
Use of Residual Lignocellulosic Biomass and Algal Biomass to Produce Biofuels

 , , , and
, , , and 
Abstract
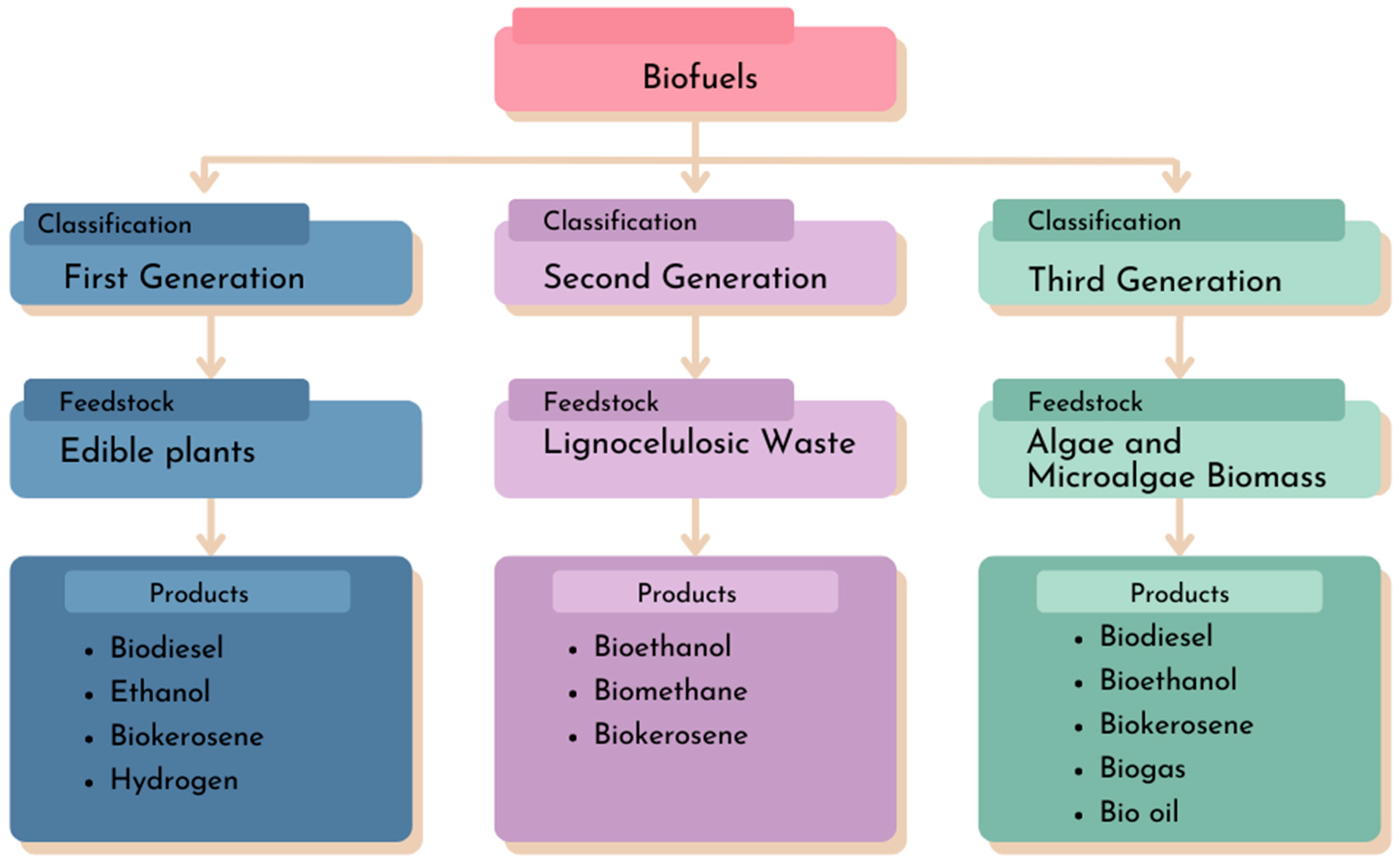
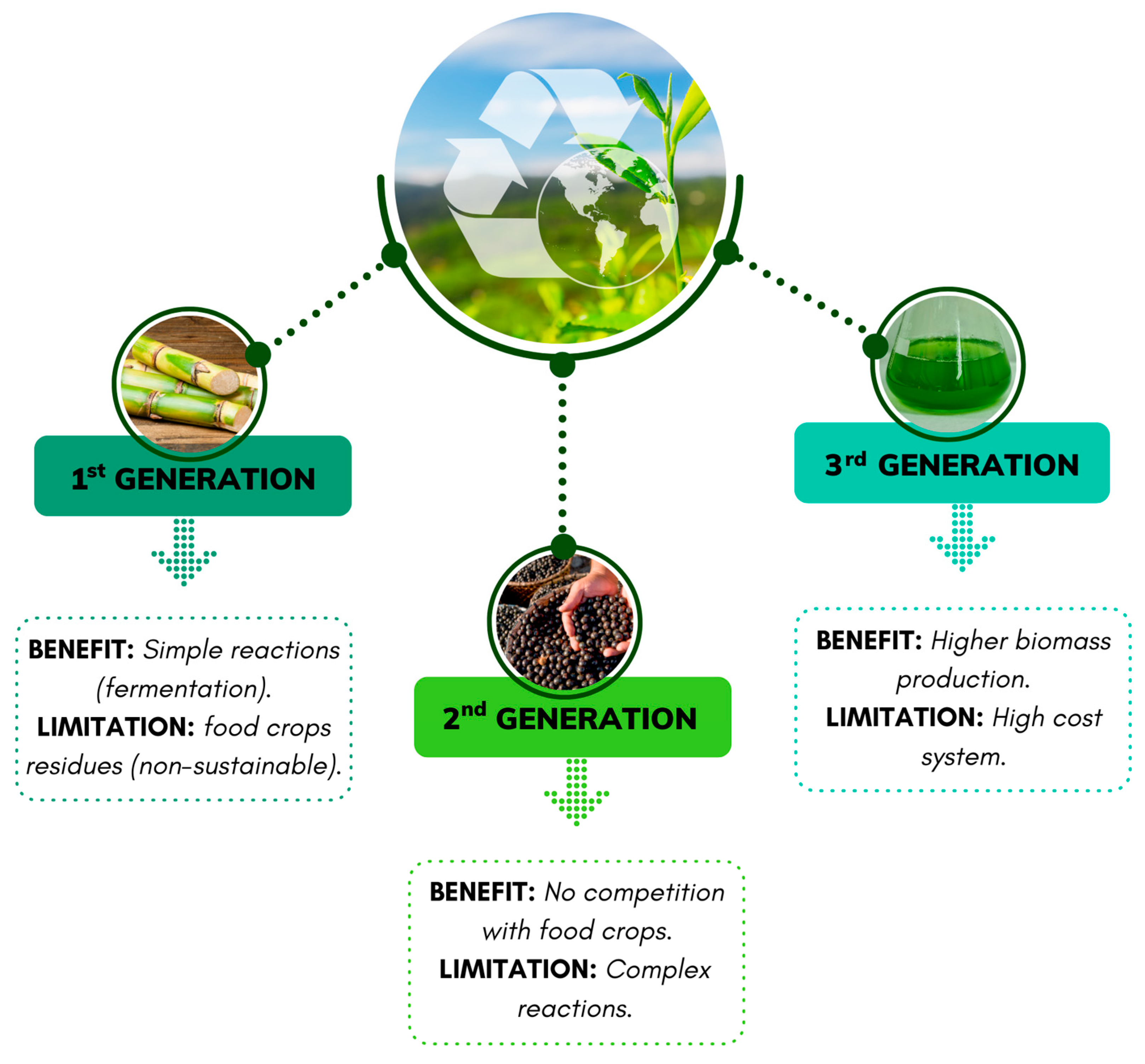
1. Introduction
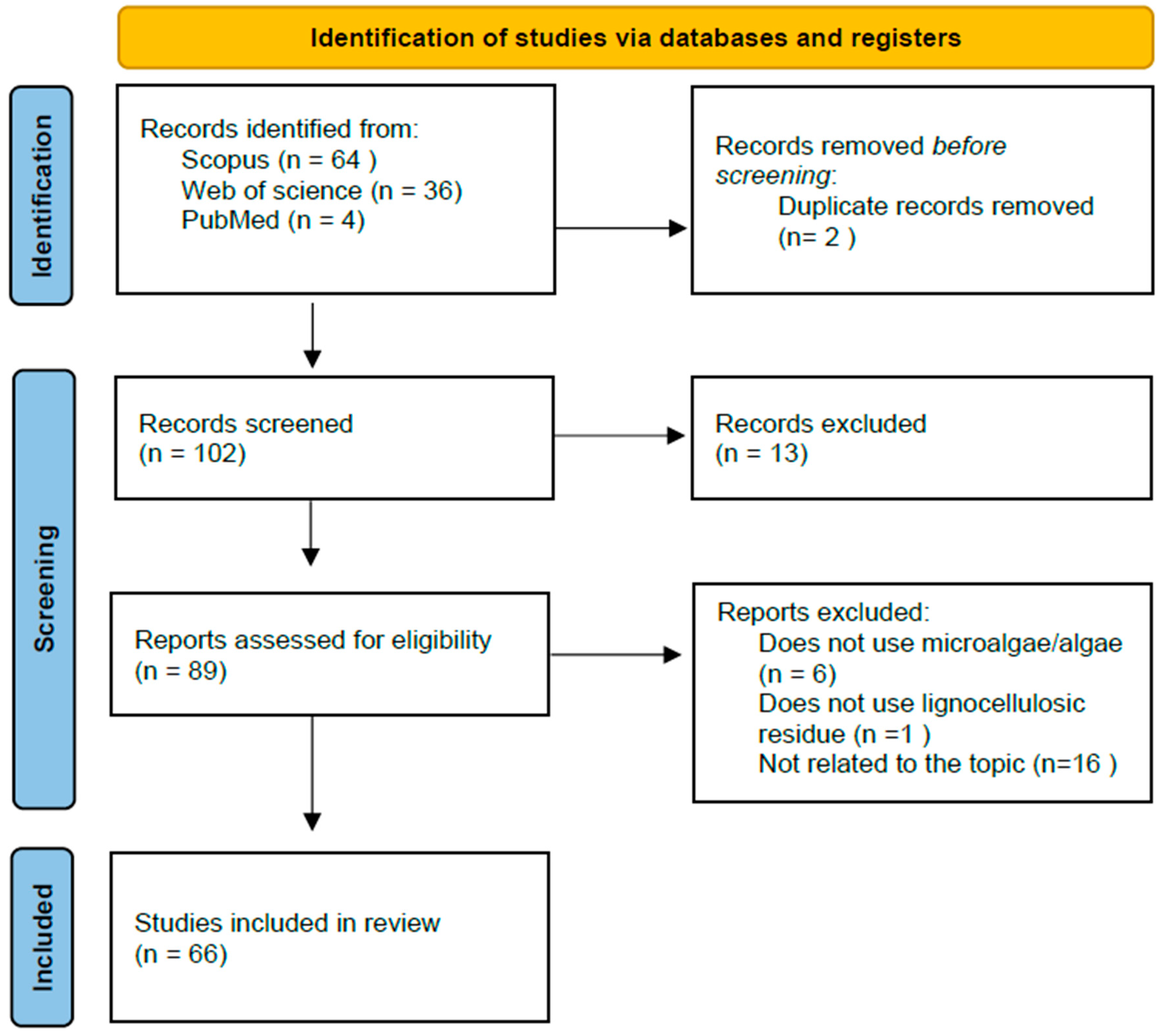
2. Research Methodology
3. Results
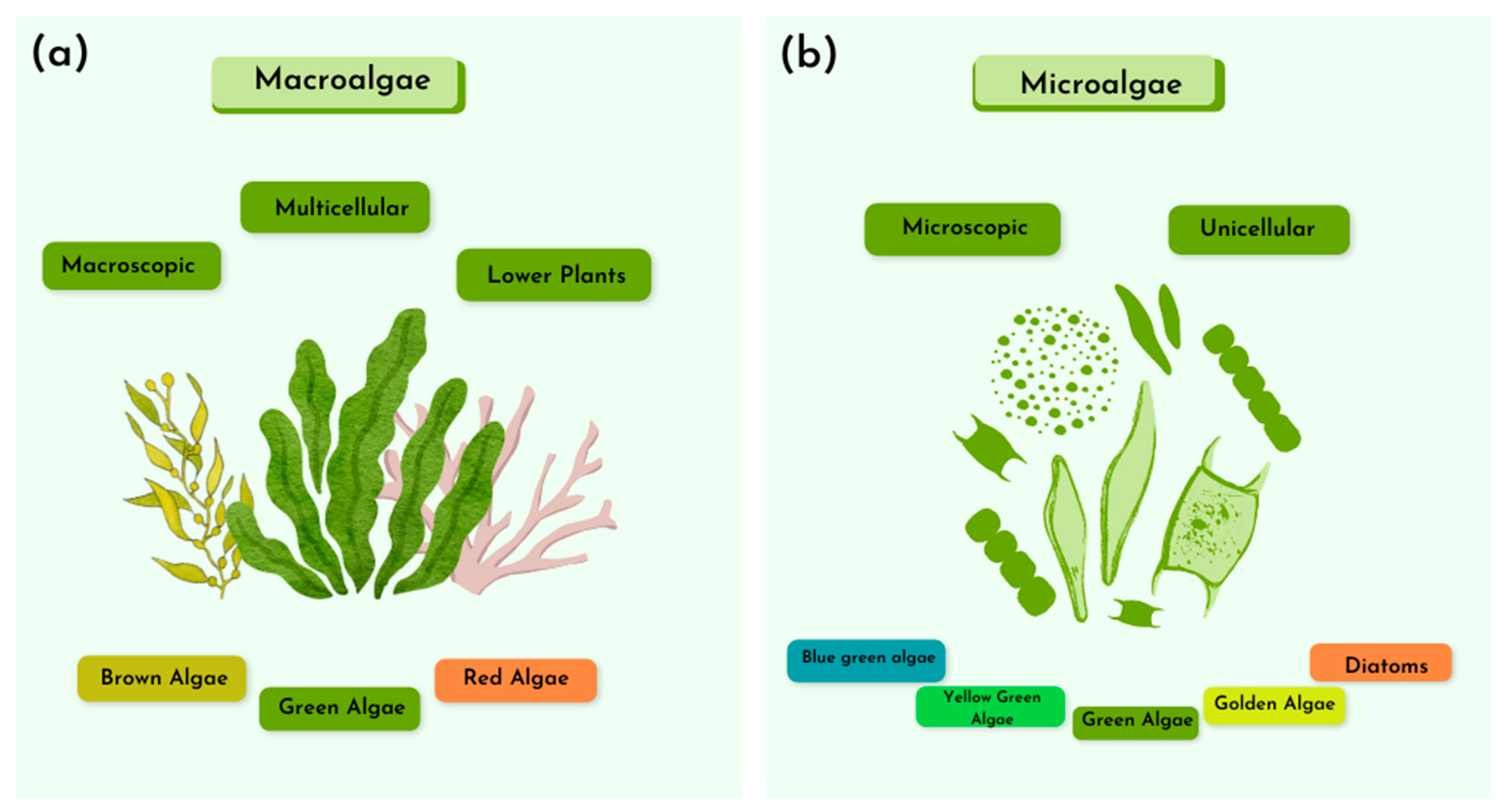
3.1. Algal and Microalgal Biomass
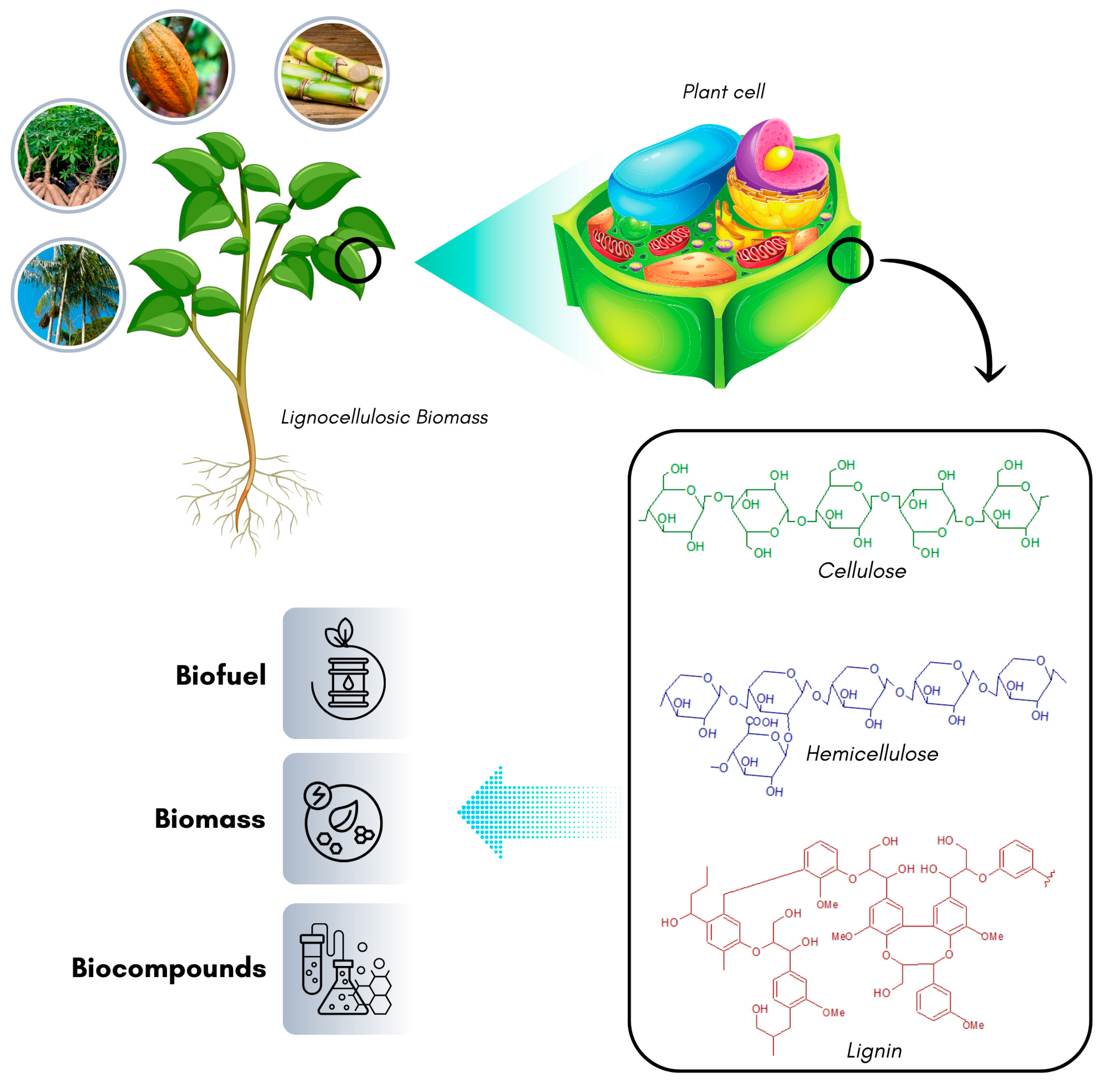
3.2. Lignocellulose Waste
Types of Lignocellulosic Waste and Applications
3.3. Structural Components of Lignocellulosic Residues
3.3.1. Cellulose
3.3.2. Hemicellulose
3.3.3. Lignin
3.4. Pretreatment of Lignocellulosic Biomass
3.4.1. Physical Pretreatments
3.4.2. Chemical Pretreatments
3.4.3. Biological Pretreatments
3.5. Enrichment of Algal and Microalgal Biomass with Lignocellulosic Waste to Produce Biofuels
3.5.1. Biodiesel
3.5.2. Bioethanol
3.5.3. Biobutanol
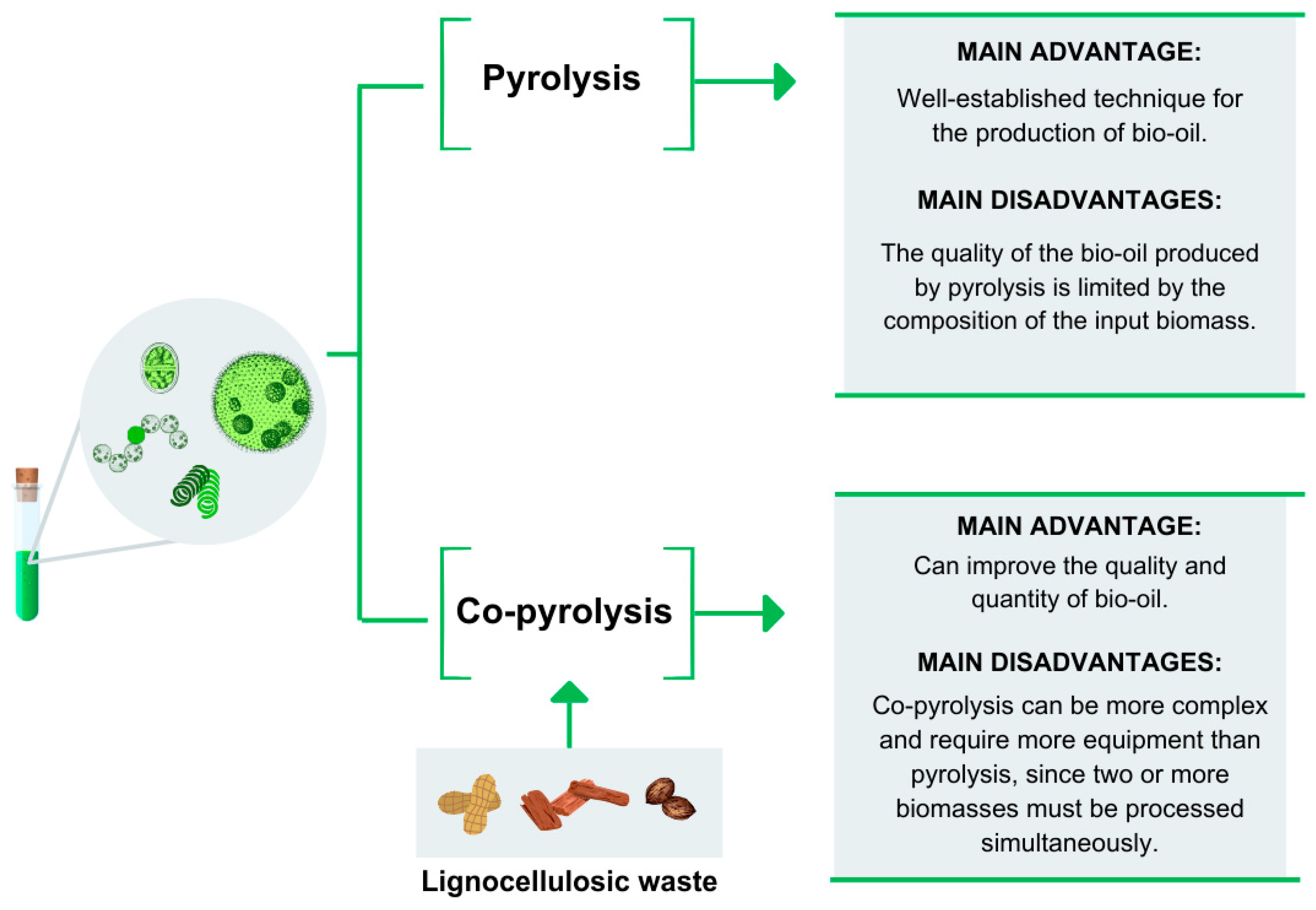
3.5.4. Bio-Oil
3.6. Biogas
3.6.1. Biohydrogen
3.6.2. Biomethane
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Condor, B.E.; De Luna, M.D.G.; Chang, Y.H.; Chen, J.H.; Leong, Y.K.; Chen, P.T.; Chen, C.Y.; Lee, D.J.; Chang, J.S. Bioethanol Production from Microalgae Biomass at High-Solids Loadings. Bioresour. Technol. 2022, 363, 128002. [Google Scholar]
- Zafar, S.U.; Mehra, A.; Nesamma, A.A.; Jutur, P.P. Innovations in Algal Biorefineries for Production of Sustainable Value Chain Biochemicals from the Photosynthetic Cell Factories. Algal. Res. Biomass Biofuels Bioprod. 2023, 69, 102949. [Google Scholar]
- Kumar, V.S.; Sarkar, S.D.; Das, B.K.; Sarkar, D.J.; Gogoi, P.; Maurye, P.; Mitra, T.; Talukder, A.K.; Ganguly, S.; Nag, S.K. Sustainable Biodiesel Production from Microalgae Graesiella Emersonii through Valorization of Garden Wastes-Based Vermicompost. Sci. Total Environ. 2022, 807, 150995. [Google Scholar] [CrossRef] [PubMed]
- Möllers, K.B.; Cannella, D.; Jørgensen, H.; Frigaard, N.U. Cyanobacterial Biomass as Carbohydrate and Nutrient Feedstock for Bioethanol Production by Yeast Fermentation. Biotechnol. Biofuels 2014, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.A.; Sharma, M.; Sharma, M.; Sharma, G.D.; Passari, A.K.; Bhasin, S. Valorization of Agro-Industrial Residues for Production of Commercial Biorefinery Products. Fuel 2022, 322, 124284. [Google Scholar] [CrossRef]
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, P.S.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Khan, T.Y.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae Biomass as a Sustainable Source for Biofuel, Biochemical and Biobased Value-Added Products: An Integrated Biorefinery Concept. Fuel 2022, 307, 121782. [Google Scholar] [CrossRef]
