
Skin Malignant Melanoma and Matrix Metalloproteinases: Promising Links to Efficient Therapies

 , and
, and
Abstract
1. Introduction
The Aims of the Review
2. Current Understanding of Malignant Melanoma (MM)
2.1. MM Etiopathogenesis: Melanin Role in MM Development, Risk Factors, Hereditary and Acquired Genetic Mutations (Figure 1)
2.1.1. Melanocytes and Melanin Role in MM Development
MM Risk Factors
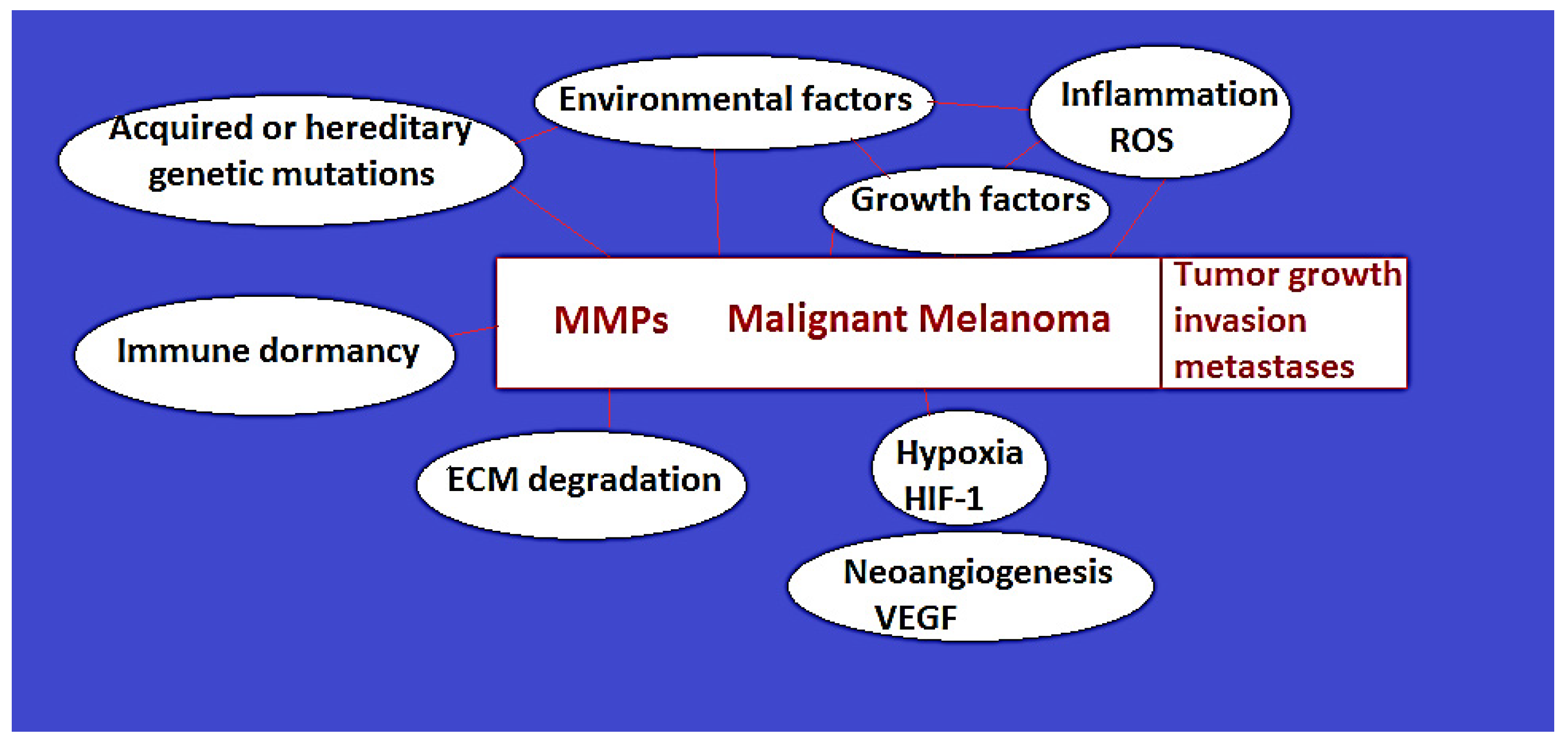
2.1.2. Hereditary Genetic Factors
2.1.3. Lifetime-Acquired Genetic Mutations
2.1.4. UV Radiation and Exposure to Other Risk Factors: Mechanisms of Triggering Genetic Mutations and MM
2.2. Malignant Melanoma Types
2.3. MM Diagnostic
2.4. MM Tumor Stages, Treatment, and Prognostic
2.4.1. MM Tumor Stages
2.4.2. MM Treatment
2.4.3. MM Patients’ Prognostic
2.5. MM Metastases
3. Current Understanding of the Role of Matrix Metalloproteinases in MM (Figure 1)
3.1. Background Information on Matrix Metalloproteinases (MMPs)

{kind=link}
{kind=link}
| MMP Class (Type) | MMP Sub-Group | Sources | Physiologic Function | Role in MM |
|---|---|---|---|---|
| MMP-1 (interstitial collagenase) | Collagenases | Stromal cells (tumor-associated fibroblasts) | Fibrillar colagen degradation: collagen type I and III | MM invasiveness, vertical growth pattern, metastases; aggressiveness (non-regressive type) |
| MMP-2 (gelatinase A) | Gelatinases | Stromal cells, fibroblasts, endothelial cells, inflammatory cells, cardiomyocytes, benign melanocytic and MM cells | Collagen IV, as well as other non-fibrillar collagens (V, VII, X), gelatin, fibronectin, and type I collagen; contractile proteins, such as light chain myosin and troponin | MM radial growth pattern; tumor angiogenesis; aggressiveness and tendency to distant metastasizing; prognostic factor and predictor for vemurafenib resistance |
| MMP-3 (stromelysin-1) | Stromelysins | Stromal (fibroblasts) and tumor cells | Degrades collagen type I | Activates MMP-1, MMP-7, and MMP-13, associated with MM invasiveness and metastases |
| MMP-7 (matrilysin-1)-smallest MMP | Matrilysins | MM, tumor cells, stromal cells | Degrades elastin—degrades many ECM components, such as collagen IV, entactin, laminin, syndecan-1, cadherin, fibronectin, cartilage proteoglycans | Tumor invasiveness and metastases |
| MMP-9 (gelatinase B) | Gelatinases | Keratinocytes, fibroblasts, immune cells (neutrophils, macrophages), tumor cells | Gelatin, collagen type IV—component of the skin basement membrane, fibronectin, elastin, laminin, fibrillin; activates growth factors, including vascular endothelial growth factor, tumor necrosis factor-alpha, transforming growth factor beta, and fibroblast growth factor | MM radial growth pattern; promotes neoangiogenesis; putative anti-tumor effect in breast or colonic neoplasia; overexpression and activation of MMP-9 in the most aggressive forms of malignant melanoma; metastases |
| MMP-11 (stromelysin 3) | Stromelysins | Limited, contradictory knowledge in MM (MM progression versus regression) | ||
