
The Influence of BMP6 on Serotonin and Glucose Metabolism
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Exogenous BMP6 and 5HT Have Opposite Impacts on Insulin Secretion and Glucose Metabolism in Pancreatic Endocrine Cells
2.2. Exogenous BMP6 Has a Positive Effect on Serotonin Secretion and a Negative Effect on the Expression of the 5HT2B Receptor in Pancreatic Endocrine Cells
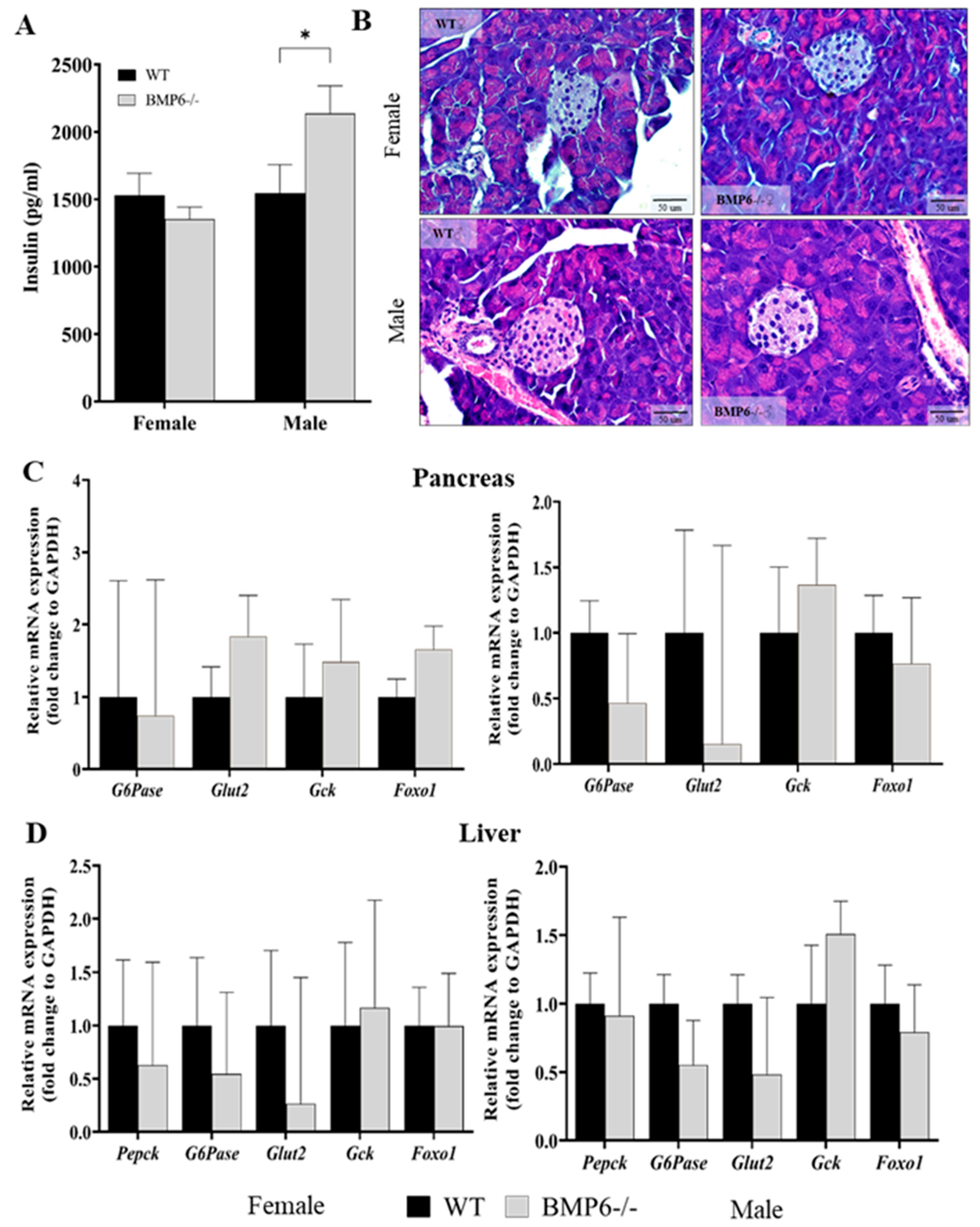
2.3. Effect of BMP6 Deficiency on Glucose Homeostasis in Mice
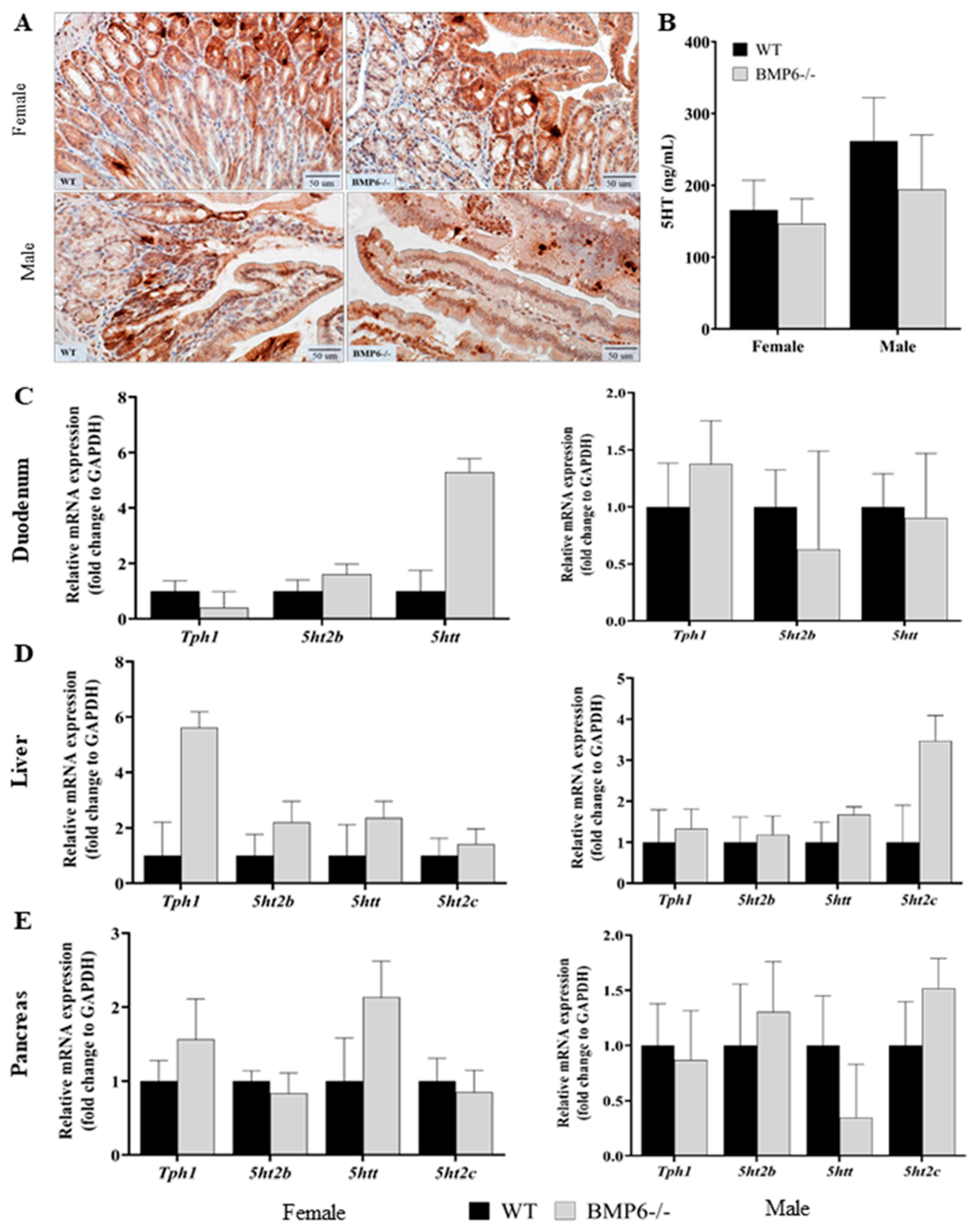
2.4. Effect of BMP6 Deficiency on Serotonin Homeostasis in Mice
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Static Insulin and Static Serotonin Secretion Assay
4.3. Animals and Animal Care
4.4. Glucose Tolerance Test (GTT) and Insulin Tolerance Test (ITT)
4.5. Gene Expression Analyses
4.6. Histological Analysis and Immunohistochemistry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patti, A.; Gennari, L.; Merlotti, D.; Dotta, F.; Nuti, R. Endocrine actions of osteocalcin. Int. J. Endocrinol. 2013, 2013, 846480. [Google Scholar] [CrossRef] [PubMed]
- Bordukalo-Niksic, T.; Kufner, V.; Vukicevic, S. The Role Of BMPs in the Regulation of Osteoclasts Resorption and Bone Remodeling: From Experimental Models to Clinical Applications. Front. Immunol. 2022, 13, 869422. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.W.; Ueharu, H.; Mishina, Y. Energy metabolism: A newly emerging target of BMP signaling in bone homeostasis. Bone 2020, 138, 115467. [Google Scholar] [CrossRef] [PubMed]
- Boon, M.R.; van den Berg, S.A.; Wang, Y.; van den Bossche, J.; Karkampouna, S.; Bauwens, M.; De Saint-Hubert, M.; van der Horst, G.; Vukicevic, S.; de Winther, M.P.; et al. BMP7 activates brown adipose tissue and reduces diet-induced obesity only at subthermoneutrality. PLoS ONE 2013, 8, e74083. [Google Scholar] [CrossRef] [PubMed]
- Grgurevic, L.; Christensen, G.L.; Schulz, T.J.; Vukicevic, S. Bone morphogenetic proteins in inflammation, glucose homeostasis and adipose tissue energy metabolism. Cytokine Growth Factor Rev. 2016, 27, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.; Alvarez-Cubela, S.; Lanzoni, G.; Vargas, N.; Prabakar, K.R.; Boulina, M.; Ricordi, C.; Inverardi, L.; Pastori, R.L.; Domínguez-Bendala, J. BMP-7 Induces Adult Human Pancreatic Exocrine-to-Endocrine Conversion. Diabetes 2015, 64, 4123–4134. [Google Scholar] [CrossRef]
