

CDC20 Holds Novel Regulation Mechanism in RPA1 during Different Stages of DNA Damage to Induce Radio-Chemoresistance
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
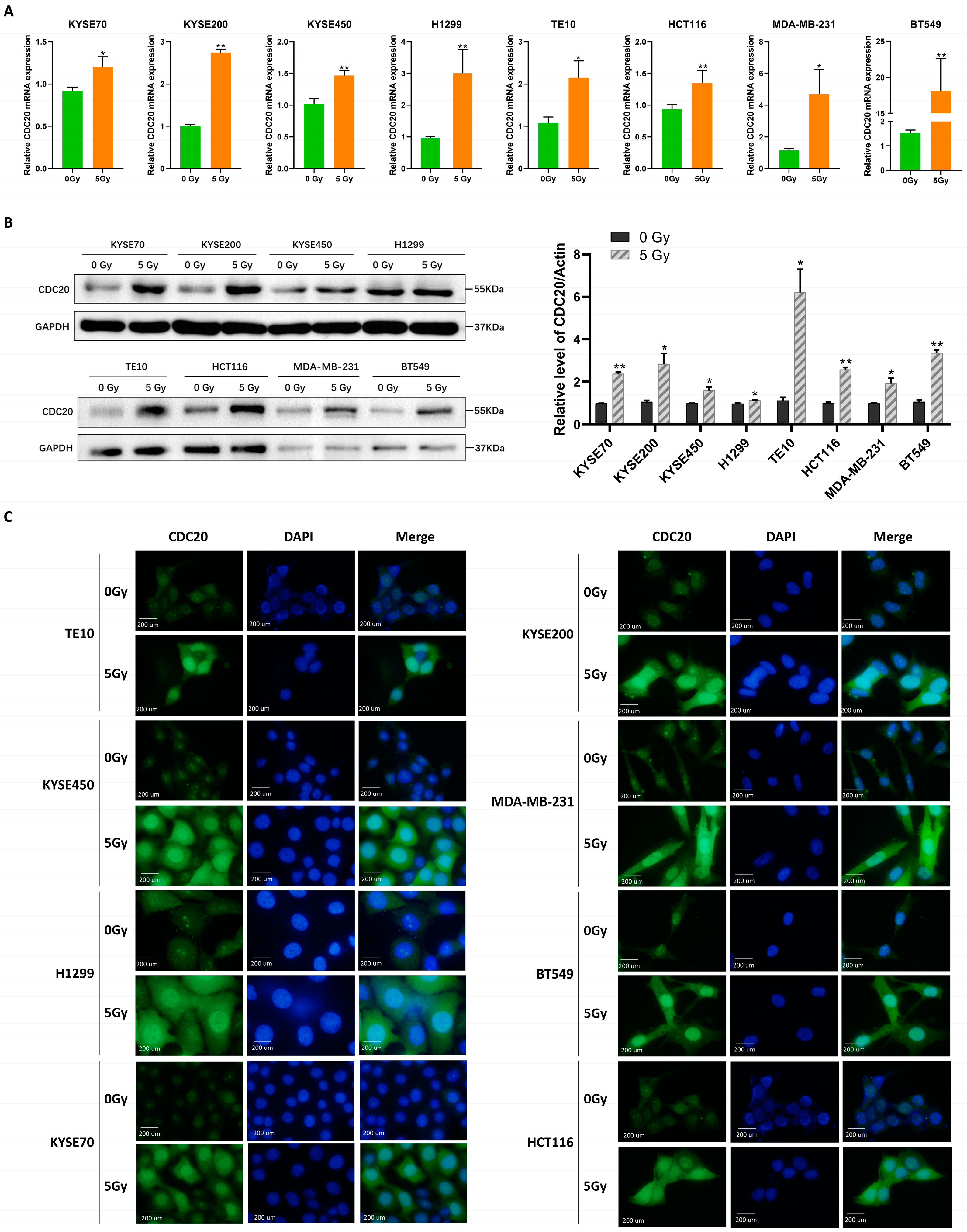
2.1. Radiation Induces Nuclear Accumulation of CDC20 and Affects DNA Damage Response
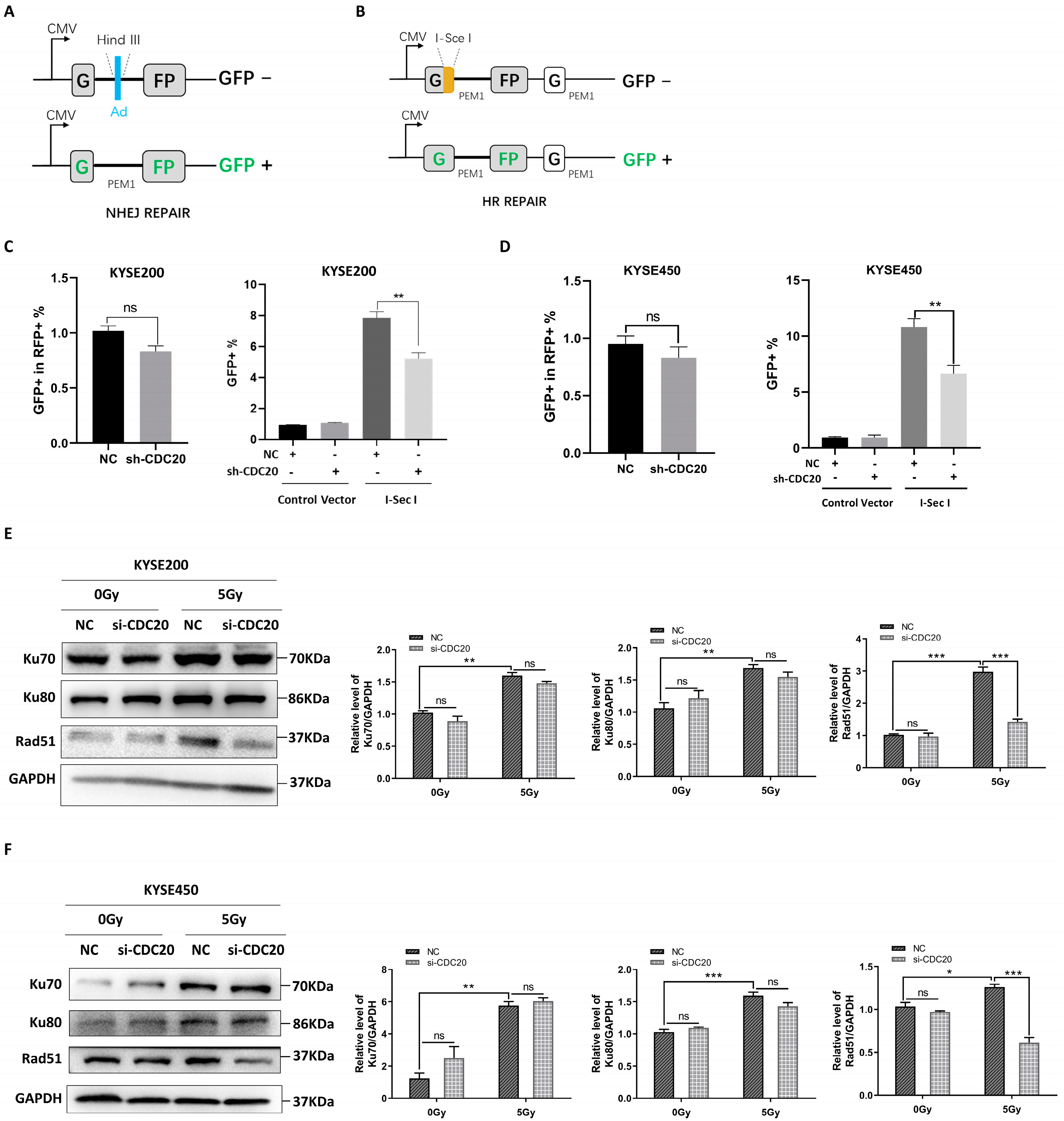
2.2. CDC20 Regulates the Homologous Repair of DSBs
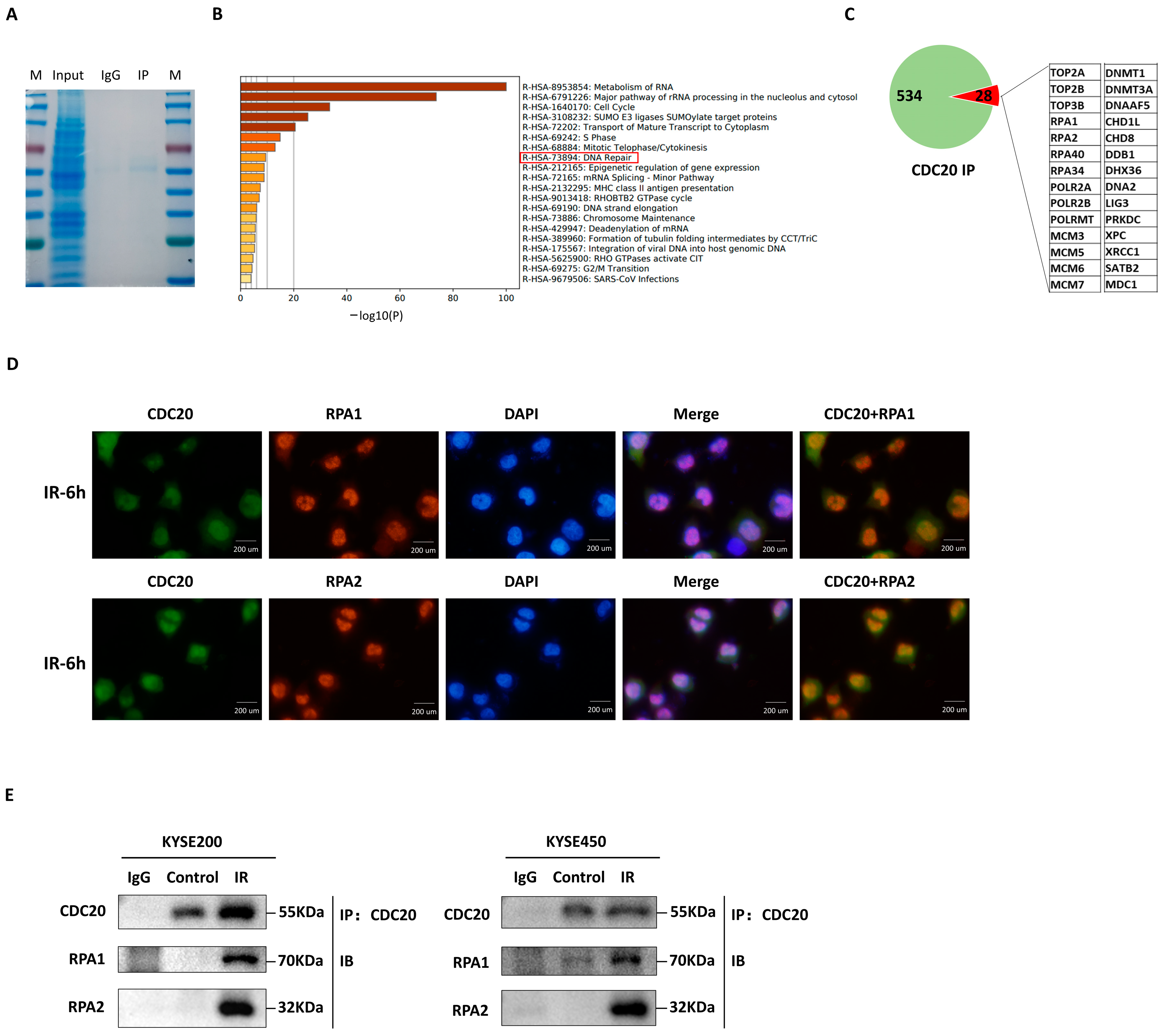
2.3. CDC20 Interacts with RPA1/RPA2 to Assist DNA Damage Repair
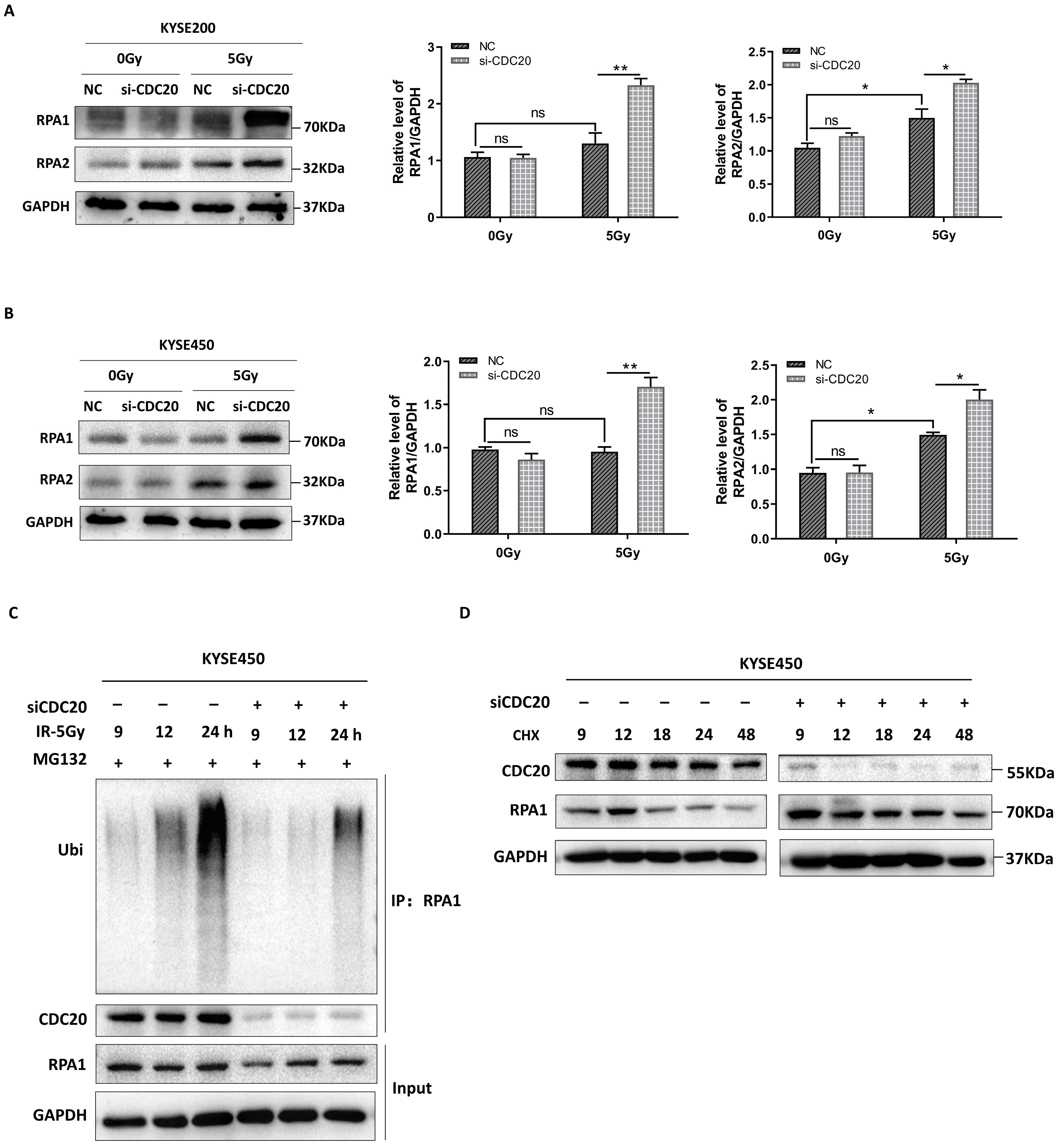
2.4. CDC20 Mediates the Timely Unloading of RPA1 during DNA Damage Repair
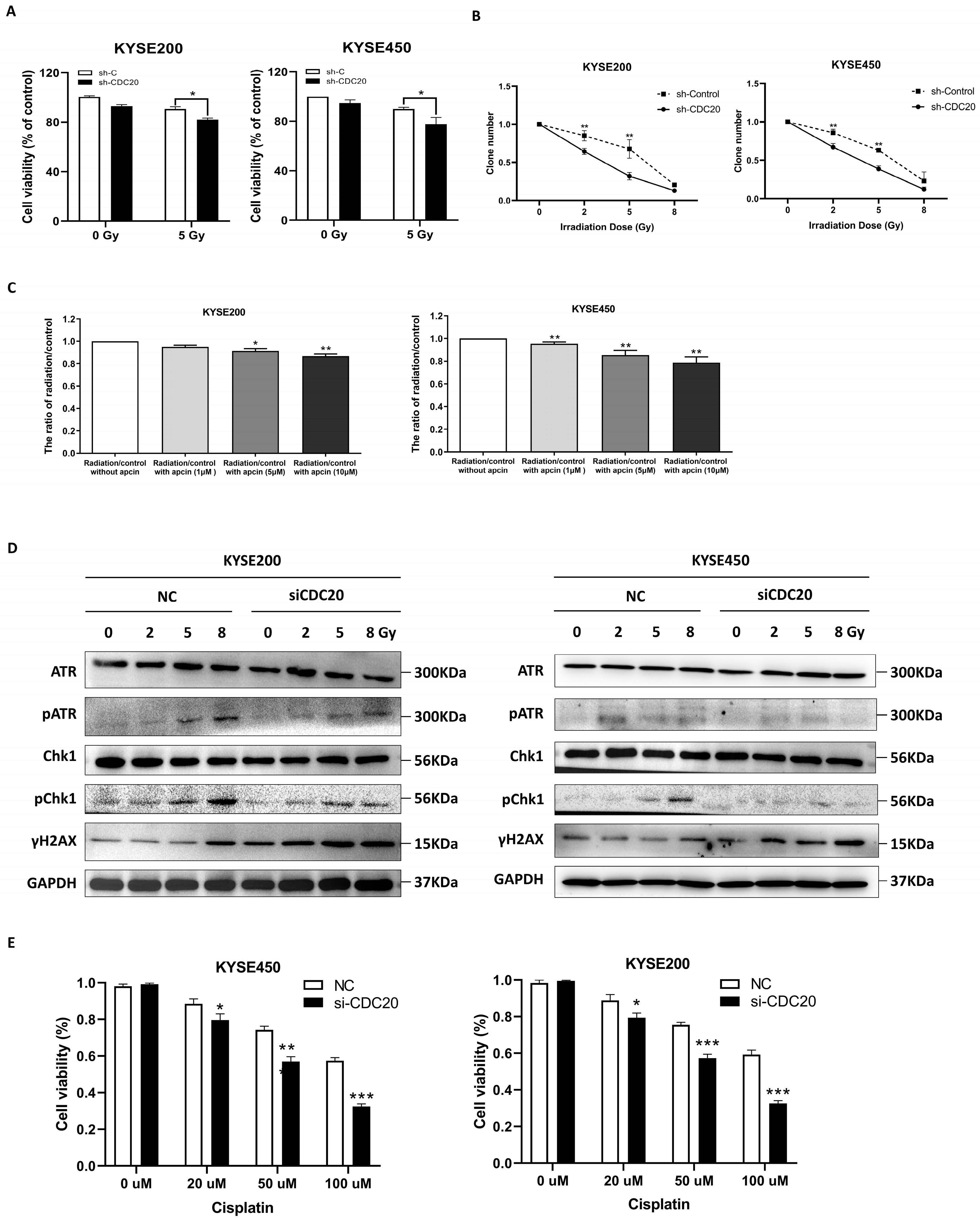
2.5. CDC20 Regulates Radio- and Chemosensitivity in Cancer Cells
2.6. Apcin Regulates Radiosensitivity In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Radiation
4.2. qRT-PCR Analysis
4.3. Western Blot
4.4. Immunofluorescence Staining
4.5. Transfection of siRNA and shRNA Sequence
4.6. Coimmunoprecipitation (Co-IP)
4.7. LC-MS/MS Assay
4.8. Cell Viability and Colony Formation Assay
4.9. Xenograft Model
4.10. IHC Analysis
4.11. TUNEL Assay
4.12. H&E Staining
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nickoloff, J.A.; Sharma, N.; Taylor, L. Clustered DNA Double-Strand Breaks: Biological Effects and Relevance to Cancer Radiotherapy. Genes 2020, 11, 99. [Google Scholar] [CrossRef]
- Huang, R.-X.; Zhou, P.-K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Larsen, B.D.; Benada, J.; Yung, P.Y.K.; Bell, R.A.V.; Pappas, G.; Urban, V.; Ahlskog, J.K.; Kuo, T.T.; Janscak, P.; Megeney, L.A.; et al. Cancer cells use self-inflicted DNA breaks to evade growth limits imposed by genotoxic stress. Science 2022, 376, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Han, Z.; Luo, Q.; Wang, Y.; Li, Q.; Zhou, L.; Zuo, H. Radiotherapy modulates tumor cell fate decisions: A review. Radiat. Oncol. 2022, 17, 196. [Google Scholar] [CrossRef]