| MMP-12 | Other types (Metalloelastase) | Fibroblasts, macrophages | Elastin | Tumor invasiveness, lymph node, and distant metastases |
| MMP-13 (collagenase-3) | Collagenases | Fibroblasts, especially tumor-associated cells | Collagen, gelatin, casein, as well as fibrinogen and growth factors | MM invasive vertical growth (metastatic) |
| MMP-23 | Other types (a membrane-type MMP) | Membrane-type MMP, found in the endoplasmic reticulum and perinuclear membranes; Malignant melanoma cells | Cytokines, chemokines, blocks Kv1.3 on T-cells | Diminished immune anti-tumor cells response; resistance to adjuvant therapies |
3.2. Physiologic Regulation of MMPs’ Level of Activity
MMPs Endogenous Inhibitors
3.3. Mechanisms behind the Increased Expression and Activity of Matrix Metalloproteinases in MM
4. MMPs’ Role in MM
4.1. MMPs’ Involvement in Tumor Development, Progression, and Metastasis
4.2. Characteristics of the Matrix Metalloproteinases That Play Roles in Malignant Melanoma Development and Progression
5. MMPs as Therapeutic Targets: Knowledge of MMPs’ Inhibitors (MMPIs) and Their Experimental and Clinical Use in MM Treatment
5.1. Synthetic Inhibitors: Results in Preclinical and Clinical Trials and Limitations of Use
5.2. Natural Inhibitors
5.3. Current Understanding of MMPIs’ Efficacy in Cancers, Including MM
5.4. Possible Explanations for the Disappointing Efficacy of MMPIs in MM
6. Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eddy, K.; Shah, R.; Chen, S. Decoding melanoma development and progression: Identification of therapeutic vulnerabilities. Front. Oncol. 2021, 10, 626129. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Napoli, S.; Scuderi, C.; Gattuso, G.; Bella, V.D.; Candido, S.; Basile, M.S.; Libra, M.; Falzone, L. Functional roles of matrix metalloproteinases and their inhibitors in melanoma. Cells 2020, 9, 1151. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of matrix metalloproteinases in photoaging and photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fruntelată, R.F.; Bakri, A.; Stoica, G.A.; Mogoantă, L.; Ionovici, N.; Popescu, G.; Pîrşcoveanu, D.F.V.; Raicea, A.; Ciurea, M.E. Assessment of tumoral and peritumoral inflammatory reaction in cutaneous malignant melanomas. Rom. J. Morphol. Embryol. 2023, 64, 41–48. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Davis, L.E.; Shalin, S.C.; Tackett, A.J. Current state of melanoma diagnosis and treatment. Cancer Biol. Ther. 2019, 20, 1366–1379. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Brînzea, A.; Nedelcu, R.I.; Ion, D.A.; Turcu, G.; Antohe, M.; Hodorogea, A.; Călinescu, A.; Pirici, D.; Popescu, R.; Popescu, C.M.; et al. Matrix metalloproteinases expression in lentigo maligna/lentigo maligna melanoma—A review of the literature and personal experience. Rom. J. Morphol. Embryol. 2019, 60, 1091–1095. [Google Scholar] [PubMed]
