCCCTGCTTGATGTGCTAC | CGTGGTGATGGTTGAATGTC |
| ASBT | TGGGTTTCTTCCTGGCTAGACT | TGTTCTGCATTCCAGTTTCCAA |
| IBABP | CAGGAGACGTGATTGAAAGGG | GCCCCCAGAGTAAGACTGGG |
| Ostα | TACAAGAACACCCTTTGCCC | CGAGGAATCCAGAGACCAAA |
| Ostβ | GTATTTTCGTGCAGAAGATGCG | TTTCTGTTTGCCAGGATGCTC |
| β-actin | TTGTAACCAACTGGGACGATATGG | GATCTTGATCTTCATGGTGCTAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nonogaki, K.; Kaji, T. The GLP-1 Receptor Agonist Liraglutide Decreases Primary Bile Acids and Serotonin in the Colon Independently of Feeding in Mice. Int. J. Mol. Sci. 2024, 25, 7784. https://doi.org/10.3390/ijms25147784
Nonogaki K, Kaji T. The GLP-1 Receptor Agonist Liraglutide Decreases Primary Bile Acids and Serotonin in the Colon Independently of Feeding in Mice. International Journal of Molecular Sciences. 2024; 25(14):7784. https://doi.org/10.3390/ijms25147784
Chicago/Turabian StyleNonogaki, Katsunori, and Takao Kaji. 2024. "The GLP-1 Receptor Agonist Liraglutide Decreases Primary Bile Acids and Serotonin in the Colon Independently of Feeding in Mice" International Journal of Molecular Sciences 25, no. 14: 7784. https://doi.org/10.3390/ijms25147784
APA StyleNonogaki, K., & Kaji, T. (2024). The GLP-1 Receptor Agonist Liraglutide Decreases Primary Bile Acids and Serotonin in the Colon Independently of Feeding in Mice. International Journal of Molecular Sciences, 25(14), 7784. https://doi.org/10.3390/ijms25147784