


Senescence in Adipose-Derived Stem Cells: Biological Mechanisms and Therapeutic Challenges

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
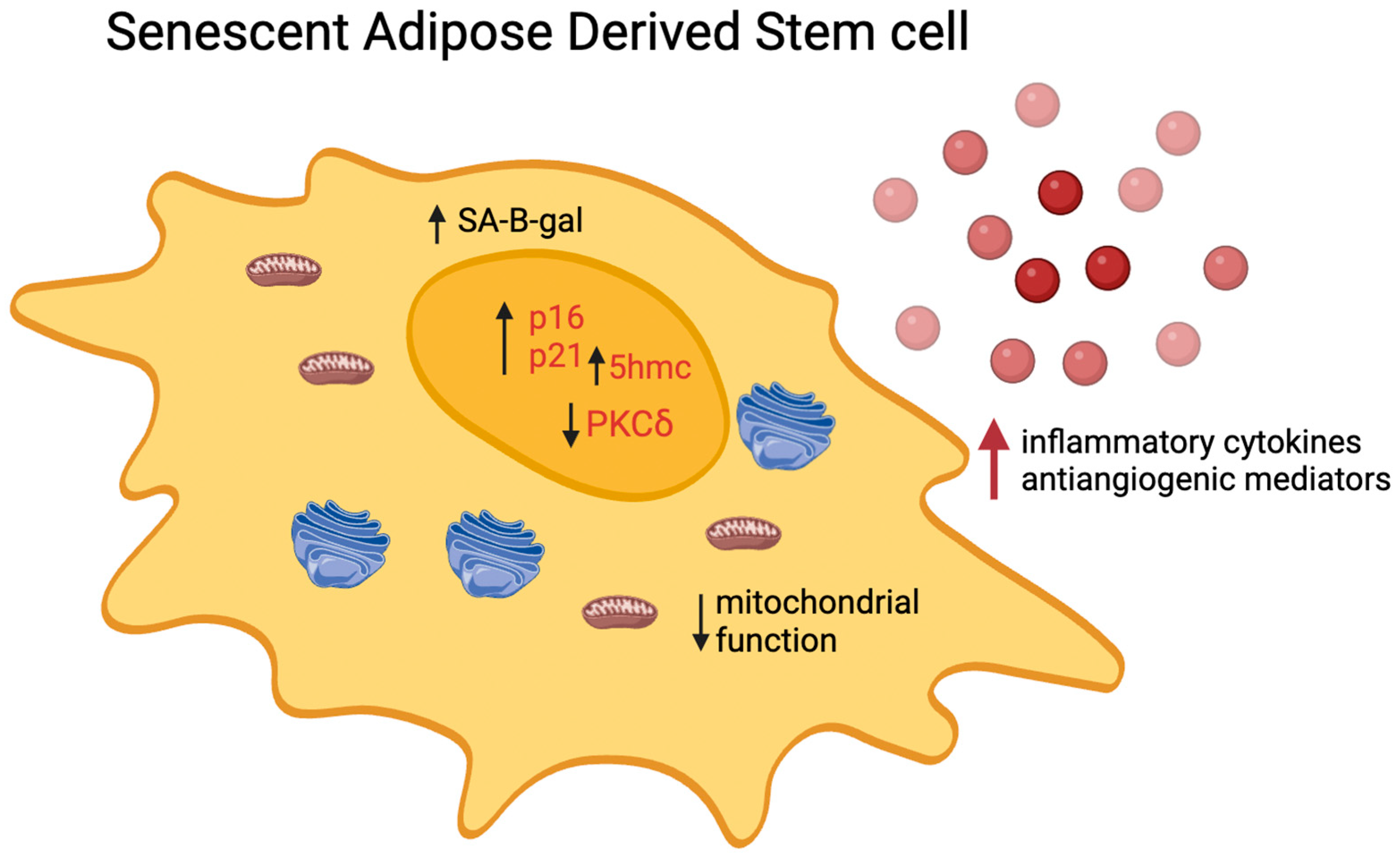
2. Cellular Senescence Phenotypes and Molecular Mechanisms of Senescence
2.1. Senescence-Associated β-galactosidase (SA-β-gal) Activity
2.2. Markers of Persistent DNA Damage
2.3. p21WAF1/CIP
2.4. Absence of Detectable Cellular Proliferation Activity
3. Main Causes of Accelerated Senescence
4. Senescence-Induced Functionality Changes in ADSCs
5. Senolytic Agents and Their Applications
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gimble, J.M.; Bunnell, B.A.; Floyd, Z.E. Prospecting for Adipose Progenitor Cell Biomarkers: Biopanning for Gold with In Vivo Phage Display. Cell Stem Cell 2011, 9, 1–2. [Google Scholar] [CrossRef]
- Storti, G.; Scioli, M.G.; Kim, B.-S.; Terriaca, S.; Fiorelli, E.; Orlandi, A.; Cervelli, V. Mesenchymal Stem Cells in Adipose Tissue and Extracellular Vesicles in Ovarian Cancer Patients: A Bridge toward Metastatic Diffusion or a New Therapeutic Opportunity? Cells Transplant. 2021, 10, 2117. [Google Scholar] [CrossRef]
- Cervelli, V.; Storti, G. Novel Strategies to Improve Graft Survival and Retention; Springer International Publishing: Berlin/Heidelberg, Germany, 2022; ISBN 978-303077455-4/978-303077454-7. [Google Scholar]
- Liu, M.; Lei, H.; Dong, P.; Fu, X.; Yang, Z.; Yang, Y.; Ma, J.; Liu, X.; Cao, Y.; Xiao, R. Adipose-Derived Mesenchymal Stem Cells from the Elderly Exhibit Decreased Migration and Differentiation Abilities with Senescent Properties. Cell Transplant. 2017, 26, 1505–1519. [Google Scholar] [CrossRef]
- Storti, G.; Scioli, M.G.; Kim, B.-S.; Orlandi, A.; Cervelli, V. Adipose-Derived Stem Cells in Bone Tissue Engineering: Useful Tools with New Applications. Stem Cells Int. 2019, 2019, 3673857. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Alicka, M.; Major, P.; Wysocki, M.; Marycz, K. Adipose-Derived Mesenchymal Stem Cells Isolated from Patients with Type 2 Diabetes Show Reduced “Stemness” through an Altered Secretome Profile, Impaired Anti-Oxidative Protection, and Mitochondrial Dynamics Deterioration. J. Clin. Med. 2019, 8, 765. [Google Scholar] [CrossRef]
- Scioli, M.G.; Storti, G.; Bielli, A.; Sanchez, M.; Scimeca, M.; Gimble, J.M.; Cervelli, V.; Orlandi, A. CD146 Expression Regulates Osteochondrogenic Differentiation of Human Adipose-Derived Stem Cells. J. Cell. Physiol. 2022, 237, 589–602. [Google Scholar] [CrossRef]
- Scioli, M.G.; Storti, G.; D’amico, F.; Gentile, P.; Kim, B.S.; Cervelli, V.; Orlandi, A. Adipose-Derived Stem Cells in Cancer Progression: New Perspectives and Opportunities. Int. J. Mol. Sci. 2019, 20, 3296. [Google Scholar] [CrossRef]
- Hofmann, E.; Soppert, J.; Ruhl, T.; Gousopoulos, E.; Gerra, S.; Storti, G.; Tian, Y.; Brandhofer, M.; Schweizer, R.; Song, S.-Y.; et al. The Role of Macrophage Migration Inhibitory Factor in Adipose-Derived Stem Cells Under Hypoxia. Front. Physiol. 2021, 12, 638448. [Google Scholar] [CrossRef]