- Zob, D.L.; Augustin, I.; Caba, L.; Panzaru, M.C.; Popa, S.; Popa, A.D.; Florea, L.; Gorduza, E.V. Genomics and epigenomics in the molecular biology of melanoma—A prerequisite for biomarkers studies. Int. J. Mol. Sci. 2022, 24, 716. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tas, F. Metastatic behavior in melanoma: Timing, pattern, survival, and influencing factors. J. Oncol. 2012, 2012, 647684. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hofmann, U.B.; Westphal, J.R.; Zendman, A.J.W.; Becker, J.C.; Ruiter, D.J.; van Muijen, G.N.P. Expression and activation of matrix metalloproteinase-2 (MMP-2) and its co-localization with membrane type 1 matrix metalloproteinase (MT1-MMP) correlate with melanoma progression. J. Pathol. 2000, 191, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Naik, P.P. Role of biomarkers in the integrated management of melanoma. Dis. Mark. 2021, 2021, 6238317. [Google Scholar] [CrossRef] [PubMed]
- Zbytek, B.; Carlson, J.A.; Granese, J.; Ross, J.; Mihm, M.C., Jr.; Slominski, A. Current concepts of metastasis in melanoma. Expert. Rev. Dermatol. 2008, 3, 569–585. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ricciardi, E.; Giordani, E.; Ziccheddu, G.; Falcone, I.; Giacomini, P.; Fanciulli, M.; Russillo, M.; Cerro, M.; Ciliberto, G.; Morrone, A.; et al. Metastatic Melanoma: Liquid Biopsy as a New Precision Medicine Approach. Int. J. Mol. Sci. 2023, 24, 4014. [Google Scholar] [CrossRef] [PubMed]
- Mahumud, R.A.; Shahjalal, M. The Emerging Burden of Genetic Instability and Mutation in Melanoma: Role of Molecular Mechanisms. Cancers 2022, 14, 6202. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.G.; Miller, N.; McInerney, N.M. A Review of Epidemiology and Cancer Biology of Malignant Melanoma. Cureus 2021, 13, e15087. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Yanping, C.; Tong, L.; Ning, L.; Yufeng, L.; Geyu, L. Predicting the Risk of Melanoma Metastasis Using an Immune Risk Score in the Melanoma Cohort. Front. Bioeng. Biotechnol. 2020, 8, 206. [Google Scholar] [CrossRef] [PubMed]
- Sundararajan, S.; Thida, A.M.; Yadlapati, S.; Koya, S. Metastatic Melanoma. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://pubmed.ncbi.nlm.nih.gov/29262232/ (accessed on 6 June 2024). [PubMed]
- Keung, E.Z.; Gershenwald, J.E. The eighth edition American Joint Committee on Cancer (AJCC) melanoma staging system: Implications for melanoma treatment and care. Expert. Rev. Anticancer Ther. 2018, 18, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Pathak, S.; Zito, P.M. Clinical Guidelines for the Staging, Diagnosis, and Management of Cutaneous Malignant Melanoma. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK572149/ (accessed on 25 August 2022).
- Savage, P. Malignant melanoma (non-metastatic). BMJ Clin. Evid. 2007, 2007, 1705. [Google Scholar] [PubMed]
- Gerlini, G.; Tripo, L.; Sestini, S.; Brandani, P.; Giannotti, V.; Gattai, R.; Borgognoni, L. Melanoma metastases occuring 40 years after primary melanoma. Acta Oncol. 2018, 57, 1418–1420. [Google Scholar] [CrossRef] [PubMed]