- Novo, N.; Romero-Tamayo, S.; Marcuello, C.; Boneta, S.; Blasco-Machin, I.; Velázquez-Campoy, A.; Villanueva, R.; Moreno-Loshuertos, R.; Lostao, A.; Medina, M.; et al. Beyond a platform protein for the degradosome assembly: The Apopto-sis-Inducing Factor as an efficient nuclease involved in chromatinolysis. PNAS Nexus. 2022, 2, pgac312. [Google Scholar] [CrossRef] [PubMed]
- Siklos, M.; Kubicek, S. Therapeutic targeting of chromatin: Status and opportunities. FEBS J. 2021, 289, 1276–1301. [Google Scholar] [CrossRef]
- Qiu, Y.; Hu, W.; Wen, M.; Zhao, W.; Xie, J.; Zhang, J.; Wang, M.; Li, H.; Zhao, Y.; Fu, S.; et al. Low Expression of ECT2 Confers Radiation Therapy Resistance Through Transcription Coupled Nucleolar DNA Damage Repair. Int. J. Radiat. Oncol. 2022, 112, 1229–1242. [Google Scholar] [CrossRef]
- Zhou, W.; Yao, Y.; Scott, A.J.; Wilder-Romans, K.; Dresser, J.J.; Werner, C.K.; Sun, H.; Pratt, D.; Sajjakulnukit, P.; Zhao, S.G.; et al. Purine metabolism regulates DNA repair and therapy resistance in glioblastoma. Nat. Commun. 2020, 11, 3811. [Google Scholar] [CrossRef]
- Jeong, S.M.; Bui, Q.T.; Kwak, M.; Lee, J.Y.; Lee, P.C.-W. Targeting Cdc20 for cancer therapy. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2022, 1877, 188824. [Google Scholar] [CrossRef]
- Gao, Y.; Wen, P.; Chen, B.; Hu, G.; Wu, L.; Xu, A.; Zhao, G. Downregulation of CDC20 Increases Radiosensitivity through Mcl-1/p-Chk1-Mediated DNA Damage and Apoptosis in Tumor Cells. Int. J. Mol. Sci. 2020, 21, 6692. [Google Scholar] [CrossRef] [PubMed]
