

Effects of SRI-32743, a Novel Quinazoline Structure-Based Compound, on HIV-1 Tat and Cocaine Interaction with Norepinephrine Transporter
 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Inhibition Effects of SRI-32743 on WT hNET
2.2. Kinetic Effects of SRI-32743 on [3H]DA Uptake in WT hNET
2.3. Effects of SRI-32743 on Cocaine-Induced Dissociation of [3H]Nisoxetine Binding
2.4. SRI-32743-Mediated [3H]DA and [3H]MPP+ Efflux
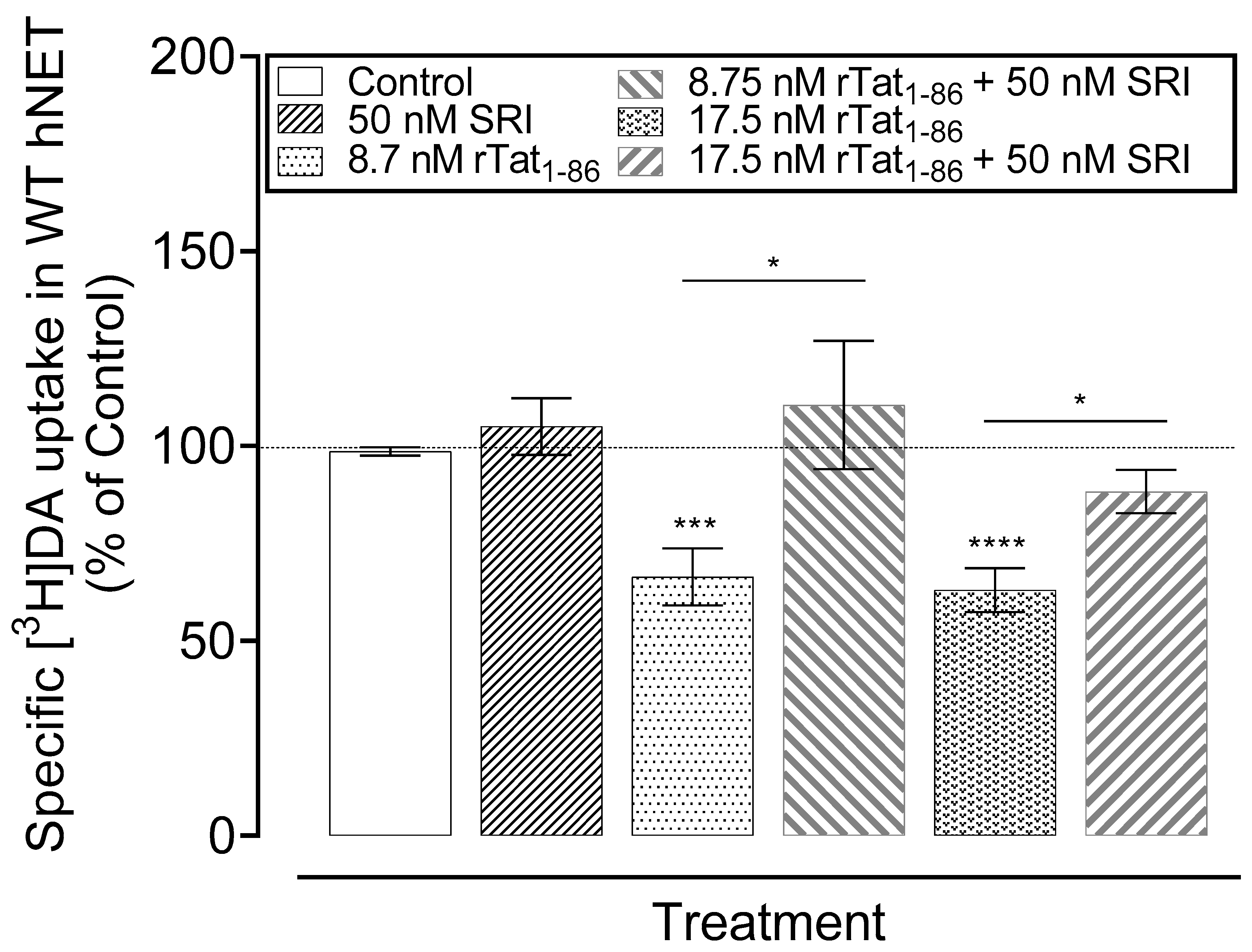
2.5. Effects of SRI-32743 on Tat-Induced Inhibition of [3H]DA Uptake via hNET
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Transfection
4.3. Competitive Inhibition [3H]DA Uptake Assay
4.4. [3H]DA Saturation Analysis
4.5. [3H]Nisoxetine Binding Assay
4.6. [3H]DA and [3H]MPP+ Efflux Assay
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Disclaimer
List of Abbreviations
| cART | Combined antiretroviral therapy |
| CHO-K1 | Chinese hamster ovary cells |
| CNS | Central nervous system |
| DA | Dopamine |
| DAT | Dopamine transporter |
| HAND | HIV-associated neurocognitive disorders |
| HIV-1 | Human immunodeficiency virus 1 |
| hDAT | Cloned human dopamine transporter |
| hNET | Cloned human norepinephrine transporter |
| NET | Norepinephrine transporter |
| SRI | Southern Research Institute |
| rTat1–86 | Recombinant HIV-1 Tat1–86 |
| RT | Room temperature |
| Tat | Transactivator of transcription |
| WT | Wild type |
| [3H]DA | Dihydroxyphenylethylamine, 3,4-[7-3H] |
| [3H]MPP+ | N-methyl-4-phenylpyridinium |
| [3H]Nisoxetine | [N-methyl-3H]Nisoxetine hydrochloride |
References
- Ambrosius, B.; Gold, R.; Chan, A.; Faissner, S. Antineuroinflammatory Drugs in HIV-Associated Neurocognitive Disorders as Potential Therapy. Neurol. Neuroimmunol. Neuroinflamm. 2019, 6, e551. [Google Scholar] [CrossRef]
- Sinharay, S.; Hammoud, D.A. Brain PET Imaging: Value for Understanding the Pathophysiology of HIV-Associated Neurocognitive Disorder (HAND). Curr. HIV/AIDS Rep. 2019, 16, 66–75. [Google Scholar] [CrossRef]
- Keng, L.D.; Winston, A.; Sabin, C.A. The Global Burden of Cognitive Impairment in People with HIV. AIDS 2023, 37, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Koutsilieri, E.; Sopper, S.; Scheller, C.; Ter Meulen, V.; Riederer, P. Involvement of Dopamine in the Progression of Aids Dementia Complex. J. Neural Transm. 2002, 109, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Shi, Z.; Liu, J.; Wang, Y. HIV Transactivator of Transcription Enhances Methamphetamine-Induced Parkinson’s-like Behavior in the Rats. NeuroReport 2014, 25, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Kesby, J.P.; Markou, A.; Semenova, S. The Effects of HIV-1 Regulatory TAT Protein Expression on Brain Reward Function, Response to Psychostimulants and Delay-Dependent Memory in Mice. Neuropharmacology 2016, 109, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, P.J.; Calderon, T.M.; Luers, A.J.; Eugenin, E.A.; Javitch, J.A.; Berman, J.W. Human Immunodeficiency Virus (HIV) Infection of Human Macrophages Is Increased by Dopamine: A Bridge between HIV-Associated Neurologic Disorders and Drug Abuse. Am. J. Pathol. 