- Mervic, L. Time course and pattern of metastasis of cutaneous melanoma differ between men and women. PLoS ONE 2012, 7, e32955. [Google Scholar] [CrossRef]
- Bastian, A.; Nichita, L.; Zurac, S. Matrix metalloproteinases in melanoma with and without regression. In The Role of Matrix Metalloproteinase in Human Body Pathologies; Tavascio, F., Ed.; IntechOpen Ltd.: London, UK, 2017. [Google Scholar] [CrossRef]
- Singvogel, K.; Schittek, B. Dormancy of cutaneous melanoma. Cancer Cell Int. 2024, 24, 88. [Google Scholar] [CrossRef]
- Tejera-Vaquerizo, A.; Nagore, E.; Meléndez, J.J.; López-Navarro, N.; Martorell-Calatayud, A.; Herrera-Acosta, E.; Traves, V.; Guillén, C.; Herrera-Ceballos, E. Chronology of metastasis in cutaneous melanoma: Growth rate model. J. Investig. Dermatol. 2012, 132, 1215–1221. [Google Scholar] [CrossRef]
- Waheed Roomi, M.; Efremov, E.; Niedzwiecki, A.; Rath, M. Matrix metalloproteinases-9 as a promising target for anti-cancer vaccine: Inhibition of melanoma tumor growth in mice immunized with syngeneic MMP-9 peptides. WCRJ 2019, 6, e1421. [Google Scholar] [CrossRef]
- Hofmann, U.B.; Westphal, J.R.; Van Muijen, G.N.; Ruiter, D.J. Matrix metalloproteinases in human melanoma. J. Investig. Dermatol. 2000, 115, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Anchan, A.; Finlay, G.; Angel, C.E.; Hucklesby, J.J.W.; Graham, S.E. Melanoma Mediated Disruption of Brain Endothelial Barrier Integrity Is Not Prevented by the Inhibition of Matrix Metalloproteinases and Proteases. Biosensors 2022, 12, 660. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Peng, K.; Zhang, Y.; Liu, D.; Chen, J. MMP2 is a immunotherapy related biomarker and correlated with cancer-associated fibroblasts infiltrate in melanoma. Cancer Cell Int. 2023, 23, 26. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mastalier Manolescu, B.S.; Lazar, A.M.; Ţiplica, G.S.; Zurac, S.A.; Reboşapcă, A.; Andreescu, B.; Popp, C.G. MMP1, MMP9, MMP11 and MMP13 in melanoma and its metastasis—Key points in understanding the mechanisms and celerity of tumor dissemination. Rom. J. Morphol. Embryol. 2024, 65, 45–52. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hadler-Osler, E.; Fadnes, B.; Stylte, I.; Uhlin-Hansen, L.; Winberg, J.-O. Regulation of matrix metalloproteinase activity in health and disease. FEBS J. 2011, 278, 28–45. [Google Scholar] [CrossRef]
- He, J.; Qin, M.; Chen, Y.; Hu, Z.; Xie, F.; Ye, L.; Hui, T. Epigenetic regulation of matrix metalloproteinases in inflammatory diseases: A narrative review. Cell Biosci. 2020, 10, 86. [Google Scholar] [CrossRef]