- Velegzhaninov, I.O.; Belykh, E.S.; Rasova, E.E.; Pylina, Y.I.; Shadrin, D.M.; Klokov, D.Y. Radioresistance, DNA Damage and DNA Repair in Cells with Moderate Overexpression of RPA1. Front. Genet. 2020, 11, 855. [Google Scholar] [CrossRef]
- Valente, D.; Gentileschi, M.P.; Guerrisi, A.; Bruzzaniti, V.; Morrone, A.; Soddu, S.; Verdina, A. Factors to Consider for the Correct Use of γH2AX in the Evaluation of DNA Double-Strand Breaks Damage Caused by Ionizing Radiation. Cancers 2022, 14, 6204. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Y.; Lu, X.; Ding, H.; Han, B.; Song, X.; Miao, H.; Cui, X.; Wei, S.; Liu, W.; et al. CDC20 regulates the cell proliferation and radiosensitivity of P53 mutant HCC cells through the Bcl-2/Bax pathway. Int. J. Biol. Sci. 2021, 17, 3608–3621. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, C.; Chu, M.; Wang, Z.-W.; Xue, B. Cdc20 induces the radioresistance of bladder cancer cells by targeting FoxO1 degradation. Cancer Lett. 2021, 500, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, E.; Mladenova, V.; Stuschke, M.; Iliakis, G. New Facets of DNA Double Strand Break Repair: Radiation Dose as Key Determinant of HR versus c-NHEJ Engagement. Int. J. Mol. Sci. 2023, 24, 14956. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, J. JmjC domain-containing protein 8 (JMJD8) represses Ku70/Ku80 expression via attenuating AKT/NF-κB/COX-2 signaling. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2019, 1866, 118541. [Google Scholar] [CrossRef]
- Tian, Y.; Liu, G.; Wang, H.; Tian, Z.; Cai, Z.; Zhang, F.; Luo, Y.; Wang, S.; Guo, G.; Wang, X.; et al. Valproic acid sensitizes breast cancer cells to hydroxyurea through inhibiting RPA2 hyperphosphorylation-mediated DNA repair pathway. DNA Repair 2017, 58, 1–12. [Google Scholar] [CrossRef]
