

Fermented Wheat Germ Protein with Histone Deacetylase Inhibitor AR42 Demonstrates Enhanced Cytotoxicity against Lymphoma Cells In Vitro and In Vivo

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Combination of AR42 with FWGP Lowers IC50 Values for AR42
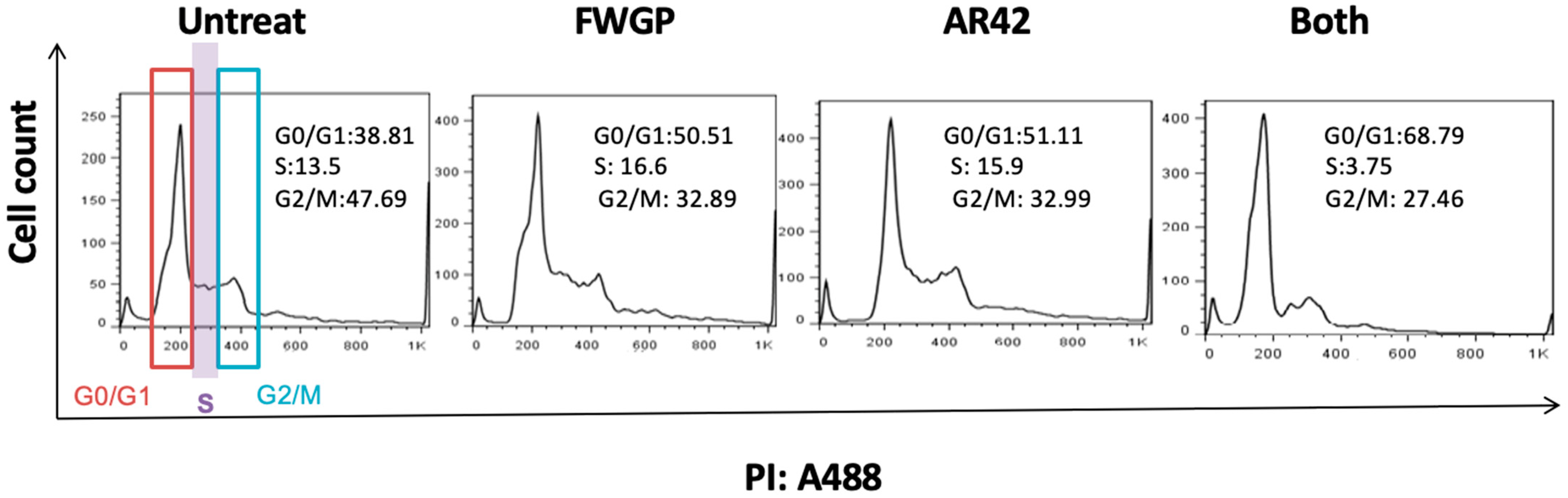
2.2. Cell Cycle Analysis in FWGP and AR42 Treated Raji Cells
2.3. In Vitro FWGP Treatment Inhibits Deacetylase Activity and Induces Cell Cycle Inhibition
2.4. Treatment of Lymphoma Xenograft Mice with FWGP + AR42 Yields Additive Tumor Growth Inhibition
3. Discussion
4. Materials and Methods
4.1. Treatments AR42 and FWGP
4.2. Cell Lines
4.3. In Vitro Cytotoxicity Assays
4.4. Flow Cytometry (FACS) for Cell Cycle Analysis
4.5. Analysis of Cellular Cytotoxicity Mechanisms
4.6. RPPA Analysis and Western Blot
4.7. In Vivo Efficacy and Toxicity in Lymphoma Xenograft Model
4.8. Statistical Analysis
4.9. Ethics
5. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poletto, S.; Novo, M.; Paruzzo, L.; Frascione, P.M.M.; Vitolo, U. Treatment strategies for patients with diffuse large B-cell lymphoma. Cancer Treat. Rev. 2022, 110, 102443. [Google Scholar] [CrossRef] [PubMed]
- Casulo, C.; Larson, M.C.; Lunde, J.J.; Habermann, T.M.; Lossos, I.S.; Wang, Y.; Nastoupil, L.J.; Strouse, C.; Chihara, D.; Martin, P.; et al. Treatment patterns and outcomes of patients with relapsed or refractory follicular lymphoma receiving three or more lines of systemic therapy (LEO CReWE): A multicentre cohort study. Lancet Haematol. 2022, 9, e289–e300. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, N.; Bond, D.; Jaglowski, S. CAR T-cell therapy for B-cell lymphoma. Curr. Probl. Cancer 2022, 46, 100826. [Google Scholar] [CrossRef] [PubMed]
- Barisone, G.A.; O’donnell, R.T.; Ma, Y.; Abuhay, M.W.; Lundeberg, K.; Gowda, S.; Tuscano, J.M. A purified, fermented, extract of Triticum aestivum has lymphomacidal activity mediated via natural killer cell activation. PLoS ONE 2018, 13, e0190860. [Google Scholar] [CrossRef] [PubMed]
- Illmer, C.; Madlener, S.; Horvath, Z.; Saiko, P.; Losert, A.; Herbacek, I.; Grusch, M.; Krupitza, G.; Fritzer-Szekeres, M.; Szekeres, T. Immunologic and Biochemical Effects of the Fermented Wheat Germ Extract Avemar. Exp. Biol. Med. 2005, 230, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Otto, C.; Hahlbrock, T.; Eich, K.; Karaaslan, F.; Jürgens, C.; Germer, C.-T.; Wiegering, A.; Kämmerer, U. Antiproliferative and antimetabolic effects behind the anticancer property of fermented wheat germ extract. BMC Complement. Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Saiko, P.; Ozsvar-Kozma, M.; Graser, G.; Lackner, A.; Grusch, M.; Madlener, S.; Krupitza, G.; Jaeger, W.; Hidvegi, M.; Agarwal, R.P.; et al. Avemar, a nontoxic fermented wheat germ extract, attenuates the growth of sensitive and 5-FdUrd/Ara-C cross-resistant H9 human lymphoma cells through induction of apoptosis. Oncol. Rep. 2009, 21, 787–791. [Google Scholar] [PubMed]
- Mueller, T.; Voigt, W. Fermented wheat germ extract—Nutritional supplement or anticancer drug? Nutr. J. 2011, 10, 89. [Google Scholar] [CrossRef]