2009, 175, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.C. Tat as One Key to HIV-Induced Immune Pathogenesis and Pat Toxoid as an Important Component of a Vaccine. Proc. Natl. Acad. Sci. USA 1999, 96, 8324–8326. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.P.; Patel, K.; Johnson, K.R.; Maric, D.; Calabresi, P.A.; Hasbun, R.; Nath, A. Induction of IL-17 and Nonclassical T-Cell Activation by HIV-Tat Protein. Proc. Natl. Acad. Sci. USA 2013, 110, 13588–13593. [Google Scholar] [CrossRef]
- Ajasin, D.; Eugenin, E.A. HIV-1 Tat: Role in Bystander Toxicity. Front. Cell. Infect. Microbiol. 2020, 10, 61. [Google Scholar] [CrossRef]
- Henderson, L.J.; Johnson, T.P.; Smith, B.R.; Reoma, L.B.; Santamaria, U.A.; Bachani, M.; Demarino, C.; Barclay, R.A.; Snow, J.; Sacktor, N.; et al. Presence of Tat and Transactivation Response Element in Spinal Fluid despite Antiretroviral Therapy. AIDS 2019, 33 (Suppl. 2), S145–S157. [Google Scholar] [CrossRef]
- Scheller, C.; Arendt, G.; Nolting, T.; Antke, C.; Sopper, S.; Maschke, M.; Obermann, M.; Angerer, A.; Husstedt, I.W.; Meisner, F.; et al. Increased Dopaminergic Neurotransmission in Therapy-Naïve Asymptomatic HIV Patients Is Not Associated with Adaptive Changes at the Dopaminergic Synapses. J. Neural Transm. 2010, 117, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.M.; Fernandez, J.B.; Singer, E.J.; Commins, D.; Waldrop-Valverde, D.; Ownby, R.L.; Kumar, M. Human Immunodeficiency Virus Type 1 in the Central Nervous System Leads to Decreased Dopamine in Different Regions of Postmortem Human Brains. J. Neurovirol. 2009, 15, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.M.; Ownby, R.L.; Waldrop-Valverde, D.; Fernandez, B.; Kumar, M. Human Immunodeficiency Virus Infection in the CNS and Decreased Dopamine Availability: Relationship with Neuropsychological Performance. J. Neurovirol. 2011, 17, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Gelman, B.B.; Lisinicchia, J.G.; Chen, T.; Johnson, K.M.; Jennings, K.; Freeman, D.H.; Soukup, V.M. Prefrontal Dopaminergic and Enkephalinergic Synaptic Accommodation in HIV-Associated Neurocognitive Disorders and Encephalitis. J. Neuroimmune Pharmacol. 2012, 7, 686–700. [Google Scholar] [CrossRef]