- Luo, Y.; Li, L.; Xu, X.H.; Wu, T.; Yang, M.L.; Zhang, C.; Mou, H.M.; Zhou, T.T.; Jia, Y.J.; Cai, C.R.R.; et al. Decreased circulating BMP-9 levels in patients with Type 2 diabetes is a signature of insulin resistance. Clin. Sci. 2017, 131, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Grzegorzewski, K.J.; Barash, S.; Zhao, Q.; Schneider, H.; Wang, Q.; Singh, M.; Pukac, L.; Bell, A.C.; Duan, R.; et al. An integrated functional genomics screening program reveals a role for BMP-9 in glucose homeostasis. Nat. Biotechnol. 2003, 21, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Ma, C.H.; Sun, T.T.; Ren, L.P. Potential roles of bone morphogenetic protein-9 in glucose and lipid homeostasis. J. Physiol. Biochem. 2020, 76, 503–512. [Google Scholar] [CrossRef]
- Pauk, M.; Bordukalo-Niksic, T.; Brkljacic, J.; Paralkar, V.M.; Brault, A.L.; Dumic-Cule, I.; Borovecki, F.; Grgurevic, L.; Vukicevic, S. A novel role of bone morphogenetic protein 6 (BMP6) in glucose homeostasis. Acta Diabetol. 2019, 56, 365–371. [Google Scholar] [CrossRef]
- Solloway, M.J.; Dudley, A.T.; Bikoff, E.K.; Lyons, K.M.; Hogan, B.L.; Robertson, E.J. Mice lacking Bmp6 function. Dev. Genet. 1998, 22, 321–339. [Google Scholar] [CrossRef]
- Pauk, M.; Kufner, V.; Rumenovic, V.; Dumic-Cule, I.; Farkas, V.; Milosevic, M.; Bordukalo-Niksic, T.; Vukicevic, S. Iron overload in aging mice induces exocrine pancreatic injury and fibrosis due to acinar cell loss. Int. J. Mol. Med. 2021, 47, 4893. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, D.; Hannon, J.P.; Martin, G.R. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol. Biochem. Behav. 2002, 71, 533–554. [Google Scholar] [CrossRef] [PubMed]
- Yabut, J.M.; Crane, J.D.; Green, A.E.; Keating, D.J.; Khan, W.I.; Steinberg, G.R. Emerging Roles for Serotonin in Regulating Metabolism: New Implications for an Ancient Molecule. Endocr. Rev. 2019, 40, 1092–1107. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.A.; Sun, E.W.; Martin, A.M.; Keating, D.J. The ever-changing roles of serotonin. Int. J. Biochem. Cell Biol. 2020, 125, 105776. [Google Scholar] [CrossRef] [PubMed]
- Erjavec, I.; Bordukalo-Niksic, T.; Brkljacic, J.; Grcevic, D.; Mokrovic, G.; Kesic, M.; Rogic, D.; Zavadoski, W.; Paralkar, V.M.; Grgurevic, L.; et al. Constitutively Elevated Blood Serotonin Is Associated with Bone Loss and Type 2 Diabetes in Rats. PLoS ONE 2016, 11, e0150102. [Google Scholar] [CrossRef] [PubMed]