- Zhang, Z.; Huo, H.; Liao, K.; Wang, Z.; Gong, Z.; Li, Y.; Liu, C.; Hu, G. RPA1 downregulation enhances nasopharyngeal cancer radiosensitivity via blocking RAD51 to the DNA damage site. Exp. Cell Res. 2018, 371, 330–341. [Google Scholar] [CrossRef]
- Caldwell, C.C.; Spies, M. Dynamic elements of replication protein A at the crossroads of DNA replication, recombination, and repair. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 482–507. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Srinivasan, K.; Abdisalaam, S.; Su, F.; Raj, P.; Dozmorov, I.; Mishra, R.; Wakeland, E.K.; Ghose, S.; Mukherjee, S.; et al. RAD51 interconnects between DNA replication, DNA repair and immunity. Nucleic Acids Res. 2017, 45, 4590–4605. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Oleinick, N.L.; Zhang, J. ATR/CHK1 inhibitors and cancer therapy. Radiother. Oncol. 2018, 126, 450–464. [Google Scholar] [CrossRef]
- Baxter, J.S.; Zatreanu, D.; Pettitt, S.J.; Lord, C.J. Resistance to DNA repair inhibitors in cancer. Mol. Oncol. 2022, 16, 3811–3827. [Google Scholar] [CrossRef] [PubMed]
- Washington, C.R.; Moore, K.N. Resistance to Poly (ADP-Ribose) Polymerase Inhibitors (PARPi): Mechanisms and Potential to Reverse. Curr. Oncol. Rep. 2022, 24, 1685–1693. [Google Scholar] [CrossRef]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.-E.; Malki, M.I. DNA Damage/Repair Management in Cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef] [PubMed]
- Ui, A.; Chiba, N.; Yasui, A. Relationship among DNA double-strand break (DSB), DSB repair, and transcription prevents genome instability and cancer. Cancer Sci. 2020, 111, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Xu, F.; Gao, Y.; Hu, G.; Zhu, K.; Lu, H.; Xu, A.; Chen, S.; Wu, L.; Zhao, G. DNA damage-induced translocation of mitochondrial factor HIGD1A into the nucleus regulates homologous recombination and radio/chemo-sensitivity. Oncogene 2022, 41, 1918–1930. [Google Scholar] [CrossRef] [PubMed]