- Ho, S.H.; Huang, S.W.; Chen, C.Y.; Hasunuma, T.; Kondo, A.; Chang, J.S. Bioethanol Production Using Carbohydrate-Rich Microalgae Biomass as Feedstock. Biorefineries Technol. 2013, 135, 191. [Google Scholar] [CrossRef] [PubMed]
- LRizza, L.S.; Smachetti, M.E.S.; Do Nascimento, M.; Salerno, G.L.; Curatti, L. Bioprospecting for Native Microalgae as an Alternative Source of Sugars for the Production of Bioethanol. Algal Res. Biomass Biofuels Bioprod. 2017, 22, 140. [Google Scholar]
- Kim, S.M.; Kim, J.Y.; Park, J.; Bae, E.H.; Kang, J.S.; Kim, K.Y.; Choi, Y.E. Volatile Fatty Acid-Treated Mixotrophic Cultivation of Lipid/Carbohydrate-Rich Cyanobacterial Species, Pseudanabaena Mucicola GO0704, for the Enhancement of Biofuel Production. Bioresour. Technol. 2023, 367, 128066. [Google Scholar] [CrossRef]
- Chong, J.W.R.; Yew, G.Y.; Khoo, K.S.; Ho, S.H.; Show, P.L. Recent Advances on Food Waste Pretreatment Technology via Microalgae for Source of Polyhydroxyalkanoates. J. Environ. Manag. 2021, 293, 112782. [Google Scholar] [CrossRef]
- Vyas, S.; Patel, A.; Risse, E.N.; Krikigianni, E.; Rova, U.; Christakopoulos, P.; Matsakas, L. Biosynthesis of microalgal lipids, proteins, lutein, and carbohydrates using fish farming wastewater and forest biomass under photoautotrophic and heterotrophic cultivation. Bioresour. Technol. 2022, 359, 127494. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Banerjee, C.; Chang, J.S.; Shukla, P. Valorization of Wastewater through Microalgae as a Prospect for Generation of Biofuel and High-Value Products. J. Clean. Prod. 2022, 362, 132114. [Google Scholar] [CrossRef]
- Karnaouri, A.; Chalima, A.; Kalogiannis, K.G.; Varamogianni-Mamatsi, D.; Lappas, A.; Topakas, E. Utilization of Lignocellulosic Biomass towards the Production of Omega-3 Fatty Acids by the Heterotrophic Marine Microalga Crypthecodinium cohnii. Bioresour. Technol. 2020, 303, 122899. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wu, Y.H.; Wang, J.H.; Wang, X.X.; Deantes-Espinosa, V.M.; Dao, G.H.; Tong, X.; Hu, H.Y. Heterotrophic Cultivation of Microalgae in Straw Lignocellulose Hydrolysate for Production of High-Value Biomass Rich in Polyunsaturated Fatty Acids (PUFA). Chem. Eng. J. 2019, 367, 37. [Google Scholar] [CrossRef]
- Mourão, M.M.; Xavier, L.P.; Urbatzka, R.; Figueiroa, L.B.; Costa, C.E.F.D.; Dias, C.G.B.T.; Schneider, M.P.C.; Vasconcelos, V.; Santos, A.V. Characterization and Biotechnological Potential of Intracellular Polyhydroxybutyrate by Stigeoclonium sp. B23 Using Cassava Peel as Carbon Source. Polymers 2021, 13, 687. [Google Scholar] [CrossRef] [PubMed]
- Manzoor, M.; Ahmad, Q.U.A.; Aslam, A.; Jabeen, F.; Rasul, A.; Schenk, P.M.; Qazi, J.I. Mixotrophic Cultivation of Scenedesmus dimorphus in Sugarcane Bagasse Hydrolysate. Environ. Prog. Sustain. Energy 2020, 39, e13334. [Google Scholar] [CrossRef]