- Tatti, O.; Arjama, M.; Ranki, A.; Weiss, S.J.; Keski-Oja, J.; Lehti, K. Membrane-type-3 matrix metalloproteinase (MT3-MMP) functions as a matrix composition-dependent effector of melanoma cell invasion. PLoS ONE 2011, 6, e28325. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Laronha, H.; Carpinteiro, I.; Portugal, J.; Azul, A.; Polido, M.; Petrova, K.T.; Salema-Oom, M.; Caldeira, J. Challenges in Matrix Metalloproteinases Inhibition. Biomolecules 2020, 10, 717. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jabłońska-Trypuć, A.; Matejczyk, M.; Rosochacki, S. Matrix metalloproteinases (MMPs), the main extracellular matrix (ECM) enzymes in collagen degradation, as a target for anticancer drugs. J. Enzyme Inhib. Med. Chem. 2016, 31 (Suppl. 1), 177–183. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Jang, N.H.; Lee, H.J. Natural Products as Regulators against Matrix Metalloproteinases for the Treatment of Cancer. Biomedicines 2024, 12, 794. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Serraino, G.F.; Jiritano, F.; Costa, D.; Ielapi, N.; Battaglia, D.; Bracale, U.M.; Mastroroberto, P.; Andreucci, M.; Serra, R. Metalloproteinases in cardiac surgery: A systematic review. Biomolecules 2023, 13, 113. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gonzalez-Avila, G.; Sommer, B.; Mendoza-Posada, D.A.; Ramos, C.; Garcia-Hernandez, A.A.; Falfan-Valencia, R. Matrix metalloproteinases participation in the metastatic process and theirdiagnostic and therapeutic applications in cancer. Crit. Rev. Oncol. Hematol. 2019, 137, 57–83, Erratum in Crit. Rev. Oncol. Hematol. 2019, 138, 172. [Google Scholar] [CrossRef] [PubMed]
- Bassiouni, W.; Ali, M.A.M.; Schulz, R. Multifunctional intracellular matrix metalloproteinases: Implications in disease. FEBS J. 2021, 288, 7162–7182. [Google Scholar] [CrossRef] [PubMed]
- Roomi, M.W.; Kalinovsky, T.; Niedzwiecki, A.; Rath, M. Modulation of MMP-2 and -9 secretion by cytokines, inducers and inhibitors in human melanoma A-2058 cells. Oncol. Rep. 2017, 37, 3681–3687. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Adhikari, N.; Banerjee, S.; Amin, S.A.; Jha, T. Matrix metalloproteinase-9 (MMP-9) and its inhibitors in cancer: A minireview. Eur. J. Med. Chem. 2020, 194, 112260. [Google Scholar] [CrossRef] [PubMed]
- Winer, A.; Adams, S.; Mignatti, P. Matrix Metalloproteinase Inhibitors in Cancer Therapy: Turning Past Failures into Future Successes. Mol. Cancer Ther. 2018, 17, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mustafa, S.; Koran, S.; AlOmair, L. Insights into the role of matrix metalloproteinases in cancer and its various therapeutic aspects: A review. Front. Mol. Biosci. 2022, 9, 896099. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of matrix metalloproteinases in angiogenesis and cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Redondo, P.; Lloret, P.; Idoate, M.; Inoges, S. Expression and serum levels of MMP-2 and MMP-9 during human melanoma progression. Clin. Exp. Dermatol. 2005, 30, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, U.B.; Eggert, A.A.O.; Blass, K.; Bröcker, E.B.; Becker, J.C. Expression of matrix metalloproteinases in the microenvironment of spontaneous and experimental melanoma metastases reflects the requirements for tumor formation. Cancer Res. 2003, 63, 8221–8225. [Google Scholar] [PubMed]