- Almaca, J.; Molina, J.; Menegaz, D.; Pronin, A.N.; Tamayo, A.; Slepak, V.; Berggren, P.O.; Caicedo, A. Human Beta Cells Produce and Release Serotonin to Inhibit Glucagon Secretion from Alpha Cells. Cell Rep. 2016, 17, 3281–3291. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Toyofuku, Y.; Lynn, F.C.; Chak, E.; Uchida, T.; Mizukami, H.; Fujitani, Y.; Kawamori, R.; Miyatsuka, T.; Kosaka, Y.; et al. Serotonin regulates pancreatic beta cell mass during pregnancy. Nat. Med. 2010, 16, 804–808. [Google Scholar] [CrossRef] [PubMed]
- Schraenen, A.; Lemaire, K.; de Faudeur, G.; Hendrickx, N.; Granvik, M.; Van Lommel, L.; Mallet, J.; Vodjdani, G.; Gilon, P.; Binart, N.; et al. Placental lactogens induce serotonin biosynthesis in a subset of mouse beta cells during pregnancy. Diabetologia 2010, 53, 2589–2599. [Google Scholar] [CrossRef]
- Paulmann, N.; Grohmann, M.; Voigt, J.P.; Bert, B.; Vowinckel, J.; Bader, M.; Skelin, M.; Jevsek, M.; Fink, H.; Rupnik, M.; et al. Intracellular Serotonin Modulates Insulin Secretion from Pancreatic β-Cells by Protein Serotonylation. PLoS Biol. 2009, 7, e1000229. [Google Scholar] [CrossRef]
- Bennet, H.; Balhuizen, A.; Medina, A.; Dekker Nitert, M.; Ottosson Laakso, E.; Essen, S.; Spegel, P.; Storm, P.; Krus, U.; Wierup, N.; et al. Altered serotonin (5-HT) 1D and 2A receptor expression may contribute to defective insulin and glucagon secretion in human type 2 diabetes. Peptides 2015, 71, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Cataldo Bascunan, L.R.; Lyons, C.; Bennet, H.; Artner, I.; Fex, M. Serotonergic regulation of insulin secretion. Acta Physiol. 2019, 225, e13101. [Google Scholar] [CrossRef] [PubMed]
- Bennet, H.; Mollet, I.G.; Balhuizen, A.; Medina, A.; Nagorny, C.; Bagge, A.; Fadista, J.; Ottosson-Laakso, E.; Vikman, P.; Dekker-Nitert, M.; et al. Serotonin (5-HT) receptor 2b activation augments glucose-stimulated insulin secretion in human and mouse islets of Langerhans. Diabetologia 2016, 59, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Ohara-Imaizumi, M.; Kim, H.; Yoshida, M.; Fujiwara, T.; Aoyagi, K.; Toyofuku, Y.; Nakamichi, Y.; Nishiwaki, C.; Okamura, T.; Uchida, T.; et al. Serotonin regulates glucose-stimulated insulin secretion from pancreatic beta cells during pregnancy. Proc. Natl. Acad. Sci. USA 2013, 110, 19420–19425. [Google Scholar] [CrossRef] [PubMed]