- Almutairi, A.W.; Al-Hasawi, Z.M.; Abomohra, A.E.F. Valorization of Lipidic Food Waste for Enhanced Biodiesel Recovery through Two-Step Conversion: A Novel Microalgae-Integrated Approach. Bioresour. Technol. 2021, 342, 125966. [Google Scholar] [CrossRef] [PubMed]
- Asimakopoulou, G.; Karnaouri, A.; Staikos, S.; Stefanidis, S.D.; Kalogiannis, K.G.; Lappas, A.A.; Topakas, E. Production of Omega-3 Fatty Acids from the Microalga Crypthecodinium Cohnii by Utilizing Both Pentose and Hexose Sugars from Agricultural Residues. Fermentation 2021, 7, 219. [Google Scholar] [CrossRef]
- Norzagaray-Valenzuela, C.D.; Valdez-Ortiz, A.; Shelton, L.M.; Jiménez-Edeza, M.; Rivera-López, J.; Valdez-Flores, M.A.; Germán-Báez, L.J. Residual Biomasses and Protein Hydrolysates of Three Green Microalgae Species Exhibit Antioxidant and Anti-Aging Activity. J. Appl. Phycol. 2017, 29, 189–198. [Google Scholar] [CrossRef]
- Rempel, A.; de Souza Sossella, F.; Margarites, A.C.; Astolfi, A.L.; Steinmetz, R.L.R.; Kunz, A.; Treichel, H.; Colla, L.M. Colla, Bioethanol from Spirulina Platensis Biomass and the Use of Residuals to Produce Biomethane: An Energy Efficient Approach. Bioresour. Technol. 2019, 288, 121588. [Google Scholar] [CrossRef]
- Tsai, T.Y.; Lo, Y.C.; Dong, C.D.; Nagarajan, D.; Chang, J.S.; Lee, D.J. Biobutanol Production from Lignocellulosic Biomass Using Immobilized Clostridium acetobutylicum. Appl. Energy 2020, 277, 115531. [Google Scholar] [CrossRef]
- Chen, J.H.; Liu, L.; Lim, P.E.; Wei, D. Effects of Sugarcane Bagasse Hydrolysate (SCBH) on Cell Growth and Fatty Acid Accumulation of Heterotrophic Chlorella protothecoides. Bioprocess Biosyst. Eng. 2019, 42, 1129. [Google Scholar]
- Spennati, E.; Casazza, A.A.; Converti, A.; Padula, M.P.; Dehghani, F.; Perego, P.; Valtchev, P. Winery Waste Valorisation as Microalgae Culture Medium: A Step Forward for Food Circular Economy. Sep. Purif. Technol. 2022, 293, 121088. [Google Scholar] [CrossRef]
- Miazek, K.; Remacle, C.; Richel, A.; Goffin, D. Beech Wood Fagus Sylvatica Dilute-Acid Hydrolysate as a Feedstock to Support Chlorella sorokiniana Biomass, Fatty Acid and Pigment Production. Bioresour. Technol. 2017, 230, 122. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Gengiah, K.; Rajendran, N.; Al-Ghanim, K.A.; Govindarajan, M.; Gurunathan, B. Process and Technoeconomic Analysis of Bioethanol Production from Residual Biomass of Marine Macroalgae Ulva lactuca. Sci. Total Environ. 2023, 868, 161661. [Google Scholar] [CrossRef]
- Freitas, B.C.; Morais, M.G.; Costa, J.A. Chlorella minutissima Grown with Xylose and Arabinose in Tubular Photobioreactors: Evaluation of Kinetics, Carbohydrate Production, and Protein Profile. Can. J. Chem. Eng. 2022, 100, S49. [Google Scholar] [CrossRef]