- Wang, G.-J.; Chang, L.; Volkow, N.D.; Telang, F.; Logan, J.; Ernst, T.; Fowler, J.S. Decreased Brain Dopaminergic Transporters in HIV-Associated Dementia Patients. Brain 2004, 127, 2452–2458. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Wang, G.-J.; Volkow, N.D.; Ernst, T.; Telang, F.; Logan, J.; Fowler, J.S. Decreased Brain Dopamine Transporters Are Related to Cognitive Deficits in HIV Patients with or without Cocaine Abuse. Neuroimage 2008, 42, 869–878. [Google Scholar] [CrossRef]
- Strauss, M.; O’Donovan, B.; Ma, Y.; Xiao, Z.; Lin, S.; Bardo, M.T.; Ortinski, P.I.; McLaughlin, J.P.; Zhu, J. [3H]Dopamine Uptake through the Dopamine and Norepinephrine Transporters Is Decreased in the Prefrontal Cortex of Transgenic Mice Expressing HIV-1 Transactivator of Transcription Protein. J. Pharmacol. Exp. Ther. 2020, 374, 241–251. [Google Scholar] [CrossRef]
- Cass, W.A.; Harned, M.E.; Peters, L.E.; Nath, A.; Maragos, W.F. HIV-1 Protein Tat Potentiation of Methamphetamine-Induced Decreases in Evoked Overflow of Dopamine in the Striatum of the Rat. Brain Res. 2003, 984, 133–142. [Google Scholar] [CrossRef]
- Gaskill, P.J.; Miller, D.R.; Gamble-George, J.; Yano, H.; Khoshbouei, H. HIV, Tat and Dopamine Transmission. Neurobiol. Dis. 2017, 105, 51–73. [Google Scholar] [CrossRef]
- Midde, N.M.; Huang, X.; Gomez, A.M.; Booze, R.M.; Zhan, C.-G.; Zhu, J. Mutation of Tyrosine 470 of Human Dopamine Transporter Is Critical for HIV-1 Tat-Induced Inhibition of Dopamine Transport and Transporter Conformational Transitions. J. Neuroimmune Pharmacol. 2013, 8, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.E.; Ferris, M.J.; Ananthan, S.; Augelli-Szafran, C.E.; Zhu, J. Novel Allosteric Modulator Southern Research Institute-32743 Reverses HIV-1 Transactivator of Transcription-Induced Increase in Dopamine Release in the Caudate Putamen of Inducible Transactivator of Transcription Transgenic Mice. J. Pharmacol. Exp. Ther. 2023, 384, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Quizon, P.M.; Wang, Y.; Adeniran, C.A.; Strauss, M.J.; Jiménez-Torres, A.C.; Patel, P.; Cirino, T.J.; Eans, S.O.; Hammond, H.R.; et al. SRI-32743, a Novel Allosteric Modulator, Attenuates HIV-1 Tat Protein-Induced Inhibition of the Dopamine Transporter and Alleviates the Potentiation of Cocaine Reward in HIV-1 Tat Transgenic Mice. Neuropharmacology 2022, 220, 109239. [Google Scholar] [CrossRef] [PubMed]
- Morón, J.A.; Brockington, A.; Wise, R.A.; Rocha, B.A.; Hope, B.T. Dopamine Uptake through the Norepinephrine Transporter in Brain Regions with Low Levels of the Dopamine Transporter: Evidence from Knock-Out Mouse Lines. J. Neurosci. 2002, 22, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.J.; Porter, K.D.; Quizon, P.M.; Davis, S.E.; Lin, S.; Yuan, Y.; Martinez-Muniz, G.A.; Sun, W.-L.; Zhan, C.-G.; Zhu, J. Mutations of Tyrosine 467 in the Human Norepinephrine Transporter Attenuate HIV-1 Tat-Induced Inhibition of Dopamine Transport While Retaining Physiological Function. PLoS ONE 2022, 17, e0275182. [Google Scholar] [CrossRef] [PubMed]