- Nugent, D.A.; Smith, D.M.; Jones, H.B. A review of islet of Langerhans degeneration in rodent models of type 2 diabetes. Toxicol. Pathol. 2008, 36, 529–551. [Google Scholar] [CrossRef] [PubMed]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loreal, O. A new mouse liver-specific gene, encoding a protein homologous to human antimicrobial peptide hepcidin, is overexpressed during iron overload. J. Biol. Chem. 2001, 276, 7811–7819. [Google Scholar] [CrossRef] [PubMed]
- Andriopoulos, B., Jr.; Corradini, E.; Xia, Y.; Faasse, S.A.; Chen, S.; Grgurevic, L.; Knutson, M.D.; Pietrangelo, A.; Vukicevic, S.; Lin, H.Y.; et al. BMP6 is a key endogenous regulator of hepcidin expression and iron metabolism. Nat. Genet. 2009, 41, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Salomao, M.A. Pathology of Hepatic Iron Overload. Clin. Liver Dis. 2021, 17, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.J.; McDougall, K.E.; Hou, S.C.; Tobias, J.H. Impaired growth plate function in bmp-6 null mice. Bone 2008, 42, 216–225. [Google Scholar] [CrossRef]
- Kim, R.Y.; Robertson, E.J.; Solloway, M.J. Bmp6 and Bmp7 are required for cushion formation and septation in the developing mouse heart. Dev. Biol. 2001, 235, 449–466. [Google Scholar] [CrossRef]
- Pauk, M.; Grgurevic, L.; Brkljacic, J.; Kufner, V.; Bordukalo-Niksic, T.; Grabusic, K.; Razdorov, G.; Rogic, D.; Zuvic, M.; Oppermann, H.; et al. Exogenous BMP7 corrects plasma iron overload and bone loss in Bmp6−/− mice. Int. Orthop. 2015, 39, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The Diverse Metabolic Roles of Peripheral Serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jeong, W.; Yun, C.; Kim, H.; Oh, C.M. Serotonergic Regulation of Hepatic Energy Metabolism. Endocrinol. Metab. 2021, 36, 1151–1160. [Google Scholar] [CrossRef] [PubMed]
- Meynard, D.; Kautz, L.; Darnaud, V.; Canonne-Hergaux, F.; Coppin, H.; Roth, M.P. Lack of the bone morphogenetic protein BMP6 induces massive iron overload. Nat. Genet. 2009, 41, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Jonnakuty, C.; Gragnoli, C. What do we know about serotonin? J. Cell. Physiol. 2008, 217, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Moon, J.H.; Kim, K.; Kim, H.; Kim, J.; Jeong, J.S.; Lee, J.; Kang, S.; Park, J.S.; Kim, H. beta-cell serotonin production is associated with female sex, old age, and diabetes-free condition. Biochem. Biophys. Res. Commun. 2017, 493, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Moon, J.H.; Kim, H. Serotonergic regulation of energy metabolism in peripheral tissues. J. Endocrinol. 2020, 245, R1–R10. [Google Scholar] [CrossRef] [PubMed]