- Tai, C.-J.C.-J.; Wang, W.-C.; Wang, C.-K.; Wu, C.-H.; Yang, M.-D.; Chang, Y.-J.; Jian, J.-Y.; Tai, C.-J. Fermented wheat germ extract induced cell death and enhanced cytotoxicity of cisplatin and 5-Fluorouracil on human hepatocellular carcinoma cells. Evidence-based complement. Altern. Med. Suppl. 2013, 2013, 121725. [Google Scholar] [CrossRef]
- Comin-Anduix, B.; Boros, L.G.G.; Marin, S.; Boren, J.; Callol-Massot, C.; Centelles, J.J.J.; Torres, J.L.L.; Agell, N.; Bassilian, S.; Cascante, M.; et al. Fermented wheat germ extract inhibits glycolysis/pentose cycle enzymes and induces apoptosis through poly(ADP-ribose) polymerase activation in Jurkat T-cell leukemia tumor cells. J. Biol. Chem. 2002, 277, 46408–46414. [Google Scholar] [CrossRef]
- Saiko, P.; Ozsvar-Kozma, M.; Madlener, S.; Bernhaus, A.; Lackner, A.; Grusch, M.; Horvath, Z.; Krupitza, G.; Jaeger, W.; Ammer, K.; et al. Avemar, a nontoxic fermented wheat germ extract, induces apoptosis and inhibits ribonucleotide reductase in human HL-60 promyelocytic leukemia cells. Cancer Lett. 2007, 250, 323–328. [Google Scholar] [CrossRef]
- Fajka-Boja, R.; Hidvegi, M.; Shoenfeld, Y.; Ion, G.; Demydenko, D.; Tomoskozi-Farkas, R.; Vizler, C.; Telekes, A.; Resetar, A.; Monostori, E. Fermented wheat germ extract induces apoptosis and downregulation of major histocompatibility complex class I proteins in tumor T and B cell lines. Int. J. Oncol. 2002, 20, 563–570. [Google Scholar] [CrossRef]
- Wang, C.-W.; Wang, C.-K.; Chang, Y.-J.; Choong, C.-Y.; Lin, C.-S.; Tai, C.-J. Preclinical evaluation on the tumor suppression efficiency and combination drug effects of fermented wheat germ extract in human ovarian carcinoma cells. Evid.-Based Complement. Altern. Med. 2015, 2015, 570785. [Google Scholar] [CrossRef] [PubMed]
- Levis, D.J.; Meckler, J.F.; O’donnell, R.T.; Tuscano, J.M. A Fermented Wheat Germ Extract Contains Protein Components Active against NSCLC Xenografts In Vivo. Curr. Issues Mol. Biol. 2023, 45, 7087–7096. [Google Scholar] [CrossRef]
- McClure, J.J.; Li, X.; Chou, C.J. Advances and Challenges of HDAC Inhibitors in Cancer Therapeutics. Adv. Cancer Res. 2018, 138, 183–211. [Google Scholar]
- Zhang, S.; Suvannasankha, A.; Crean, C.D.; White, V.L.; Chen, C.S.; Farag, S.S. The novel histone deacetylase inhibitor, AR-42, inhibits gp130/Stat3 pathway and induces apoptosis and cell cycle arrest in multiple myeloma cells. Int. J. Cancer 2011, 129, 204–213. [Google Scholar] [CrossRef]
- Lucas, D.M.; Alinari, L.; West, D.A.; Davis, M.E.; Edwards, R.B.; Johnson, A.J.; Blum, K.A.; Hofmeister, C.C.; Freitas, M.A.; Parthun, M.R.; et al. The novel deacetylase inhibitor AR-42 demonstrates pre-clinical activity in B-Cell malignancies In Vitro and In Vivo. PLoS ONE 2010, 5, e10941. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, M.J.; Tangutur, A.D.; Manyam, R.R. Epigenetic modulation and understanding of HDAC inhibitors in cancer therapy. Life Sci. 2021, 277, 119504. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Xu, B.; Yao, Y.; Yu, X.; Shen, J. The novel HDAC inhibitor AR-42-induced anti-colon cancer cell activity is associated with ceramide production. Biochem. Biophys. Res. Commun. 2015, 463, 545–550. [Google Scholar] [CrossRef]