- Kantor, L.; Hewlett, G.H.K.; Park, Y.H.; Richardson-Burns, S.M.; Mellon, M.J.; Gnegy, M.E. Protein Kinase C and Intracellular Calcium Are Required for Amphetamine-Mediated Dopamine Release via the Norepinephrine Transporter in Undifferentiated PC12 Cells. J. Pharmacol. Exp. Ther. 2001, 297, 1016–1024. [Google Scholar] [PubMed]
- Robertson, S.D.; Matthies, H.J.G.; Galli, A. A Closer Look at Amphetamine-Induced Reverse Transport and Trafficking of the Dopamine and Norepinephrine Transporters. Mol. Neurobiol. 2009, 39, 73–80. [Google Scholar] [CrossRef]
- Chen, N.; Justice, J.B. Cocaine Acts as an Apparent Competitive Inhibitor at the Outward-Facing Conformation of the Human Norepinephrine Transporter: Kinetic Analysis of Inward and Outward Transport. J. Neurosci. 1998, 18, 10257–10268. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.; Ringsted, K.B.; Bang-Andersen, B.; Strømgaard, K.; Kristensen, A.S. Binding Site Residues Control Inhibitor Selectivity in the Human Norepinephrine Transporter but Not in the Human Dopamine Transporter. Sci. Rep. 2015, 5, 15650. [Google Scholar] [CrossRef]
- Tatsumi, M.; Groshan, K.; Blakely, R.D.; Richelson, E. Pharmacological Profile of Antidepressants and Related Compounds at Human Monoamine Transporters. Eur. J. Pharmacol. 1997, 340, 249–258. [Google Scholar] [CrossRef]
- Eshleman, A.J.; Carmolli, M.; Cumbay, M.; Martens, C.R.; Neve, K.A.; Janowsky, A. Characteristics of Drug Interactions with Recombinant Biogenic Amine Transporters Expressed in the Same Cell Type. J. Pharmacol. Exp. Ther. 1999, 289, 877–885. [Google Scholar] [PubMed]
- Zhou, J. Norepinephine Transporter Inhibitors and Their Therapeutic Potential. Drugs Future 2004, 29, 1235. [Google Scholar] [CrossRef] [PubMed]
- Tejani-Butt, S.M.; Ordway, G.A. Effect of Age on [3H]Nisoxetine Binding to Uptake Sites for Norepinephrine in the Locus Coeruleus of Humans. Brain Res. 1992, 583, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Buck, K.J.; Amara, S.G. Chimeric Dopamine-Norepinephrine Transporters Delineate Structural Domains Influencing Selectivity for Catecholamines and 1-Methyl-4-Phenylpyridinium. Proc. Natl. Acad. Sci. USA 1994, 91, 12584–12588. [Google Scholar] [CrossRef] [PubMed]
- Buchmayer, F.; Schicker, K.; Steinkellner, T.; Geier, P.; Stübiger, G.; Hamilton, P.J.; Jurik, A.; Stockner, T.; Yang, J.-W.; Montgomery, T.; et al. Amphetamine Actions at the Serotonin Transporter Rely on the Availability of Phosphatidylinositol-4,5-Bisphosphate. Proc. Natl. Acad. Sci. USA 2013, 110, 11642–11647. [Google Scholar] [CrossRef] [PubMed]
