
Threonine Deficiency Increases Triglyceride Deposition in Primary Duck Hepatocytes by Reducing STAT3 Phosphorylation
 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Influence of Thr on Activity and Fat Deposition in Hepatocytes
2.2. Thr Regulates the JAK-STAT Pathway in Primary Duck Hepatocytes
2.3. Transcriptome Analysis at Different Thr Levels
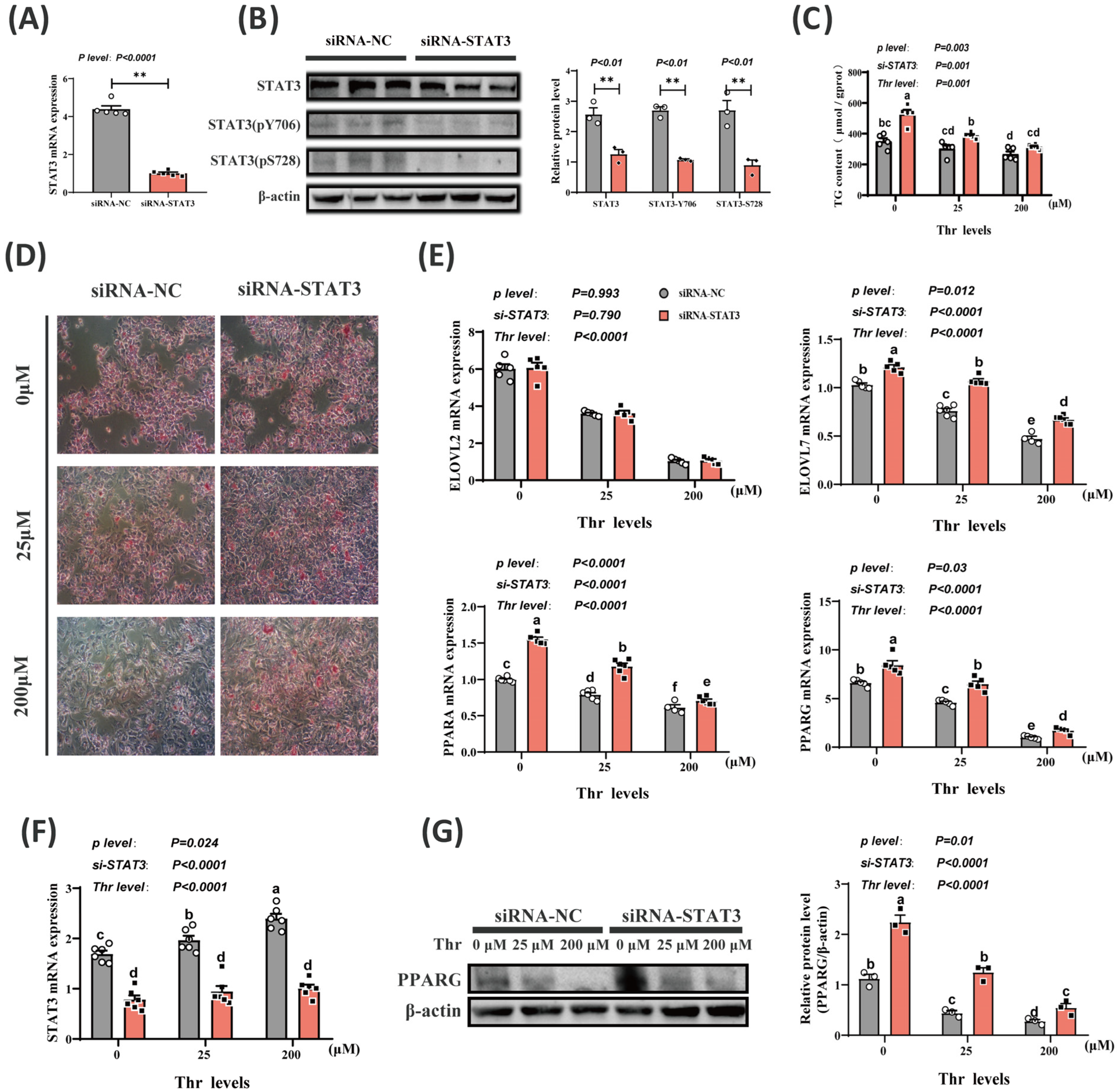
2.4. Thr Deficiency-Induced Increase in Lipid Deposition Is Caused by Reduction in STAT3 Levels
2.5. Transcriptome Analysis after STAT3 Knockdown
2.6. Thr Deficiency Promotes Triglyceride Accumulation through Inactivation of STAT3
3. Discussion
4. Materials and Methods
4.1. Isolation, Culture, and Thr Treatment of Hepatocyte
4.2. Transfection with Short Interfering RNA (siRNA) and Stattic Treatment
4.3. Cell Viability Assays
4.4. Lactate Dehydrogenase (LDH) Content
4.5. Oil Red O (ORO) Staining
4.6. Determination of Triglyceride (TG) Content
4.7. RNA Isolation, Reverse Transcription, and Quantitative Real-Time PCR
4.8. Western Blotting and Immunoprecipitation
4.9. Transcriptome Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Q.D.; Zhang, K.Y.; Zhang, Y.; Bai, S.P.; Ding, X.M.; Wang, J.P.; Peng, H.W.; Tian, G.; Xuan, Y.; Su, Z.W.; et al. Effects of dietary protein levels and protease supplementation on growth performance, carcass traits, meat quality, and standardized ileal digestibility of amino acid in Pekin ducks fed a complex diet. Poult. Sci. 2020, 99, 3557–3566. [Google Scholar] [CrossRef]
- Xie, M.; Jiang, Y.; Tang, J.; Wen, Z.G.; Zhang, Q.; Huang, W.; Hou, S.S. Effects of low-protein diets on growth performance and carcass yield of growing White Pekin ducks. Poult. Sci. 2017, 96, 1370–1375. [Google Scholar] [CrossRef]
- Wu, Y.; Tang, J.; Wen, Z.; Zhang, B.; Cao, J.; Zhao, L.; Guo, Z.; Xie, M.; Zhou, Z.; Hou, S. Dietary methionine deficiency stunts growth and increases fat deposition via suppression of fatty acids transportation and hepatic catabolism in Pekin ducks. J. Anim. Sci. Biotechnol. 2022, 13, 61. [Google Scholar] [CrossRef]
- Xia, W.G.; Abouelezz, K.F.M.; Fouad, A.M.; Chen, W.; Ruan, D.; Wang, S.; Azzam, M.M.M.; Luo, X.; Fan, Q.L.; Zhang, Y.N.; et al. Productivity, reproductive performance, and fat deposition of laying duck breeders in response to concentrations of dietary energy and protein. Poult. Sci. 2019, 98, 3729–3738. [Google Scholar] [CrossRef]
- Liu, J.B.; Yan, H.L.; Zhang, Y.; Hu, Y.D.; Zhang, H.F. Effects of dietary energy and protein content and lipid source on growth performance and carcass traits in Pekin ducks. Poult. Sci. 2019, 98, 4829–4837. [Google Scholar] [CrossRef]