- Nagappan, S.; Devendran, S.; Tsai, P.C.; Dahms, H.U.; Ponnusamy, V.K. Ponnusamy, Potential of Two-Stage Cultivation in Microalgae Biofuel Production. Fuel 2019, 252, 339. [Google Scholar] [CrossRef]
- Tarhan, S.Z.; Koçer, A.T.; Özçimen, D.; Gökalp, İ. Cultivation of Green Microalgae by Recovering Aqueous Nutrients in Hydrothermal Carbonization Process Water of Biomass Wastes. J. Water Process Eng. 2021, 40, 101783. [Google Scholar] [CrossRef]
- Ramos-Martinez, E.M.; Fimognari, L.; Rasmussen, M.K.; Sakuragi, Y. Secretion of Acetylxylan Esterase From Chlamydomonas reinhardtii Enables Utilization of Lignocellulosic Biomass as a Carbon Source. Front. Bioeng. Biotechnol. 2019, 7, 35. [Google Scholar] [CrossRef]
- Wensel, P.C.; Bule, M.; Gao, A.; Pelaez-Samaniego, M.R.; Yu, L.; Hiscox, W.; Helms, G.L.; Davis, W.C.; Kirchhoff, H.; Garcia-Perez, M.; et al. Biorefinery Processing of Waste to Supply Cost-Effective and Sustainable Inputs for Two-Stage Microalgal Cultivation. Appl. Sci. 2022, 12, 1485. [Google Scholar] [CrossRef]
- Saral, J.S.; Ranganathan, P. A Hydrothermal Co-Liquefaction of Spirulina Platensis with Rice Husk, Coconut Shell and HDPE for Biocrude Production. Bioresour. Technol. 2022, 363, 127911. [Google Scholar] [CrossRef]
- Yew, G.Y.; Khoo, K.S.; Chia, W.Y.; Ho, Y.C.; Law, C.L.; Leong, H.Y.; Show, P.L. A Novel Lipids Recovery Strategy for Biofuels Generation on Microalgae Chlorella Cultivation with Waste Molasses. J. Water Process Eng. 2020, 38, 101665. [Google Scholar] [CrossRef]
- Liang, Y. Producing Liquid Transportation Fuels from Heterotrophic Microalgae. Appl. Energy 2013, 104, 860. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Su, C.H.; Yu, Y.K.; Huong, D.T.M. Huong, Sugarcane Bagasse as a Novel Carbon Source for Heterotrophic Cultivation of Oleaginous Microalga Schizochytrium sp. Ind. Crops Prod. 2018, 121, 99. [Google Scholar] [CrossRef]
- Manzoor, M.; Jabeen, F.; Ahmad, Q.U.A.; Younis, T.; Eltanahy, E.; Schenk, P.M. Sugarcane Bagasse Hydrolysate as Organic Carbon Substrate for Mixotrophic Cultivation of Nannochloropsissp Br2. Waste Biomass Valorization 2021, 12, 2321. [Google Scholar] [CrossRef]
- Singh, A.; Bajar, S.; Devi, A.; Bishnoi, N.R. Adding Value to Agro-Industrial Waste for Cellulase and Xylanase Production via Solid-State Bioconversion. Biomass Convers. Biorefinery 2021, 13, 7481–7490. [Google Scholar] [CrossRef]
- Ampese, L.C.; Buller, L.S.; Myers, J.; Timko, M.T.; Martins, G.; Forster-Carneiro, T. Valorization of Macaúba Husks from Biodiesel Production Using Subcritical Water Hydrolysis Pretreatment Followed by Anaerobic Digestion. J. Environ. Chem. Engineering 2021, 9, 105656. [Google Scholar] [CrossRef]
- Patsalou, M.; Chrysargyris, A.; Tzortzakis, N.; Koutinas, M. A Biorefinery for Conversion of Citrus Peel Waste into Essential Oils, Pectin, Fertilizer and Succinic Acid via Different Fermentation Strategies. Waste Manag. 2020, 113, 469. [Google Scholar] [CrossRef]