- Kern, S.; Eichler, H.; Stoeve, J.; Klüter, H.; Bieback, K. Comparative Analysis of Mesenchymal Stem Cells from Bone Marrow, Umbilical Cord Blood, or Adipose Tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef]
- Zhu, Y.; Liu, T.; Song, K.; Fan, X.; Ma, X.; Cui, Z. Adipose-Derived Stem Cell: A Better Stem Cell than BMSC. Cell Biochem. Funct. 2008, 26, 664–675. [Google Scholar] [CrossRef]
- Ruhl, T.; Storti, G.; Pallua, N. Proliferation, Metabolic Activity, and Adipogenic Differentiation of Human Preadipocytes Exposed to 2 Surfactants In Vitro. J. Pharm. Sci. 2018, 107, 1408–1415. [Google Scholar] [CrossRef]
- Storti, G.; Favi, E.; Albanesi, F.; Kim, B.-S.; Cervelli, V. Adipose-Derived Stem/Stromal Cells in Kidney Transplantation: Status Quo and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 11188. [Google Scholar] [CrossRef]
- De Araújo, R.S.; Mussalem, M.G.V.B.; Carrijo, G.S.; Bani, J.V.d.F.; Ferreira, L.M. Adipose Tissue Derivatives in Peripheral Nerve Regeneration after Transection: A Systematic Review. Bioengineering. 2024, 11, 697. [Google Scholar] [CrossRef]
- Tang, H.; He, Y.; Liang, Z.; Li, J.; Dong, Z.; Liao, Y. The Therapeutic Effect of Adipose-Derived Stem Cells on Soft Tissue Injury after Radiotherapy and Their Value for Breast Reconstruction. Stem Cell Res. Ther. 2022, 13, 493. [Google Scholar] [CrossRef]
- Surowiecka, A.; Chrapusta, A.; Klimeczek-Chrapusta, M.; Korzeniowski, T.; Drukała, J.; Strużyna, J. Mesenchymal Stem Cells in Burn Wound Management. Int. J. Mol. Sci. 2022, 23, 15339. [Google Scholar] [CrossRef]
- Zayed, M.A.; Sultan, S.; Alsaab, H.O.; Yousof, S.M.; Alrefaei, G.I.; Alsubhi, N.H.; Alkarim, S.; Al Ghamdi, K.S.; Bagabir, S.A.; Jana, A.; et al. Stem-Cell-Based Therapy: The Celestial Weapon against Neurological Disorders. Cells 2022, 11, 3478. [Google Scholar] [CrossRef]
- Raghuram, A.C.; Yu, R.P.; Lo, A.Y.; Sung, C.J.; Bircan, M.; Thompson, H.J.; Wong, A.K. Role of Stem Cell Therapies in Treating Chronic Wounds: A Systematic Review. World J. Stem Cells 2020, 12, 659–675. [Google Scholar] [CrossRef]
- Gao, J.G.; Yu, M.S.; Zhang, M.M.; Gu, X.W.; Ren, Y.; Zhou, X.X.; Chen, D.; Yan, T.L.; Li, Y.M.; Jin, X. Adipose-Derived Mesenchymal Stem Cells Alleviate TNBS-Induced Colitis in Rats by Influencing Intestinal Epithelial Cell Regeneration, Wnt Signaling, and T Cell Immunity. World J. Gastroenterol. 2020, 26, 3750–3766. [Google Scholar] [CrossRef]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and Limits of Adipose-Derived Stem Cells (ADSCs) and Mesenchymal Stem Cells (MSCs) in Wound Healing. Int. J. Mol. Sci. 2020, 21, 1306. [Google Scholar] [CrossRef]
- Alió del Barrio, J.L.; De la Mata, A.; De Miguel, M.P.; Arnalich-Montiel, F.; Nieto-Miguel, T.; El Zarif, M.; Cadenas-Martín, M.; López-Paniagua, M.; Galindo, S.; Calonge, M.; et al. Corneal Regeneration Using Adipose-Derived Mesenchymal Stem Cells. Cells 2022, 11, 2549. [Google Scholar] [CrossRef]