- Saredi, G.; Rouse, J. Ways to unwind with HROB, a new player in homologous recombination. Genes Dev. 2019, 33, 1293–1294. [Google Scholar] [CrossRef] [PubMed]
- Skucha, A.; Ebner, J.; Grebien, F. Roles of SETD2 in Leukemia—Transcription, DNA-Damage, and Beyond. Int. J. Mol. Sci. 2019, 20, 1029. [Google Scholar] [CrossRef]
- Cappadocia, L.; Kochańczyk, T.; Lima, C.D. DNA asymmetry promotes SUMO modification of the single-stranded DNA-binding protein RPA. EMBO J. 2021, 40, e103787. [Google Scholar] [CrossRef]
- Barbour, A.T.; Wuttke, D.S. RPA-like single-stranded DNA-binding protein complexes including CST serve as specialized processivity factors for polymerases. Curr. Opin. Struct. Biol. 2023, 81, 102611. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, N.; Origanti, S.; Davenport, E.P.; Gandhi, D.; Kaniecki, K.; Mehl, R.A.; Greene, E.C.; Dockendorff, C.; Antony, E. Monitoring Replication Protein A (RPA) dynamics in homologous recombination through site-specific incorporation of non-canonical amino acids. Nucleic Acids Res. 2017, 45, 9413–9426. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, H.J.; Lee, J.S.; Jeoung, D.; Kang, C.M.; Bae, S.; Lee, S.J.; Kwon, S.H.; Kang, D.; Lee, Y.S. A novel function for HSF1-induced mitotic exit failure and genomic instability through direct interaction between HSF1 and Cdc20. Oncogene 2008, 27, 2999–3009. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-X.; Pan, Y.-Y.; You, C.-G. CDK1, CCNB1, CDC20, BUB1, MAD2L1, MCM3, BUB1B, MCM2, and RFC4 May Be Potential Therapeutic Targets for Hepatocellular Carcinoma Using Integrated Bioinformatic Analysis. BioMed Res. Int. 2019, 2019, 1245072. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Brill, S.J. Rfc4 Interacts with Rpa1 and Is Required for both DNA replication and DNA damage checkpoints in Saccharomyces cerevisiae. Mol. Cell. Biol. 2001, 21, 3725–3737. [Google Scholar] [CrossRef] [PubMed]
- Bartek, J.; Hodny, Z. SUMO boosts the DNA damage response barrier against cancer. Cancer Cell 2010, 17, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Li, X.; Shen, J.; Zhao, Y.; Zhong, S.; Lai, L.; Niu, H.; Qi, Z. ssDNA accessibility of Rad51 is regulated by orchestrating multiple RPA dynamics. Nat. Commun. 2023, 14, 3864. [Google Scholar] [CrossRef] [PubMed]
- Di, Z.; Sanyuan, S.; Hong, L.; Dahai, Y. Enhanced radiosensitivity and G2/M arrest were observed in radioresistant esophageal cancer cells by knocking down RPA expression. Cell Biochem. Biophys. 2014, 70, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Clemente-Blanco, A. Cell Cycle and DNA Repair Regulation in the Damage Response: Protein Phosphatases Take Over the Reins. Int. J. Mol. Sci. 2020, 21, 446. [Google Scholar] [CrossRef]
- Yam, C.Q.X.; Lim, H.H.; Surana, U. DNA damage checkpoint execution and the rules of its disengagement. Front. Cell Dev. Biol. 2022, 10, 1020643. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, L.; Wang, Z.; Wang, J.; Roychoudhury, S.; Tomasik, B.; Wu, G.; Wang, G.; Rao, X.; Zhou, R. Replication Stress: A Review of Novel Targets to Enhance Radiosensitivity-From Bench to Clinic. Front. Oncol. 2022, 12, 838637. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Ge, Y.; Zang, Y.; Xu, M.; Jin, L.; Wang, Y.; Xu, X.; Xue, B.; Wang, Z.; Wang, L. CDC20 promotes radioresistance of prostate cancer by activating Twist1 expression. Apoptosis 2023, 28, 1584–1595. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.D.; Cleary, R.T.; Gujar, A.; Mahlokozera, T.; Kim, A.H. CDC20 regulates sensitivity to chemotherapy and radiation in glioblastoma stem cells. PLoS ONE 2022, 17, e0270251. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Meng, J. CDC20: A novel therapeutic target in cancer. Am. J. Transl. Res. 2023, 15, 678–693. [Google Scholar] [PubMed]
- Volonte, D.; Sedorovitz, M.; Galbiati, F. Impaired Cdc20 signaling promotes senescence in normal cells and apoptosis in non-small cell lung cancer cells. J. Biol. Chem. 2022, 298, 102405. [Google Scholar] [CrossRef]
- Deng, D.-X.; Li, C.-Y.; Zheng, Z.-Y.; Wen, B.; Liao, L.-D.; Zhang, X.-J.; Li, E.-M.; Xu, L.-Y. Prenylated PALM2 Promotes the Migration of Esophageal Squamous Cell Carcinoma Cells Through Activating Ezrin. Mol. Cell. Proteom. 2023, 22, 100593. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; He, H.; Pillai, S.; Xiong, Y.; Challa, S.; Xu, L.; Chellappan, S.; Yang, S. GATA3 transcription factor abrogates Smad4 transcription factor-mediated fascin overexpression, invadopodium formation, and breast cancer cell invasion. J. Biol. Chem. 2013, 288, 36971–36982. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Song, M.; Wang, P.; Zhao, R.; Chen, H.; Zhang, M.; Shi, Y.; Liu, K.; Liu, F.; Yang, R.; et al. Targeted therapy of the AKT kinase inhibits esophageal squamous cell carcinoma growth in vitro and in vivo. Int. J. Cancer 2019, 145, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Lai, C.; He, Q.-M.; Cai, Y.; Yu, H.; Zhong, W.; Chen, S.; Wu, F.-C.; Chen, H. Integrated proteomics and phosphoproteomics analyses of esophageal cancer cells with different invasive abilities. Life Sci. 2023, 332, 122078. [Google Scholar] [CrossRef]