- Desch, A.; Strozyk, E.A.; Bauer, A.T.; Huck, V.; Niemeyer, V.; Wieland, T.; Schneider, S.W. Highly invasive melanoma cells activate the vascular endothelium via an MMP-2/integrin αvβ5-induced secretion of VEGF-A. Am. J. Pathol. 2012, 181, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Leight, J.L.; Tokuda, E.Y.; Jones, C.E.; Lin, A.J.; Anseth, K.S. Multifunctional bioscaffolds for 3D culture of melanoma cells reveal increased MMP activity and migration with BRAF kinase inhibition. Proc. Natl. Acad. Sci. USA 2015, 112, 5366–5371. [Google Scholar] [CrossRef] [PubMed]
- Cotignola, J.; Reva, B.; Mitra, N.; Ishill, N.; Chuai, S.; Patel, A.; Shah, S.; Vanderbeek, G.; Coit, D.; Busam, K.; et al. Matrix metalloproteinase-9 (MMP-9) polymorphisms in patients with cutaneous malignant melanoma. BMC Med. Genet. 2007, 8, 10. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Moogk, D.; da Silva, I.P.; Ma, M.W.; Friedman, E.B.; de Miera, E.V.-S.; Darvishian, F.; Scanlon, P.; Perez-Garcia, A.; Pavlick, A.C.; Bhardvaj, N.; et al. Melanoma expression of matrix metalloproteinase-23 is associated with blunted tumor immunity and poor responses to immunotherapy. J. Transl. Med. 2014, 12, 342. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, Y.; Huang, L.; Yu, J. Evaluation of heparanase and matrix metalloproteinase-9 in patients with cutaneous malignant melanoma. J. Dermatol. 2012, 39, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Salemi, R.; Falzone, L.; Madonna, G.; Polesel, J.; Cinà, D.; Mallardo, D.; Ascierto, P.A.; Libra, M.; Candido, S. MMP-9 as a Candidate Marker of Response to BRAF Inhibitors in Melanoma Patients with BRAFV600E Mutation Detected in Circulating-Free DNA. Front. Pharmacol. 2018, 9, 856. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zurac, S.; Neagu, M.; Constantin, C.; Cioplea, M.; Nedelcu, R.; Bastian, A.; Popp, C.; Nichita, L.; Andrei, R.; Tebeica, T.; et al. Variations in the expression of TIMP1, TIMP2 and TIMP3 in cutaneous melanoma with regression and their possible function as prognostic predictors. Oncol. Lett. 2016, 11, 3354–3360. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wylie, S.; MacDonald, I.C.; Varghese, H.J.; Schmidt, E.E.; Morris, V.L.; Groom, A.C.; Chambers, A.F. The matrix metalloproteinase inhibitor batimastat inhibits angiogenesis in liver metastases of B16F1 melanoma cells. Clin. Exp. Metastasis 1999, 17, 111–117. [Google Scholar] [CrossRef]
- de Almeida, L.G.N.; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. Matrix Metalloproteinases: From Molecular Mechanisms to Physiology, Pathophysiology, and Pharmacology. Pharmacol. Rev. 2022, 74, 712–768. [Google Scholar] [CrossRef] [PubMed]
- Belotti, D.; Paganoni, P.; Giavazzi, R. MMP inhibitors: Experimental and clinical studies. Int. J. Biol. Mark. 1999, 14, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Sobirzhanovna, B.N. New Aspects of Pathology and Norms of Matrix Metalloproteinases. Web Med. J. Med. Pract. Nurs. 2024, 2, 34–37. Available online: https://webofjournals.com/index.php/5/article/view/925 (accessed on 2 July 2024).