- Issa, M.R.; Naja, A.S.; Bouji, N.Z.; Sagherian, B.H. The Role of Adipose-Derived Mesenchymal Stem Cells in Knee Osteoarthritis: A Meta-Analysis of Randomized Controlled Trials. Ther. Adv. Musculoskelet. Dis. 2022, 14, 1–13. [Google Scholar] [CrossRef]
- Huang, T.; Tong, H.; Zhou, H.; Wang, J.; Hu, L.; Wang, Y.; Huang, Z. ADSC-Exosomes Alleviate MTX-Induced Rat Neuronal Damage by Activating Nrf2-ARE Pathway. J. Mol. Neurosci. 2022, 72, 1334–1344. [Google Scholar] [CrossRef]
- Neves Vialle, E.; Fracaro, L.; Barchiki, F.; Correa Dominguez, A.; De Oliveira Arruda, A.; Olandoski, M.; Roberto, P.; Brofman, S.; Lúcia, C.; Rebelatto, K. Human Adipose-Derived Stem Cells Reduce Cellular Damage after Experimental Spinal Cord Injury in Rats. Biomedicines. 2023, 11, 1394. [Google Scholar] [CrossRef]
- Hu, X.; Pan, J.; Li, Y.; Jiang, Y.; Zheng, H.; Shi, R.; Zhang, Q.; Liu, C.; Tian, H.; Zhang, Z.; et al. Extracellular Vesicles from Adipose-Derived Stem Cells Promote Microglia M2 Polarization and Neurological Recovery in a Mouse Model of Transient Middle Cerebral Artery Occlusion. Stem Cell Res. Ther. 2022, 13, 21. [Google Scholar] [CrossRef]
- Grun, L.K.; Maurmann, R.M.; Scholl, J.N.; Fogaça, M.E.; Schmitz, C.R.R.; Dias, C.K.; Gasparotto, J.; Padoin, A.V.; Mottin, C.C.; Klamt, F.; et al. Obesity Drives Adipose-Derived Stem Cells into a Senescent and Dysfunctional Phenotype Associated with P38MAPK/NF-KB Axis. Immun. Ageing 2023, 20, 51. [Google Scholar] [CrossRef]
- Mantovani, C.M.; Terenghi, G.; Magnaghi, V. Senescence in Adipose-Derived Stem Cells and Its Implications in Nerve Regeneration. Neural Regen. Res. 2014, 9, 10–15. [Google Scholar] [CrossRef]
- Weng, Z.; Wang, Y.; Ouchi, T.; Liu, H.; Qiao, X.; Wu, C.; Zhao, Z.; Li, L.; Li, B. Mesenchymal Stem/Stromal Cell Senescence: Hallmarks, Mechanisms, and Combating Strategies. Stem Cells Transl. Med. 2022, 11, 356–371. [Google Scholar] [CrossRef]
- Li, R.; Teng, Y.; Guo, Y.; Ren, J.; Li, R.; Luo, H.; Chen, D.; Feng, Z.; Fu, Z.; Zou, X.; et al. Aging-related decrease of histone methyltransferase SUV39H1 in adipose-derived stem cells enhanced SASP. Mech. Ageing Dev. 2023, 215, 111868. [Google Scholar] [CrossRef]
- Lunyak, V.V.; Amaro-Ortiz, A.; Gaur, M. Mesenchymal Stem Cells Secretory Responses: Senescence Messaging Secretome and Immunomodulation Perspective. Front. Genet. 2017, 8, 220. [Google Scholar] [CrossRef]
- Gaur, M.; Wang, L.; Ortiz, A.; Dobke, M.; Jordan, K.; Lunyak, V. Acute Genotoxic Stress-Induced Senescence in Human Mesenchymal Cells Drives a Unique Composition of Senescence Messaging Secretome (SMS). J. Stem Cell Res. Ther. 2017, 7, 8. [Google Scholar] [CrossRef]
- Gaur, M.; Dobke, M.; Lunyak, V.V. Methods and Strategies for Procurement, Isolation, Characterization, and Assessment of Senescence of Human Mesenchymal Stem Cells from Adipose Tissue. In Stem Cells and Aging: Methods and Protocols; Turksen, K., Ed.; Springer: New York, NY, USA, 2019; pp. 37–92. ISBN 978-1-4939-9713-8. [Google Scholar]