- Beigbeder, J.B.; Boboescu, I.Z.; Damay, J.; Duret, X.; Bhatti, S.; Lavoie, J.M. Phytoremediation of Bark-Hydrolysate Fermentation Effluents and Bioaccumulation of Added-Value Molecules by Designed Microalgal Consortia. Algal Res. Biomass Biofuels Bioprod. 2019, 42, 101585. [Google Scholar] [CrossRef]
- Bolonhesi, I.B.D.T.M.; Andreani, C.L.; De Melo, M.R.; Gomes, S.D.; Lopes, D.D. Biomass Immobilization in Hydrolyzed Lignocellulosic Material Can Enhance Biohydrogen Production from Cassava Residues. Biochem. Eng. J. 2023, 190, 108725. [Google Scholar] [CrossRef]
- Li, W.; Shen, Y.; Liu, H.; Huang, X.; Xu, B.; Zhong, C.; Jia, S. Bioconversion of Lignocellulosic Biomass into Bacterial Nanocellulose: Challenges and Perspectives. Green Chem. Eng. 2022, 4, 160–172. [Google Scholar] [CrossRef]
- De Medeiros, T.D.M.; Dufossé, L.; Bicas, J.L. Lignocellulosic Substrates as Starting Materials for the Production of Bioactive Biopigments. Food Chem. X 2022, 13, 100223. [Google Scholar] [CrossRef] [PubMed]
- Khomlaem, C.; Aloui, H.; Singhvi, M.; Kim, B.S. Production of Polyhydroxyalkanoates and Astaxanthin from Lignocellulosic Biomass in High Cell Density Membrane Bioreactor. Chem. Eng. J. 2023, 451, 138641. [Google Scholar] [CrossRef]
- Lu, H.; Yadav, V.; Bilal, M.; Iqbal, H.M. Iqbal, Bioprospecting Microbial Hosts to Valorize Lignocellulose Biomass—Environmental Perspectives and Value-Added Bioproducts. Chemosphere 2022, 288, 132574. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tahmasebi, A.; Yu, J.; Xu, J.; Huang, F.; Mamaeva, A. A Comparative Study of Microwave-Induced Pyrolysis of Lignocellulosic and Algal Biomass. Bioresour. Technol. 2015, 190, 89. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, A.; Sharma, R.K. Conversion of Lignocellulosic Biomass: Production of Bioethanol and Bioelectricity Using Wheat Straw Hydrolysate in Electrochemical Bioreactor. Heliyon 2023, 9, e12951. [Google Scholar] [CrossRef]
- Dos Santos, L.C.; Adarme, O.F.H.; Baêta, B.E.L.; Gurgel, L.V.A.; De Aquino, S.F. Production of Biogas (Methane and Hydrogen) from Anaerobic Digestion of Hemicellulosic Hydrolysate Generated in the Oxidative Pretreatment of Coffee Husks. Bioresour. Technol. 2018, 263, 601. [Google Scholar] [CrossRef] [PubMed]
- Buratto, R.T.; Cocero, M.J.; Martín, Á. Characterization of Industrial Açaí Pulp Residues and Valorization by Microwave-Assisted Extraction. Chem. Eng. Process. Process Intensif. 2021, 160, 108269. [Google Scholar] [CrossRef]
- Targhi, N.K.; Tavakoli, O.; Nazemi, A.H. Co-Pyrolysis of Lentil Husk Wastes and Chlorella vulgaris: Bio-Oil and Biochar Yields Optimization. J. Anal. Appl. Pyrolysis 2022, 165, 105548. [Google Scholar] [CrossRef]