- Zeng, Q.F.; Cherry, P.; Doster, A.; Murdoch, R.; Adeola, O.; Applegate, T.J. Effect of dietary energy and protein content on growth and carcass traits of Pekin ducks. Poult. Sci. 2015, 94, 384–394. [Google Scholar] [CrossRef]
- Ma, S.; Zhang, K.; Shi, S.; Li, X.; Che, C.; Chen, P.; Liu, H. Low-protein diets supplemented with isoleucine alleviate lipid deposition in broilers through activating 5′ adenosine monophosphate-activated protein kinase and janus kinase 2/signal transducer and activator of transcription 3 signaling pathways. Poult. Sci. 2023, 102, 102441. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhu, Y.W.; Xie, M.; Tang, J.; Wen, Z.G.; Qiao, S.Y.; Hou, S.S. Interactions of dietary protein and threonine on growth performance in Pekin ducklings from 1 to 14 days of age. Poult. Sci. 2018, 97, 262–266. [Google Scholar] [CrossRef]
- Yap, Y.W.; Rusu, P.M.; Chan, A.Y.; Fam, B.C.; Jungmann, A.; Solon-Biet, S.M.; Barlow, C.K.; Creek, D.J.; Huang, C.; Schittenhelm, R.B.; et al. Restriction of essential amino acids dictates the systemic metabolic response to dietary protein dilution. Nat. Commun. 2020, 11, 2894. [Google Scholar] [CrossRef]
- Chen, J.; Qian, D.; Wang, Z.; Sun, Y.; Sun, B.; Zhou, X.; Hu, L.; Shan, A.; Ma, Q. Threonine supplementation prevents the development of fat deposition in mice fed a high-fat diet. Food Funct. 2022, 13, 7772–7780. [Google Scholar] [CrossRef]
- Jiang, Y.; Tang, J.; Xie, M.; Wen, Z.G.; Qiao, S.Y.; Hou, S.S. Threonine supplementation reduces dietary protein and improves lipid metabolism in Pekin ducks. Br. Poult. Sci. 2017, 58, 687–693. [Google Scholar] [CrossRef]
- Heeren, J.; Scheja, L. Metabolic-associated fatty liver disease and lipoprotein metabolism. Mol. Metab. 2021, 50, 101238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hong, F.; Ma, C.; Yang, S. Hepatic Lipid Metabolism Disorder and Atherosclerosis. Endocr. Metab. Immune. Disord Drug Targets 2022, 22, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Kalish, B.T.; Fell, G.L.; Nandivada, P.; Puder, M. Clinically Relevant Mechanisms of Lipid Synthesis, Transport, and Storage. JPEN J. Parenter. Enteral Nutr. 2015, 39, 8S–17S. [Google Scholar] [CrossRef]
- Chang, C.C.; Sia, K.C.; Chang, J.F.; Lin, C.M.; Yang, C.M.; Huang, K.Y.; Lin, W.N. Lipopolysaccharide promoted proliferation and adipogenesis of preadipocytes through JAK/STAT and AMPK-regulated cPLA2 expression. Int. J. Med. Sci. 2019, 16, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Ashmawy, A.I.; El-Abhar, H.S.; Abdallah, D.M.; Ali, M.A. Chloroquine modulates the sulforaphane anti-obesity mechanisms in a high-fat diet model: Role of JAK-2/STAT-3/SOCS-3 pathway. Eur. J. Pharmacol. 2022, 927, 175066. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, X.; Liu, W.; Zhou, Y.; Wei, W.; Liu, D.; Wong, C.C.; Sung, J.J.Y.; Yu, J. Activated Natural Killer Cell Promotes Nonalcoholic Steatohepatitis Through Mediating JAK/STAT Pathway. Cell Mol. Gastroenterol Hepatol. 2022, 13, 257–274. [Google Scholar] [CrossRef]