- Hayflick, L.; Moorhead, P.S. The Serial Cultivation of Human Diploid Cell Strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Geesman, G.J.; Hostikka, S.L.; Atallah, M.; Blackwell, B.; Lee, E.; Cook, P.J.; Pasaniuc, B.; Shariat, G.; Halperin, E.; et al. Inhibition of Activated Pericentromeric SINE/Alu Repeat Transcription in Senescent Human Adult Stem Cells Reinstates Self-Renewal. Cell Cycle 2011, 10, 3016–3030. [Google Scholar] [CrossRef] [PubMed]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic Review of Patient Factors Affecting Adipose Stem Cell Viability and Function: Implications for Regenerative Therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.F.; Niu, P.; Wang, L.; Vogelsang, M.; Gaur, M.; Krastins, B.; Zhao, Y.; Smagul, A.; Nussupbekova, A.; Akanov, A.A.; et al. Opposing Activities of Oncogenic MIR17HG and Tumor Suppressive MIR100HG Clusters and Their Gene Targets Regulate Replicative Senescence in Human Adult Stem Cells. NPJ Aging Mech. Dis. 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Shi, G.; Lei, X.; Huang, Y.; Li, X.; Bai, L.; Qin, C. Age-Related Alteration in Characteristics, Function, and Transcription Features of ADSCs. Stem Cell Res. Ther. 2021, 12, 473. [Google Scholar] [CrossRef]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-Associated β-Galactosidase Is Lysosomal β-Galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Xue, L.; Lin, H.; Luo, J. Depletion of NUAK2 blocks the stemness and angiogenesis and facilitates senescence of lung adenocarcinoma cells via enhancing ferroptosis. Cell Div. 2024, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Siennicka, K.; Zołocińska, A.; Dȩbski, T.; Pojda, Z. Comparison of the Donor Age-Dependent and In Vitro Culture-Dependent Mesenchymal Stem Cell Aging in Rat Model. Stem Cells Int. 2021, 1, 6665358. [Google Scholar] [CrossRef]
- Targa, L. Contribution to the Study of Mesenchymal Stromal Stem Cells Heterogeneity, Focus On Surface Markers and Senescence. Doctoral Thesis, Université de Lorraine, Lorraine, France, 2019. [Google Scholar]
- De Mera-Rodríguez, J.A.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Is Senescence-Associated β-Galactosidase a Reliable in Vivo Marker of Cellular Senescence During Embryonic Development? Front. Cell Dev. Biol. 2021, 9, 623175. [Google Scholar] [CrossRef]
- Kohli, J.; Wang, B.; Brandenburg, S.M.; Basisty, N.; Evangelou, K.; Varela-Eirin, M.; Campisi, J.; Schilling, B.; Gorgoulis, V.; Demaria, M. Algorithmic assessment of cellular senescence in experimental and clinical specimens. Nat. Protoc. 2021, 16, 2471–2498. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Liao, C.; Liu, G.; Xu, Y.; Tan, J.; Song, Z. Age-Related Changes in the Regenerative Potential of Adipose-Derived Stem Cells Isolated from the Prominent Fat Pads in Human Lower Eyelids. PLoS ONE 2016, 11, e0166590. [Google Scholar] [CrossRef] [PubMed]