- Gomes, M.A.; dos Santos Rocha, M.S.R.; Barbosa, K.L.; de Abreu, Í.B.S.; de Oliveira Pimentel, W.R.; De Farias Silva, C.E.; Almeida, R.M.R.G.; de Magalhaes Cabral Albuquerque, E.C.; Vieira, R.C. Agricultural Coconut Cultivation Wastes as Feedstock for Lignocellulosic Ethanol Production by Kluyveromyces marxianus. Waste Biomass Valorization 2021, 12, 4943. [Google Scholar] [CrossRef]
- Maceiras, R.; Alfonsín, V.; Seguí, L.; González, J.F. Microwave Assisted Alkaline Pretreatment of Algae Waste in the Production of Cellulosic Bioethanol. Energies 2021, 14, 5981. [Google Scholar] [CrossRef]
- Rezaei, M.; Amiri, H.; Shafiei, M. Aqueous Pretreatment of Triticale Straw for Integrated Production of Hemicellulosic Methane and Cellulosic Butanol. Renew. Energy 2021, 171, 971. [Google Scholar] [CrossRef]
- Tsolcha, O.N.; Patrinou, V.; Economou, C.N.; Dourou, M.; Aggelis, G.; Tekerlekopoulou, A.G. Utilization of Biomass Derived from Cyanobacteria-Based Agro-Industrial Wastewater Treatment and Raisin Residue Extract for Bioethanol Production. Water 2021, 13, 486. [Google Scholar] [CrossRef]
- Liu, L.; Chen, J.; Lim, P.E.; Wei, D. Enhanced Single Cell Oil Production by Mixed Culture of Chlorella pyrenoidosa and Rhodotorula glutinis Using Cassava Bagasse Hydrolysate as Carbon Source. Bioresour. Technol. 2018, 255, 140. [Google Scholar] [CrossRef]
- Olatundun, E.A.; Borokini, O.O.; Betiku, E. Cocoa Pod Husk-Plantain Peel Blend as a Novel Green Heterogeneous Catalyst for Renewable and Sustainable Honne Oil Biodiesel Synthesis: A Case of Biowastes-to-Wealth. Renew. Energy 2020, 166, 163. [Google Scholar] [CrossRef]
- Vignesh, N.S.; Vimali, E.; Sangeetha, R.; Arumugam, M.; Ashokkumar, B.; Ganeshmoorthy, I.; Varalakshmi, P. Sustainable Biofuel from Microalgae: Application of Lignocellulosic Wastes and Bio-Iron Nanoparticle for Biodiesel Production. Fuel 2020, 278, 118326. [Google Scholar] [CrossRef]
- Mu, J.; Li, S.; Chen, D.; Xu, H.; Han, F.; Feng, B.; Li, Y. Enhanced Biomass and Oil Production from Sugarcane Bagasse Hydrolysate (SBH) by Heterotrophic Oleaginous Microalga Chlorella protothecoides. Bioresour. Technol. 2015, 185, 99. [Google Scholar] [CrossRef]
- Maity, S.; Mallick, N. Bioprospecting Marine Microalgae and Cyanobacteria as Alternative Feedstocks for Bioethanol Production. Sustain. Chem. Pharm. 2022, 29, 100798. [Google Scholar] [CrossRef]
- Muharja, M.; Darmayanti, R.F.; Fachri, B.A.; Palupi, B.; Rahmawati, I.; Rizkiana, M.F.; Amini, H.W.; Putri, D.K.Y.; Setiawan, F.A.; Asrofi, M.; et al. Biobutanol Production from Cocoa Pod Husk through a Sequential Green Method: Depectination, Delignification, Enzymatic Hydrolysis, and Extractive Fermentation. Bioresour. Technol. Rep. 2023, 21, 101298. [Google Scholar] [CrossRef]