- Cathcart, J.M.; Cao, J. MMP Inhibitors: Past, present and future. Front. Biosci. (Landmark Ed.) 2015, 20, 1164–1178. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Eckhardt, S.G. Development of matrix metalloproteinase inhibitors in cancer therapy. J. Natl. Cancer Inst. 2001, 93, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Marusak, C.; Bayles, I.; Ma, J.; Gooyit, M.; Gao, M.; Chang, M.; Bedogni, B. The thiirane-based selective MT1-MMP/MMP2 inhibitor ND-322 reduces melanoma tumor growth and delays metastatic dissemination. Pharmacol. Res. 2016, 113, 515–520. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Blackburn, J.S.; Rhodes, C.H.; Coon, C.I.; Brinckerhoff, C.E. RNA interference inhibition of matrix metalloproteinase-1 prevents melanoma metastasis by reducing tumor collagenase activity and angiogenesis. Cancer Res. 2007, 67, 10849–10858. [Google Scholar] [CrossRef] [PubMed]
- Marusak, C.; Thakur, V.; Li, Y.; Freitas, J.T.; Zmina, P.M.; Thakur, V.S.; Chang, M.; Gao, M.; Tan, J.; Xiao, M.; et al. Targeting Extracellular Matrix Remodeling Restores BRAF Inhibitor Sensitivity in BRAFi-resistant Melanoma. Clin. Cancer Res. 2020, 26, 6039–6050. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, F.; Zhi, J.; Zhao, R.; Sun, Y.; Wen, H.; Cai, H.; Chen, W.; Jiang, X.; Bai, R. Discovery of matrix metalloproteinase inhibitors as anti-skin photoaging agents. Eur. J. Med. Chem. 2024, 267, 116152. [Google Scholar] [CrossRef] [PubMed]
- Ågren, M.S.; Auf dem Keller, U. Matrix Metalloproteinases: How Much Can They Do? Int. J. Mol. Sci. 2020, 21, 2678. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]

| Synthetic Inhibitors |
Catalytic inhibitors
|
| Allosteric and exosite inhibitors |
| Monoclonal antibodies against a specific MMP |
| Interference RNAs |
| Recombinant TIMPs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazar, A.M.; Costea, D.O.; Popp, C.G.; Mastalier, B. Skin Malignant Melanoma and Matrix Metalloproteinases: Promising Links to Efficient Therapies. Int. J. Mol. Sci. 2024, 25, 7804. https://doi.org/10.3390/ijms25147804
Lazar AM, Costea DO, Popp CG, Mastalier B. Skin Malignant Melanoma and Matrix Metalloproteinases: Promising Links to Efficient Therapies. International Journal of Molecular Sciences. 2024; 25(14):7804. https://doi.org/10.3390/ijms25147804
Chicago/Turabian StyleLazar, Angela Madalina, Daniel Ovidiu Costea, Cristiana Gabriela Popp, and Bogdan Mastalier. 2024. "Skin Malignant Melanoma and Matrix Metalloproteinases: Promising Links to Efficient Therapies" International Journal of Molecular Sciences 25, no. 14: 7804. https://doi.org/10.3390/ijms25147804
APA StyleLazar, A. M., Costea, D. O., Popp, C. G., & Mastalier, B. (2024). Skin Malignant Melanoma and Matrix Metalloproteinases: Promising Links to Efficient Therapies. International Journal of Molecular Sciences, 25(14), 7804. https://doi.org/10.3390/ijms25147804