- Zajic, L.B.; Webb, T.L.; Webb, P.; Coy, J.W.; Dow, S.W.; Quimby, J.M. Comparison of Proliferative and Immunomodulatory Potential of Adipose-Derived Mesenchymal Stem Cells from Young and Geriatric Cats. J. Feline Med. Surg. 2017, 19, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Bagge, J.; MacLeod, J.N.; Berg, L.C. Cellular Proliferation of Equine Bone Marrow- and Adipose Tissue-Derived Mesenchymal Stem Cells Decline With Increasing Donor Age. Front. Vet. Sci. 2020, 7, 602403. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Liao, W.; Sheng, P.; Fu, M.; He, A.; Huang, G. Biological Character of Human Adipose-Derived Adult Stem Cells and Influence of Donor Age on Cell Replication in Culture. Sci. China Ser. C-Life Sci. 2007, 50, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor Age Negatively Impacts Adipose Tissue-Derived Mesenchymal Stem Cell Expansion and Differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef]
- Niu, P.; Smagul, A.; Wang, L.; Sadvakas, A.; Sha, Y.; Pérez, L.M.; Nussupbekova, A.; Amirbekov, A.; Akanov, A.A.; Gálvez, B.G.; et al. Transcriptional Profiling of Interleukin-2-Primed Human Adipose Derived Mesenchymal Stem Cells Revealed Dramatic Changes in Stem Cells Response Imposed by Replicative Senescence. Oncotarget 2015, 6, 17938–17957. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.S.; Carter, G.; El Bassit, G.; Patel, A.A.; Cooper, D.R.; Murr, M.; Patel, N.A. Adipose-Derived Stem Cells from Lean and Obese Humans Show Depot Specific Differences in Their Stem Cell Markers, Exosome Contents and Senescence: Role of Protein Kinase C Delta (PKCδ) in Adipose Stem Cell Niche. Stem Cell Investig. 2016, 3, 2016. [Google Scholar] [CrossRef]
- Lopes-Paciencia, S.; Saint-Germain, E.; Rowell, M.C.; Ruiz, A.F.; Kalegari, P.; Ferbeyre, G. The Senescence-Associated Secretory Phenotype and Its Regulation. Cytokine 2019, 117, 15–22. [Google Scholar] [CrossRef]
- Acosta, J.C.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.P.; Athineos, D.; Kang, T.W.; Lasitschka, F.; Andrulis, M.; et al. A Complex Secretory Program Orchestrated by the Inflammasome Controls Paracrine Senescence. Nat. Cell Biol. 2013, 15, 978–990. [Google Scholar] [CrossRef]
- Borkowska, J.; Domaszewska-Szostek, A.; Kołodziej, P.; Wicik, Z.; Połosak, J.; Buyanovskaya, O.; Charzewski, L.; Stańczyk, M.; Noszczyk, B.; Puzianowska-Kuznicka, M. Alterations in 5hmC Level and Genomic Distribution in Aging-Related Epigenetic Drift in Human Adipose Stem Cells. Epigenomics 2020, 12, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; Yu, J.H.; Liao, Y.T.; Liu, K.H.; Chiang, E.R.; Chang, M.C.; Wang, J. pan Comparison of the Infant and Adult AdiposeDerived Mesenchymal Stem Cells in Proliferation, Senescence, Anti-Oxidative Ability and Differentiation Potential. Tissue Eng. Regen. Med. 2022, 19, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Xiong, H.; Chen, J.; Yang, X.; Liu, Y.; Guo, J.; Jiang, T.; Xu, Z.; Yuan, M.; Liu, Y.; et al. The Whole Profiling and Competing Endogenous RNA Network Analyses of Noncoding RNAs in Adipose-Derived Stem Cells from Diabetic, Old, and Young Patients. Stem Cell Res. Ther. 2021, 12, 313. [Google Scholar] [CrossRef] [PubMed]