- López-Linares, J.C.; García-Cubero, M.T.; Lucas, S.; Coca, M. Integral Valorization of Cellulosic and Hemicellulosic Sugars for Biobutanol Production: ABE Fermentation of the Whole Slurry from Microwave Pretreated Brewer’s Spent Grain. Biomass Bioenergy 2020, 135, 105524. [Google Scholar]
- Rasam, S.; Azizi, K.; Moraveji, M.K.; Akbari, A.; Soria-Verdugo, A. Insights into the Co-Pyrolysis of Olive Stone, Waste Polyvinyl Chloride and Spirulina Microalgae Blends through Thermogravimetric Analysis. Algal Res. Biomass Biofuels Bioprod. 2022, 62, 102635. [Google Scholar] [CrossRef]
- Vuppaladadiyam, A.K.; Antunes, E.; Sanchez, P.B.; Duan, H.; Zhao, M. Influence of Microalgae on Synergism during Co-Pyrolysis with Organic Waste Biomass: A Thermogravimetric and Kinetic Analysis. Renew. Energy 2021, 167, 42. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lineage | Compound | Bioproduct | Reference |
|---|---|---|---|
| Chlorella pyrenoidosa | PUFAs | Food supplement: Omega-3 | [14] |
| Phaeodactylum tricornutum | Fucoxanthin/PUFAs | Nutraceuticals: Anti-obesity, anti-diabetic, and anti-cancer/omega-3 | [9] |
| Stigeoclonium sp. B23 | Neutral lipids | Bioplastics: Polyhydroxybutyrate (PHB) | [15] |
| Nannochloropsis sp. BR2 | Carotenoid | Natural antioxidant: Betacarotene | [16] |
| Scenedesmus obliquus SAG276-10 | Fatty acids | Biofuel: Biodiesel | [17] |
| Topics | Databases |
|---|---|
| Lignocellulosic waste AND valorization AND hydrolysate AND microalgae | WOS, SCOPUS AND PUBMED |
| Microalgae AND lignocellulosic waste | |
| Lignocellulosic waste AND biomass AND hydrolysate AND microalgae | |
| Microalgae AND biomass AND hydrolysate | |
| Microalgae AND lignocellulosic waste AND biorefinery | |
| Microalgae OR algal AND biorefinery AND biomass | |
| Lignocellulosic waste AND valorization AND hydrolyzed | |
| Waste biomass AND lignocellulosic AND hydrolyzed AND algae | |
| Microalgae AND lignocellulosic biomass AND biofuel | |
| Lignocellulosic AND cyanobacteria | |
| Lignocellulosic waste AND microalgae AND carbon source | |
| Carbon source AND lignocellulosic waste AND microalgae | |
| Biofuel AND residual biomass AND hydrolyzed | |
| Biomass valorization AND cyanobacteria | |
| Biomass valorization AND waste AND biorefinery | |
| Biomass waste AND Amazonic AND valorization | |
| Feedstock valorization AND agro-industrial wastes AND lignocellulosic waste biomass | |
| Hemicellulosic AND biofuel AND hydrolysate | |
| Hemicellulosic AND biofuel AND hydrolysate AND microalgae | |
| Lignocellulosic AND biohydrogen AND microalgae | |
| Lignocellulosic AND hydrolyzed AND carbon source | |
| Lignocellulosic waste AND recovery AND hydrolysate | |
| Lignocellulosic waste AND Amazonic AND valorization | |
| Lignocellulosic waste AND hydrolysate AND carbon source | |