- Liu, J.; Wang, F.; Luo, F. The Role of JAK/STAT Pathway in Fibrotic Diseases: Molecular and Cellular Mechanisms. Biomolecules 2023, 13, 119. [Google Scholar] [CrossRef] [PubMed]
- Pang, Q.; You, L.; Meng, X.; Li, Y.; Deng, T.; Li, D.; Zhu, B. Regulation of the JAK/STAT signaling pathway: The promising targets for cardiovascular disease. Biochem. Pharmacol. 2023, 213, 115587. [Google Scholar] [CrossRef]
- Inoue, H.; Ogawa, W.; Ozaki, M.; Haga, S.; Matsumoto, M.; Furukawa, K.; Hashimoto, N.; Kido, Y.; Mori, T.; Sakaue, H.; et al. Role of STAT-3 in regulation of hepatic gluconeogenic genes and carbohydrate metabolism in vivo. Nat. Med. 2004, 10, 168–174. [Google Scholar] [CrossRef]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: Role of signal transducer and activator of transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Cernkovich, E.R.; Deng, J.; Bond, M.C.; Combs, T.P.; Harp, J.B. Adipose-specific disruption of signal transducer and activator of transcription 3 increases body weight and adiposity. Endocrinology 2008, 149, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Dong, M.; Liu, W.; Gao, C.; Jia, Y.; Zhang, X.; Xiao, X.; Liu, Q.; Lin, H. Peripheral IL-6/STAT3 signaling promotes beiging of white fat. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119080. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhou, Y.; Lei, W.; Zhang, K.; Shi, J.; Hu, Y.; Shu, G.; Song, J. Signal transducer and activator of transcription 3 (STAT3) regulates adipocyte differentiation via peroxisome-proliferator-activated receptor gamma (PPARgamma). Biol. Cell 2009, 102, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Jin, Z.; Yi, C.O.; Oh, J.; Jeong, E.A.; Lee, J.Y.; Park, K.A.; Kim, K.E.; Lee, J.E.; Kim, H.J.; et al. Effects of lobeglitazone on insulin resistance and hepatic steatosis in high-fat diet-fed mice. PLoS ONE 2018, 13, e0200336. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Guo, W.; Yang, Y.; Wu, J. JAK2/STAT3 pathway is involved in the early stage of adipogenesis through regulating C/EBPβ transcription. J. Cell Biochem. 2011, 112, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xie, M.; Fan, W.; Xue, J.; Zhou, Z.; Tang, J.; Chen, G.; Hou, S. Transcriptome Analysis Reveals Differential Expression of Genes Regulating Hepatic Triglyceride Metabolism in Pekin Ducks During Dietary Threonine Deficiency. Front. Genet. 2019, 10, 710. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhuang, Z.; Jia, W.; Wen, Z.; Xie, M.; Bai, H.; Bi, Y.; Wang, Z.; Chang, G.; Hou, S.; et al. Proteomic and phosphoproteomic analysis reveal threonine deficiency increases hepatic lipid deposition in Pekin ducks via reducing STAT phosphorylation. Anim. Nutr. 2023, 13, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Viviani, R.; Sechi, A.M.; Lenaz, G. Lipid metabolism in fatty liver of lysine- and threonine-deficient rats. J. Lipid Res. 1966, 7, 473–478. [Google Scholar] [CrossRef]
- Zarrin-Kavyani, S.; Khatibjoo, A.; Fattahnia, F.; Taherpour, K. Effect of threonine and potassium carbonate on broiler chicken performance, immunity, carcass traits, and small intestine morphology. Trop Anim. Health Prod. 2020, 52, 943–953. [Google Scholar] [CrossRef]
- Omrani, A.H.; Mousavi, S.N.; Foroudi, F.; Jafarabadi, G.A.; Hosseini, S.A.; Alahyaribeik, S. The effects of probiotic and threonine application on the carcass yield, internal organ development, intestinal morphology and cecal microbiota of broilers challenged with Clostridium perfringens. Res. Vet. Sci. 2023, 160, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Haro, D.; Marrero, P.F.; Relat, J. Nutritional Regulation of Gene Expression: Carbohydrate-, Fat- and Amino Acid-Dependent Modulation of Transcriptional Activity. Int. J. Mol. Sci. 2019, 20, 1386. [Google Scholar] [CrossRef]
- Caiazzo, G.; Caiazzo, A.; Napolitano, M.; Megna, M.; Potestio, L.; Fornaro, L.; Parisi, M.; Luciano, M.A.; Ruggiero, A.; Testa, A.; et al. The Use of JAK/STAT Inhibitors in Chronic Inflammatory Disorders. J. Clin. Med. 2023, 12, 2865. [Google Scholar] [CrossRef] [PubMed]
- Dodington, D.W.; Desai, H.R.; Woo, M. JAK/STAT—Emerging Players in Metabolism. Trends Endocrinol Metab. 2018, 29, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Greenspan, D.S. ECM roles in the function of metabolic tissues. Trends Endocrinol. Metab. 2012, 23, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, J.; Yan, W.; Zhou, K.; Cao, Y.; Cai, W. p38α MAPK antagonizing JNK to control the hepatic fat accumulation in pediatric patients onset intestinal failure. Cell Death Dis. 2017, 8, e3110. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Wang, J. Wnt/β-Catenin Signaling and Obesity. Front. Physiol. 2018, 9, 792. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, J.; Chen, W.; Xie, X.; Chu, X.; Valencak, T.G.; Wang, Y.; Shan, T. Comprehensive evaluation of the metabolic effects of porcine CRTC3 overexpression on subcutaneous adipocytes with metabolomic and transcriptomic analyses. J. Anim. Sci. Biotechnol. 2021, 12, 19. [Google Scholar] [CrossRef]