- Rennert, R.C.; Sorkin, M.; Januszyk, M.; Duscher, D.; Kosaraju, R.; Chung, M.T.; Lennon, J.; Radiya-Dixit, A.; Raghvendra, S.; Maan, Z.N.; et al. Diabetes Impairs the Angiogenic Potential of Adipose-Derived Stem Cells by Selectively Depleting Cellular Subpopulations. Stem Cell Res. Ther. 2014, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Alicka, M.; Kornicka-Garbowska, K.; Kucharczyk, K.; Kȩpska, M.; Rocken, M.; Marycz, K. Age-Dependent Impairment of Adipose-Derived Stem Cells Isolated from Horses. Stem Cell Res. Ther. 2020, 11, 4. [Google Scholar] [CrossRef]
- Ren, S.; Li, C.; Xiong, H.; Wu, Q.; Wu, X.; Xiong, Z.; Dong, L.; Shu, B.; Wei, W.; Ma, C.; et al. The Rejuvenation and Functional Restoration of Aged Adipose Stem Cells by DUXAP10 Knockdown via the Regulation of the MiR-214-3p/RASSF5 Axis. Stem Cells Transl. Med. 2024, 13, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Meng, Y.; Zhu, X.; Saadiq, I.M.; Jordan, K.L.; Eirin, A.; Lerman, L.O. Metabolic Syndrome Increases Senescence Associated Micro-RNAs in Extracellular Vesicles Derived from Swine and Human Mesenchymal Stem/Stromal Cells. Cell Commun. Signal. 2020, 18, 124. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Martin, E.C.; Matossian, M.D.; Brock, C.K.; Nguyen, K.; Collins-Burow, B.; Burow, M.E. The Effect of Obesity on Adipose-Derived Stromal Cells and Adipose Tissue and Their Impact on Cancer. Cancer Metastasis Rev. 2022, 41, 549–573. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, G.; Hu, Y.; Mohsin, A.; Chen, Z.; Hao, W.; Li, Z.; Gao, W.Q.; Guo, M.; Xu, H. Uncovering Impaired Mitochondrial and Lysosomal Function in Adipose-Derived Stem Cells from Obese Individuals with Altered Biological Activity. Stem Cell Res. Ther. 2024, 15, 12. [Google Scholar] [CrossRef]
- Mizunoe, Y.; Kobayashi, M.; Tagawa, R.; Nakagawa, Y.; Shimano, H.; Higami, Y. Association between Lysosomal Dysfunction and Obesity-Related Pathology: A Key Knowledge to Prevent Metabolic Syndrome. Int. J. Mol. Sci. 2019, 20, 3688. [Google Scholar] [CrossRef]
- Smith, J.; Su, X.; El-Maghrabi, R.; Stahl, P.D.; Abumrad, N.A. Opposite Regulation of CD36 Ubiquitination by Fatty Acids and Insulin: Effects on Fatty Acid Uptake. J. Biol. Chem. 2008, 283, 13578–13585. [Google Scholar] [CrossRef]