- Booth, L.; Roberts, J.L.; Sander, C.; Lee, J.; Kirkwood, J.M.; Poklepovic, A.; Dent, P. The HDAC inhibitor AR42 interacts with pazopanib to kill trametinib/dabrafenib-resistant melanoma cells in vitro and in vivo. Oncotarget 2017, 8, 16367–16386. [Google Scholar] [CrossRef]
- Li, D.R.; Zhang, H.; Peek, E.; Wang, S.; Du, L.; Li, G.; Chin, A.I. Synergy of Histone-Deacetylase Inhibitor AR-42 with Cisplatin in Bladder Cancer. J. Urol. 2015, 194, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Sborov, D.W.; Canella, A.; Hade, E.M.; Mo, X.; Khountham, S.; Wang, J.; Ni, W.; Poi, M.; Coss, C.; Liu, Z.; et al. A phase 1 trial of the HDAC inhibitor AR-42 in patients with multiple myeloma and T- and B-cell lymphomas. Leuk. Lymphoma 2017, 58, 2310–2318. [Google Scholar] [CrossRef] [PubMed]
- Liva, S.G.; Coss, C.C.; Wang, J.; Blum, W.; Klisovic, R.; Bhatnagar, B.; Walsh, K.; Geyer, S.; Zhao, Q.; Garzon, R.; et al. Phase I study of AR-42 and decitabine in acute myeloid leukemia. Leuk. Lymphoma 2020, 61, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Welling, D.B.; Collier, K.A.; Burns, S.S.; Oblinger, J.L.; Shu, E.; Miles-Markley, B.A.; Hofmeister, C.C.; Makary, M.S.; Slone, W.; Blakeley, J.O.; et al. Early phase clinical studies of AR-42, a histone deacetylase inhibitor, for neurofibromatosis type 2-associated vestibular schwannomas and meningiomas. Laryngoscope Investig. Otolaryngol. 2021, 6, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Ny, L.; Jespersen, H.; Karlsson, J.; Alsén, S.; Filges, S.; All-Eriksson, C.; Andersson, B.; Carneiro, A.; Helgadottir, H.; Levin, M.; et al. The PEMDAC phase 2 study of pembrolizumab and entinostat in patients with metastatic uveal melanoma. Nat. Commun. 2021, 12, 5155. [Google Scholar] [CrossRef]
- Connolly, R.M.; Zhao, F.; Miller, K.D.; Lee, M.J.; Piekarz, R.L.; Smith, K.L.; Brown-Glaberman, U.A.; Winn, J.S.; Faller, B.A.; Ontilio, A.A.; et al. E2112: Randomized Phase III Trial of Endocrine Therapy Plus Entinostat or Placebo in Hormone Receptor-Positive Advanced Breast Cancer. A Trial of the ECOG-ACRIN Cancer Research Group. J. Clin. Oncol. 2021, 39, 3171–3181. [Google Scholar] [CrossRef] [PubMed]
- Canella, A.; Nieves, H.C.; Sborov, D.W.; Cascione, L.; Radomska, H.S.; Smith, E.; Stiff, A.; Consiglio, J.; Caserta, E.; Rizzotto, L.; et al. HDAC inhibitor AR-42 decreases CD44 expression and sensitizes myeloma cells to lenalidomide. Oncotarget 2015, 6, 31134–31150. [Google Scholar] [CrossRef]
- Kong, Y.; Barisone, G.A.; Sidhu, R.S.; O’donnell, R.T.; Tuscano, J.M. Efficacy of Combined Histone Deacetylase and Checkpoint Kinase Inhibition in a Preclinical Model of Human Burkitt Lymphoma. Mol. Med. 2015, 21, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Barisone, G.A.; Abuhay, M.; O’donnell, R.T.; Buksh, Z.; Yousefian, F.; Tuscano, J.M. Histone deacetylase inhibition enhances the lymphomacidal activity of the anti-CD22 monoclonal antibody HB22.7. Leuk. Res. 2014, 38, 1320–1326. [Google Scholar] [CrossRef]
- Guzman, M.L.; Yang, N.; Sharma, K.K.; Balys, M.; Corbett, C.A.; Jordan, C.T.; Becker, M.W.; Steidl, U.; Abdel-Wehab, O.; Levine, R.L.; et al. Selective activity of the histone deacetylase inhibitor AR-42 against leukemia stem cells: A novel potential strategy in acute myelogenous leukemia. Mol. Cancer Ther. 2014, 13, 1979–1990. [Google Scholar] [CrossRef]