- Goyvaerts, L.; Schraenen, A.; Schuit, F. Serotonin competence of mouse beta cells during pregnancy. Diabetologia 2016, 59, 1356–1363. [Google Scholar] [CrossRef]
- Guzel, T.; Mirowska-Guzel, D. The Role of Serotonin Neurotransmission in Gastrointestinal Tract and Pharmacotherapy. Molecules 2022, 27, 1680. [Google Scholar] [CrossRef]
- Gill, R.K.; Kumar, A.; Malhotra, P.; Maher, D.; Singh, V.; Dudeja, P.K.; Alrefai, W.; Saksena, S. Regulation of intestinal serotonin transporter expression via epigenetic mechanisms: Role of HDAC2. Am. J. Physiol. Cell Physiol. 2013, 304, C334–C341. [Google Scholar] [CrossRef]
- Najjar, S.A.; Hung, L.Y.; Margolis, K.G. Serotonergic Control of Gastrointestinal Development, Motility, and Inflammation. Compr. Physiol. 2023, 13, 4851–4868. [Google Scholar] [CrossRef] [PubMed]
- El-Merahbi, R.; Loffler, M.; Mayer, A.; Sumara, G. The roles of peripheral serotonin in metabolic homeostasis. FEBS Lett. 2015, 589, 1728–1734. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kosaka, Y.; Kishimoto, N.; Wang, J.; Smith, S.B.; Honig, G.; Kim, H.; Gasa, R.M.; Neubauer, N.; Liou, A.; et al. Convergence of the insulin and serotonin programs in the pancreatic beta-cell. Diabetes 2011, 60, 3208–3216. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Berglund, E.D.; Sohn, J.W.; Holland, W.L.; Chuang, J.C.; Fukuda, M.; Rossi, J.; Williams, W.; Jones, J.E.; Zigman, J.M.; et al. 5-HTRs expressed by pro-opiomelanocortin neurons regulate insulin sensitivity in liver. Nat. Neurosci. 2010, 13, 1457–1459. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhu, Y.X.; Zhou, W.B.; Gao, L.; Yuan, L.; Han, X. Serotonin Receptor 2C and Insulin Secretion. PLoS ONE 2013, 8, 54250. [Google Scholar] [CrossRef]
- Marmentini, C.; Soares, G.M.; Bronczek, G.A.; Piovan, S.; Mareze-Costa, C.E.; Carneiro, E.M.; Boschero, A.C.; Kurauti, M.A. Aging Reduces Insulin Clearance in Mice. Front. Endocrinol. 2021, 12, 679492. [Google Scholar] [CrossRef]