- Sitte, H.H.; Freissmuth, M. Amphetamines, New Psychoactive Drugs and the Monoamine Transporter Cycle. Trends Pharmacol. Sci. 2015, 36, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-L.; Quizon, P.M.; Yuan, Y.; Strauss, M.J.; McCain, R.; Zhan, C.-G.; Zhu, J. Mutational Effects of Human Dopamine Transporter at Tyrosine88, Lysine92, and Histidine547 on Basal and HIV-1 Tat-Inhibited Dopamine Transport. Sci. Rep. 2019, 9, 3843. [Google Scholar] [CrossRef] [PubMed]
- Guptaroy, B.; Zhang, M.; Bowton, E.; Binda, F.; Shi, L.; Weinstein, H.; Galli, A.; Javitch, J.A.; Neubig, R.R.; Gnegy, M.E. A Juxtamembrane Mutation in the N Terminus of the Dopamine Transporter Induces Preference for an Inward-Facing Conformation. Mol. Pharmacol. 2009, 75, 514–524. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SRI-32743 (nM) | Vmax | Km |
|---|---|---|
| 0 (Control) | 0.145 ± 0.010 | 0.064 ± 0.007 |
| 5 | 0.176 ± 0.011 | 0.047 ± 0.004 |
| 50 | 0.188 ± 0.031 | 0.035 ± 0.003 * |
| 500 | 0.049 ± 0.009 * | 0.024 ± 0.009 ** |
| SRI-32743 (nM) | Vmax | Km |
|---|---|---|
| 0 (Control) | 0.700 ± 0.045 | 0.284 ± 0.101 |
| 5 | 0.616 ± 0.082 | 0.492 ± 0.289 |
| 50 | 0.559 ± 0.057 | 0.414 ± 0.200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-Torres, A.C.; Porter, K.D.; Hastie, J.A.; Adeniran, C.; Moukha-Chafiq, O.; Nguyen, T.H.; Ananthan, S.; Augelli-Szafran, C.E.; Zhan, C.-G.; Zhu, J. Effects of SRI-32743, a Novel Quinazoline Structure-Based Compound, on HIV-1 Tat and Cocaine Interaction with Norepinephrine Transporter. Int. J. Mol. Sci. 2024, 25, 7881. https://doi.org/10.3390/ijms25147881
Jiménez-Torres AC, Porter KD, Hastie JA, Adeniran C, Moukha-Chafiq O, Nguyen TH, Ananthan S, Augelli-Szafran CE, Zhan C-G, Zhu J. Effects of SRI-32743, a Novel Quinazoline Structure-Based Compound, on HIV-1 Tat and Cocaine Interaction with Norepinephrine Transporter. International Journal of Molecular Sciences. 2024; 25(14):7881. https://doi.org/10.3390/ijms25147881
Chicago/Turabian StyleJiménez-Torres, Ana Catya, Katherine D. Porter, Jamison A. Hastie, Charles Adeniran, Omar Moukha-Chafiq, Theresa H. Nguyen, Subramaniam Ananthan, Corinne E. Augelli-Szafran, Chang-Guo Zhan, and Jun Zhu. 2024. "Effects of SRI-32743, a Novel Quinazoline Structure-Based Compound, on HIV-1 Tat and Cocaine Interaction with Norepinephrine Transporter" International Journal of Molecular Sciences 25, no. 14: 7881. https://doi.org/10.3390/ijms25147881
APA StyleJiménez-Torres, A. C., Porter, K. D., Hastie, J. A., Adeniran, C., Moukha-Chafiq, O., Nguyen, T. H., Ananthan, S., Augelli-Szafran, C. E., Zhan, C.-G., & Zhu, J. (2024). Effects of SRI-32743, a Novel Quinazoline Structure-Based Compound, on HIV-1 Tat and Cocaine Interaction with Norepinephrine Transporter. International Journal of Molecular Sciences, 25(14), 7881. https://doi.org/10.3390/ijms25147881