- Sun, Y.; Dai, H.; Chen, S.; Zhang, Y.; Wu, T.; Cao, X.; Zhao, G.; Xu, A.; Wang, J.; Wu, L. Disruption of Chromosomal Architecture of cox2 Locus Sensitizes Lung Cancer Cells to Radiotherapy. Mol. Ther. 2018, 26, 2456–2465. [Google Scholar] [CrossRef]
- Wang, R.; Shang, Y.; Chen, B.; Xu, F.; Zhang, J.; Zhang, Z.; Zhao, X.; Wan, X.; Xu, A.; Wu, L.; et al. Protein disulfide isomerase blocks the interaction of LC3II-PHB2 and promotes mTOR signaling to regulate autophagy and radio/chemo-sensitivity. Cell Death Dis. 2022, 13, 851. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, B.; Wang, R.; Xu, A.; Wu, L.; Lu, H.; Zhao, G. Knockdown of RRM1 in tumor cells promotes radio-/chemotherapy induced ferroptosis by regulating p53 ubiquitination and p21-GPX4 signaling axis. Cell Death Discov. 2022, 8, 343. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; Wen, P.; Shao, C.; Ye, C.; Chen, Y.; You, J.; Su, Z. CDC20 Holds Novel Regulation Mechanism in RPA1 during Different Stages of DNA Damage to Induce Radio-Chemoresistance. Int. J. Mol. Sci. 2024, 25, 8383. https://doi.org/10.3390/ijms25158383
Gao Y, Wen P, Shao C, Ye C, Chen Y, You J, Su Z. CDC20 Holds Novel Regulation Mechanism in RPA1 during Different Stages of DNA Damage to Induce Radio-Chemoresistance. International Journal of Molecular Sciences. 2024; 25(15):8383. https://doi.org/10.3390/ijms25158383
Chicago/Turabian StyleGao, Yang, Pengbo Wen, Chenran Shao, Cheng Ye, Yuji Chen, Junyu You, and Zhongjing Su. 2024. "CDC20 Holds Novel Regulation Mechanism in RPA1 during Different Stages of DNA Damage to Induce Radio-Chemoresistance" International Journal of Molecular Sciences 25, no. 15: 8383. https://doi.org/10.3390/ijms25158383