| Lignocellulosic waste AND hydrolysate | |
| Lignocellulosic waste AND microalgae/cyanobacteria AND hydrolysate | |
| Microalgae AND biomass AND waste | |
| Microalgae AND cocoa waste AND biofuel | |
| Microalgae AND corn waste AND biofuels | |
| Microalgae AND hydrolyzed AND corn waste | |
| Microalgae AND hydrolyzed AND waste | |
| Microalgae AND lignocellulosic biomass AND biofuel | |
| Microalgae AND lignocellulosic waste AND hydrolysate AND carbon source | |
| Photosynthetic microorganism AND lignocellulosic waste AND hydrolysate AND carbon source | |
| Pyrolysis AND microalgae AND bio-oil | |
| Waste AND husk AND reuse | |
| Microalgae AND waste AND hydrolysate | |
| Lignocellulosic waste OR husk AND valorization | |
| Microalgae AND biomass AND waste | |
| Lignocellulosic waste AND valorization AND hydrolysate AND microalgae | |
| Microalgae AND lignocellulosic waste | |
| Lignocellulosic waste AND biomass AND hydrolysate AND microalgae |
| Substrate | Cellulose (%) | Hemicellulose (%) | Lignin (%) | Reference |
|---|---|---|---|---|
| Coffee husk | 32.5 | 20.8 | 27.1 | [48] |
| Açaí seed | 8.5 | 48.1 | 16.4 | [49] |
| Sugarcane bagasse | 45.7 | 23.4 | 18.6 | [35] |
| Citrus peel | 22.4 | 8 | 0.6 | [39] |
| Lentil shell | 27.7 | 8 | 7.1 | [50] |
| Coconut shell | 23.3 | 12.2 | 40 | [51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, D.T.d.; Mescouto, V.A.d.; Paiva, R.d.J.; Silva, S.R.F.d.; Santos, L.A.B.; Serra, G.M.; Xavier, L.P.; Noronha, R.C.R.; Nascimento, L.A.S.d. Use of Residual Lignocellulosic Biomass and Algal Biomass to Produce Biofuels. Int. J. Mol. Sci. 2024, 25, 8299. https://doi.org/10.3390/ijms25158299
Oliveira DTd, Mescouto VAd, Paiva RdJ, Silva SRFd, Santos LAB, Serra GM, Xavier LP, Noronha RCR, Nascimento LASd. Use of Residual Lignocellulosic Biomass and Algal Biomass to Produce Biofuels. International Journal of Molecular Sciences. 2024; 25(15):8299. https://doi.org/10.3390/ijms25158299
Chicago/Turabian StyleOliveira, Deborah Terra de, Vanessa Albuquerque de Mescouto, Rutiléia de Jesus Paiva, Sara Roberta Ferreira da Silva, Luiz Augusto Barbosa Santos, Gustavo Marques Serra, Luciana Pereira Xavier, Renata Coelho Rodrigues Noronha, and Luís Adriano Santos do Nascimento. 2024. "Use of Residual Lignocellulosic Biomass and Algal Biomass to Produce Biofuels" International Journal of Molecular Sciences 25, no. 15: 8299. https://doi.org/10.3390/ijms25158299
APA StyleOliveira, D. T. d., Mescouto, V. A. d., Paiva, R. d. J., Silva, S. R. F. d., Santos, L. A. B., Serra, G. M., Xavier, L. P., Noronha, R. C. R., & Nascimento, L. A. S. d. (2024). Use of Residual Lignocellulosic Biomass and Algal Biomass to Produce Biofuels. International Journal of Molecular Sciences, 25(15), 8299. https://doi.org/10.3390/ijms25158299