- Oteng, A.B.; Kersten, S. Mechanisms of Action of trans Fatty Acids. Adv. Nutr. 2020, 11, 697–708. [Google Scholar] [CrossRef]
- Sato, H.; Taketomi, Y.; Ushida, A.; Isogai, Y.; Kojima, T.; Hirabayashi, T.; Miki, Y.; Yamamoto, K.; Nishito, Y.; Kobayashi, T.; et al. The adipocyte-inducible secreted phospholipases PLA2G5 and PLA2G2E play distinct roles in obesity. Cell Metab. 2014, 20, 119–132. [Google Scholar] [CrossRef]
- Sato, H.; Taketomi, Y.; Miki, Y.; Murase, R.; Yamamoto, K.; Murakami, M. Secreted Phospholipase PLA2G2D Contributes to Metabolic Health by Mobilizing ω3 Polyunsaturated Fatty Acids in WAT. Cell Rep. 2020, 31, 107579. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.F.; Tocher, D.R.; Monroig, O. Long-chain polyunsaturated fatty acid biosynthesis in chordates: Insights into the evolution of Fads and Elovl gene repertoire. Prog. Lipid Res. 2016, 62, 25–40. [Google Scholar] [CrossRef]
- Zhu, K.C.; Song, L.; Guo, H.Y.; Guo, L.; Zhang, N.; Liu, B.S.; Jiang, S.G.; Zhang, D.C. Elovl4a participates in LC-PUFA biosynthesis and is regulated by PPARαβ in golden pompano Trachinotus ovatus (Linnaeus 1758). Sci. Rep. 2019, 9, 4684. [Google Scholar] [CrossRef]
- Chan, P.C.; Liao, M.T.; Hsieh, P.S. The Dualistic Effect of COX-2-Mediated Signaling in Obesity and Insulin Resistance. Int. J. Mol. Sci. 2019, 20, 3115. [Google Scholar] [CrossRef]
- Zhen, Y.; Wu, Q.; Ding, Y.; Zhang, W.; Zhai, Y.; Lin, X.; Weng, Y.; Guo, R.; Zhang, Y.; Feng, J.; et al. Exogenous hydrogen sulfide promotes hepatocellular carcinoma cell growth by activating the STAT3-COX-2 signaling pathway. Oncol. Lett. 2018, 15, 6562–6570. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Y.; Liu, N.; Chen, J.; Guo, L.; Dai, Z.; Wang, C.; Wu, Z.; Wu, G. Dietary L-arginine supplementation reduces lipid accretion by regulating fatty acid metabolism in Nile tilapia (Oreochromis niloticus). J. Anim. Sci. Biotechnol. 2020, 11, 82. [Google Scholar] [CrossRef]
- Anghel, S.I.; Wahli, W. Fat poetry: A kingdom for PPAR gamma. Cell Res. 2007, 17, 486–511. [Google Scholar] [CrossRef]
- Janani, C.; Ranjitha Kumari, B.D. PPAR gamma gene—A review. Diabetes Metab. Syndr. 2015, 9, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Bauters, D.; Scroyen, I.; Van Hul, M.; Lijnen, H.R. Gelatinase A (MMP-2) promotes murine adipogenesis. Biochim. Biophys. Acta 2015, 1850, 1449–1456. [Google Scholar] [CrossRef]
- Waldron, A.L.; Schroder, P.A.; Bourgon, K.L.; Bolduc, J.K.; Miller, J.L.; Pellegrini, A.D.; Dubois, A.L.; Blaszkiewicz, M.; Townsend, K.L.; Rieger, S. Oxidative stress-dependent MMP-13 activity underlies glucose neurotoxicity. J. Diabetes Complicat. 2018, 32, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Zhang, H.; Zhang, S. Toll-like receptor 2 mediates deposition of collagen I in adipose tissue of high fat diet-induced obese mice. Mol. Med. Rep. 2018, 17, 5958–5963. [Google Scholar] [CrossRef] [PubMed]