- Shanik, M.H.; Xu, Y.; Skrha, J.; Dankner, R.; Zick, Y.; Roth, J. Insulin resistance and hyperinsulinemia: Is hyperinsulinemia the cart or the horse? Diabetes Care 2008, 31 (Suppl. 2), S262–S268. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Arnold, A.P.; Reue, K. A Guide for the Design of Pre-clinical Studies on Sex Differences in Metabolism. Cell Metab. 2017, 25, 1216–1230. [Google Scholar] [CrossRef]
- McCullough, L.D.; de Vries, G.J.; Miller, V.M.; Becker, J.B.; Sandberg, K.; McCarthy, M.M. NIH initiative to balance sex of animals in preclinical studies: Generative questions to guide policy, implementation, and metrics. Biol. Sex Differ. 2014, 5, 15. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, B.W.; Wang, X.C.; Zha, X.J.; Sheng, C.J.; Yang, P.; Qu, S. Potential Functions of the BMP Family in Bone, Obesity, and Glucose Metabolism. J. Diabetes Res. 2021, 2021, 6707464. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, I.; Dörpholz, G.; Ott, C.E.; Kragesteen, B.; Schanze, N.; Lee, C.T.; Köhrle, J.; Mundlos, S.; Ruschke, K.; Knaus, P. BMPs as new insulin sensitizers: Enhanced glucose uptake in mature 3T3-L1 adipocytes via PPARγ and GLUT4 upregulation. Sci. Rep. 2017, 7, 17192. [Google Scholar] [CrossRef] [PubMed]
- Kousteni, S. FoxO1, the transcriptional chief of staff of energy metabolism. Bone 2012, 50, 437–443. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Ogawa, Y. Forkhead box class O family member proteins: The biology and pathophysiological roles in diabetes. J. Diabetes Investig. 2017, 8, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Altomonte, J.; Richter, A.; Harbaran, S.; Suriawinata, J.; Nakae, J.; Thung, S.N.; Meseck, M.; Accili, D.; Dong, H. Inhibition of Foxo1 function is associated with improved fasting glycemia in diabetic mice. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E718–E728. [Google Scholar] [CrossRef] [PubMed]
- Tuduri, E.; Soriano, S.; Almagro, L.; Garcia-Heredia, A.; Rafacho, A.; Alonso-Magdalena, P.; Nadal, A.; Quesada, I. The Effects of Aging on Male Mouse Pancreatic beta-Cell Function Involve Multiple Events in the Regulation of Secretion: Influence of Insulin Sensitivity. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Tuduri, E.; Soriano, S.; Almagro, L.; Montanya, E.; Alonso-Magdalena, P.; Nadal, A.; Quesada, I. The pancreatic beta-cell in ageing: Implications in age-related diabetes. Ageing Res. Rev. 2022, 80, 101674. [Google Scholar] [CrossRef]