- Yang, Y.-T.; Balch, C.; Kulp, S.K.; Mand, M.R.; Nephew, K.P.; Chen, C.-S. A rationally designed histone deacetylase inhibitor with distinct antitumor activity against ovarian cancer. Neoplasia 2009, 11, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Batlevi, C.L.; Kasamon, Y.; Bociek, R.G.; Lee, P.; Gore, L.; Copeland, A.; Sorensen, R.; Ordentlich, P.; Cruickshank, S.; Kunkel, L.; et al. ENGAGE- 501: Phase II study of entinostat (SNDX-275) in relapsed and refractory Hodgkin lymphoma. Haematologica 2016, 101, 968–975. [Google Scholar] [CrossRef]
- Demidov, L.V.; Manziuk, L.V.; Kharkevitch, G.Y.; Pirogova, N.A.; Artamonova, E.V. Adjuvant fermented wheat germ extract (Avemar) nutraceutical improves survival of high-risk skin melanoma patients: A randomized, pilot, phase II clinical study with a 7-year follow-up. Cancer Biother. Radiopharm. 2008, 23, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Weitzen, R.; Epstein, N.; Oberman, B.; Shevetz, R.; Hidvegi, M.; Berger, R. Fermented Wheat Germ Extract (FWGE) as a Treatment Additive for Castration-Resistant Prostate Cancer: A Pilot Clinical Trial. Nutr. Cancer 2021, 74, 1338–1346. [Google Scholar] [CrossRef] [PubMed]
- Heimbach, J.T.; Sebestyen, G.; Semjen, G.; Kennepohl, E. Safety studies regarding a standardized extract of fermented wheat germ. Int. J. Toxicol. 2007, 26, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Boros, L.G.; Nichelatti, M.; Shoenfeld, Y. Fermented wheat germ extract (Avemar) in the treatment of cancer and autoimmune diseases. Ann. N. Y. Acad. Sci. 2005, 1051, 529–542. [Google Scholar] [CrossRef]
- Barisone, G.A.; Ngo, T.; Tran, M.; Cortes, D.; Shahi, M.H.; Nguyen, T.-V.; Perez-Lanza, D.; Matayasuwan, W.; Díaz, E. Role of MXD3 in proliferation of DAOY human medulloblastoma cells. PLoS ONE 2012, 7, e38508. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meckler, J.F.; Levis, D.J.; Kong, Y.; O’Donnell, R.T.; Vang, D.P.; Tuscano, J.M. Fermented Wheat Germ Protein with Histone Deacetylase Inhibitor AR42 Demonstrates Enhanced Cytotoxicity against Lymphoma Cells In Vitro and In Vivo. Int. J. Mol. Sci. 2024, 25, 7866. https://doi.org/10.3390/ijms25147866
Meckler JF, Levis DJ, Kong Y, O’Donnell RT, Vang DP, Tuscano JM. Fermented Wheat Germ Protein with Histone Deacetylase Inhibitor AR42 Demonstrates Enhanced Cytotoxicity against Lymphoma Cells In Vitro and In Vivo. International Journal of Molecular Sciences. 2024; 25(14):7866. https://doi.org/10.3390/ijms25147866
Chicago/Turabian StyleMeckler, Joshua F., Daniel J. Levis, Yanguo Kong, Robert T. O’Donnell, Daniel P. Vang, and Joseph M. Tuscano. 2024. "Fermented Wheat Germ Protein with Histone Deacetylase Inhibitor AR42 Demonstrates Enhanced Cytotoxicity against Lymphoma Cells In Vitro and In Vivo" International Journal of Molecular Sciences 25, no. 14: 7866. https://doi.org/10.3390/ijms25147866
APA StyleMeckler, J. F., Levis, D. J., Kong, Y., O’Donnell, R. T., Vang, D. P., & Tuscano, J. M. (2024). Fermented Wheat Germ Protein with Histone Deacetylase Inhibitor AR42 Demonstrates Enhanced Cytotoxicity against Lymphoma Cells In Vitro and In Vivo. International Journal of Molecular Sciences, 25(14), 7866. https://doi.org/10.3390/ijms25147866