- Hui, W.; Litherland, G.J.; Elias, M.S.; Kitson, G.I.; Cawston, T.E.; Rowan, A.D.; Young, D.A. Leptin produced by joint white adipose tissue induces cartilage degradation via upregulation and activation of matrix metalloproteinases. Ann. Rheum. Dis. 2012, 71, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.C. Importance of plasma matrix metalloproteinases (MMP) and tissue inhibitors of metalloproteinase (TIMP) in development of fibrosis in agnogenic myeloid metaplasia. Leuk Lymphoma 2005, 46, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Mitrut, R.; Stepan, A.E.; Margaritescu, C.; Andreiana, B.C.; Kesse, A.M.; Simionescu, C.E.; Militaru, C. Immunoexpression of MMP-8, MMP-9 and TIMP-2 in dilated cardiomyopathy. Rom. J. Morphol. Embryol. 2019, 60, 119–124. [Google Scholar] [PubMed]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuna, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef] [PubMed]
- Naim, A.; Pan, Q.; Baig, M.S. Matrix Metalloproteinases (MMPs) in Liver Diseases. J. Clin. Exp. Hepatol. 2017, 7, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chen, J.; Sun, H.; Zhang, Y.; Zou, D. New insights into fibrosis from the ECM degradation perspective: The macrophage-MMP-ECM interaction. Cell Biosci. 2022, 12, 117. [Google Scholar]
- Chen, J.Y.; Wu, Y.P.; Li, C.Y.; Jheng, H.F.; Kao, L.Z.; Yang, C.C.; Leu, S.Y.; Lien, I.C.; Weng, W.T.; Tai, H.C.; et al. PPARγ activation improves the microenvironment of perivascular adipose tissue and attenuates aortic stiffening in obesity. J. Biomed. Sci. 2021, 28, 22. [Google Scholar] [CrossRef] [PubMed]
- Bao, Q.; Wang, L.; Hu, X.; Yuan, C.; Zhang, Y.; Chang, G.; Chen, G. Developmental Changes of Duckling Liver and Isolation of Primary Hepatocytes. Animals 2023, 13, 1820. [Google Scholar] [CrossRef]
- Schust, J.; Sperl, B.; Hollis, A.; Mayer, T.U.; Berg, T. Stattic: A small-molecule inhibitor of STAT3 activation and dimerization. Chem. Biol. 2006, 13, 1235–1242. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes 1 | Sequence (5′-3′) | Product Length (5′-3′) |
|---|---|---|
| PPARA | F: ACCAGCATCCAGTCCTTCATCCA | 146 bp |
| R: AACCTTCACAAGCATGTACTCCGTAA | ||
| PPARG | F: CCCAAGTTTGAGTTCGCTGT | 192 bp |
| R: GCTGTGACGACTCTGGATGA | ||
| ELOVL2 | F: CCAAGGTGCTGTGGTGGTAT | 162 bp |
| R: CGCAGGGTATCCAGTTCAGG | ||
| ELOVL7 | F: AGCACTGGTTACCTTGCCTC | 174 bp |
| R: GCGTGTGTGCCCTTAACAA | ||
| PDGFRB | F: CCACCCATGCCTCCGATGAAATC | 131 bp |
| R: CCTCTTCCTGTAGCAATCCACCAAG | ||
| IL-6 | F: CGTGTGCGAGAACAGCATG | 364 bp |