- Ahren, B.; Pacini, G. Glucose effectiveness: Lessons from studies on insulin-independent glucose clearance in mice. J. Diabetes Investig. 2021, 12, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, C.; Fransson, U.; Hallgard, E.; Spegel, P.; Holm, C.; Krogh, M.; Warell, K.; James, P.; Mulder, H. Metabolomic and proteomic analysis of a clonal insulin-producing beta-cell line (INS-1 832/13). J. Proteome Res. 2008, 7, 400–411. [Google Scholar] [CrossRef]
- Skelin, M.; Rupnik, M.; Cencic, A. Pancreatic beta cell lines and their applications in diabetes mellitus research. ALTEX-Altern. Anim. Exp. 2010, 27, 105–113. [Google Scholar] [CrossRef]
- Asfari, M.; Janjic, D.; Meda, P.; Li, G.; Halban, P.A.; Wollheim, C.B. Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology 1992, 130, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Benede-Ubieto, R.; Estevez-Vazquez, O.; Ramadori, P.; Cubero, F.J.; Nevzorova, Y.A. Guidelines and Considerations for Metabolic Tolerance Tests in Mice. Diabetes Metab. Syndr. Obes. 2020, 13, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Sigmond, J.; Kroep, J.R.; Loves, W.; Codacci-Pisanelli, G.; Peters, G.J. Quantitative real time PCR of deoxycytidine kinase mRNA by Light Cycler PCR in relation to enzyme activity and gemcitabine sensitivity. Cancer Lett. 2004, 213, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Cardiff, R.D.; Miller, C.H.; Munn, R.J. Manual hematoxylin and eosin staining of mouse tissue sections. Cold Spring Harb. Protoc. 2014, 2014, 665–668. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milešević, M.; Matić Jelić, I.; Rumenović, V.; Ivanjko, N.; Vukičević, S.; Bordukalo-Nikšić, T. The Influence of BMP6 on Serotonin and Glucose Metabolism. Int. J. Mol. Sci. 2024, 25, 7842. https://doi.org/10.3390/ijms25147842
Milešević M, Matić Jelić I, Rumenović V, Ivanjko N, Vukičević S, Bordukalo-Nikšić T. The Influence of BMP6 on Serotonin and Glucose Metabolism. International Journal of Molecular Sciences. 2024; 25(14):7842. https://doi.org/10.3390/ijms25147842
Chicago/Turabian StyleMilešević, Marina, Ivona Matić Jelić, Viktorija Rumenović, Natalia Ivanjko, Slobodan Vukičević, and Tatjana Bordukalo-Nikšić. 2024. "The Influence of BMP6 on Serotonin and Glucose Metabolism" International Journal of Molecular Sciences 25, no. 14: 7842. https://doi.org/10.3390/ijms25147842