| R: GTCTCGGAGGATGAGGTG | ||
| JAK1 | F: CTGTGCAGATACGATCCAGAAGGTG | 101 bp |
| R: TTCTTGAGGTCAGCGATGTGATTCC | ||
| JAK2 | F: ACCTATTTGCACAGTGGCGAGATG | 139 bp |
| R: AGTGGTGTTTGGTCCCTTTCTTTGG | ||
| TYK2 | F: TCAACATCGGCAAGGACACCAAC | 80 bp |
| R: CTCGTTCATCCCGTGCCAGTTC | ||
| STAT1 | F: ACGCAGGAAGCAGAACGAATGAG | 139 bp |
| R: TTTGAGATCACGACAATGGGAAGGG | ||
| STAT3 | F: AGGAGGAGGCGTTTGGGAAGTAC | 135 bp |
| R: CGATGGTGTTGCTGAAGGAGGTG | ||
| STAT5B | F: TCACCGACATCATCTCTGCCCT | 212 bp |
| R: ATTGCGGGTGCTTTCGTTCTT | ||
| MMP1 | F: GCCTACACGGACCCCAATGA | 195 bp |
| R: CAGCATGTATCTGCCCTTGAAGA | ||
| MMP13 | F: ATGGAAGCAGGCTACCCCAG | 162 bp |
| R: AGTGTGCAGGACACGGACAA | ||
| TIMP4 | F: TCTGCGATTCTGCTTTAGTGAT | 354 bp |
| R: TACAGCCACAGCCCATTTGA | ||
| PTGS2 | F: GAAAAACCACGACCAGGTGC | 198 bp |
| R: ACAGCCTTTCACGTTGTTGC | ||
| β-actin | F: ATGTCGCCCTGGATTTCG | 165 bp |
| R: CACAGGACTCCATACCCAAGAA |
| Antibodies 1 | Cat No. | Source | Dilution of WB |
|---|---|---|---|
| STAT3 | A1192 | ABclonal, Wuhan, China | 1:1000 |
| p-STAT3(Y706) | bs-22386R | Bioss, Beijing, China | 1:1000 |
| p-STAT3(S728) | AF3294 | Affinity, Cincinnati, OH, USA | 1:1000 |
| PPARG | ab178860 | abcam, Cambridge, UK | 1:1000 |
| β-actin | NB600-532 | Novus, Colorado, USA | 1:5000 |
| HRP-labeled Goat Anti-rabbit IgG(H+L) | ab205718 | abcam, Cambridge, UK | 1:10,000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuang, Z.; Jia, W.; Wu, L.; Li, Y.; Lu, Y.; Xu, M.; Bai, H.; Bi, Y.; Wang, Z.; Chen, S.; et al. Threonine Deficiency Increases Triglyceride Deposition in Primary Duck Hepatocytes by Reducing STAT3 Phosphorylation. Int. J. Mol. Sci. 2024, 25, 8142. https://doi.org/10.3390/ijms25158142
Zhuang Z, Jia W, Wu L, Li Y, Lu Y, Xu M, Bai H, Bi Y, Wang Z, Chen S, et al. Threonine Deficiency Increases Triglyceride Deposition in Primary Duck Hepatocytes by Reducing STAT3 Phosphorylation. International Journal of Molecular Sciences. 2024; 25(15):8142. https://doi.org/10.3390/ijms25158142
Chicago/Turabian StyleZhuang, Zhong, Wenqian Jia, Lei Wu, Yongpeng Li, Yijia Lu, Minghong Xu, Hao Bai, Yulin Bi, Zhixiu Wang, Shihao Chen, and et al. 2024. "Threonine Deficiency Increases Triglyceride Deposition in Primary Duck Hepatocytes by Reducing STAT3 Phosphorylation" International Journal of Molecular Sciences 25, no. 15: 8142. https://doi.org/10.3390/ijms25158142
APA StyleZhuang, Z., Jia, W., Wu, L., Li, Y., Lu, Y., Xu, M., Bai, H., Bi, Y., Wang, Z., Chen, S., Jiang, Y., & Chang, G. (2024). Threonine Deficiency Increases Triglyceride Deposition in Primary Duck Hepatocytes by Reducing STAT3 Phosphorylation. International Journal of Molecular Sciences, 25(15), 8142. https://doi.org/10.3390/ijms25158142