- Parmley, L.A.; Elkins, N.D.; Fini, M.A.; Liu, Y.E.; Repine, J.E.; Wright, R.M. Alpha-4/Beta-1 and Alpha-L/Beta-2 Integrins Mediate Cytokine Induced Lung Leukocyte-Epithelial Adhesion and Injury. Br. J. Pharmacol. 2007, 152, 915–929. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.W.; He, S.P.; Lan, J.G.; Zhu, W.Z. Honokiol Ameliorates Cisplatin-Induced Acute Kidney Injury via Inhibition of Mitochondrial Fission. Br. J. Pharmacol. 2022, 179, 3886–3904. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Yao, Y.; Hu, H.; Wu, J.; Li, J.; Li, L.; Wu, J.; Sun, M.; Deng, Z.; Zhang, Y.; et al. PDHA1 Hyperacetylation-Mediated Lactate Overproduction Promotes Sepsis-Induced Acute Kidney Injury via Fis1 Lactylation. Cell Death Dis. 2023, 14, 457. [Google Scholar] [CrossRef]
- Eirin, A.; Thaler, R.; Glasstetter, L.M.; Xing, L.; Zhu, X.Y.; Osborne, A.C.; Mondesir, R.; Bhagwate, A.V.; Lerman, A.; van Wijnen, A.J.; et al. Obesity-Driven Mitochondrial Dysfunction in Human Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells Involves Epigenetic Changes. Cell Death Dis. 2024, 15, 387. [Google Scholar] [CrossRef]
- Glasstetter, L.M.; Oderinde, T.S.; Mirchandani, M.; Rajagopalan, K.S.; Barsom, S.H.; Thaler, R.; Siddiqi, S.; Zhu, X.Y.; Tang, H.; Jordan, K.L.; et al. Obesity and Dyslipidemia Are Associated with Partially Reversible Modifications to DNA Hydroxymethylation of Apoptosis- and Senescence-Related Genes in Swine Adipose-Derived Mesenchymal Stem/Stromal Cells. Stem Cell Res. Ther. 2023, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Rajagopalan, K.S.; Kazeminia, S.; Glasstetter, L.M.; Farahani, R.A.; Zhu, X.Y.; Tang, H.; Jordan, K.L.; Chade, A.R.; Lerman, A.; Lerman, L.O.; et al. Metabolic Syndrome Induces Epigenetic Alterations in Mitochondria-Related Genes in Swine Mesenchymal Stem Cells. Cells 2023, 12, 1274. [Google Scholar] [CrossRef] [PubMed]
- Henry, A.G.; Aghamohammadzadeh, S.; Samaroo, H.; Chen, Y.; Mou, K.; Needle, E.; Hirst, W.D. Pathogenic LRRK2 Mutations, through Increased Kinase Activity, Produce Enlarged Lysosomes with Reduced Degradative Capacity and Increase ATP13A2 Expression. Hum. Mol. Genet. 2015, 24, 6013–6028. [Google Scholar] [CrossRef]
- Park, J.T.; Lee, Y.S.; Cho, K.A.; Park, S.C. Adjustment of the Lysosomal-Mitochondrial Axis for Control of Cellular Senescence. Ageing Res. Rev. 2018, 47, 176–182. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, D.; Liu, Z.; Jin, W.; Huang, G.; Wei, Z.; Wang, D.; Deng, C. Diabetes-Induced Glucolipotoxicity Impairs Wound Healing Ability of Adipose-Derived Stem Cells-through the MiR-1248/CITED2/HIF-1α Pathway. Aging 2020, 12, 6947–6965. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Kalodimou, V.E.; Mavrogonatou, E.; Karamanou, K.; Yiacoumettis, A.M.; Panagiotou, P.N.; Pratsinis, H.; Kletsas, D. Decreased Differentiation Capacity and Altered Expression of Extracellular Matrix Components in Irradiation-Mediated Senescent Human Breast Adipose-Derived Stem Cells. IUBMB Life 2022, 74, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Ko, Y.J.; Lee, M.W.; Park, H.J.; Park, Y.J.; Kim, D.I.; Sung, K.W.; Koo, H.H.; Yoo, K.H. Effect of Low Oxygen Tension on the Biological Characteristics of Human Bone Marrow Mesenchymal Stem Cells. Cell Stress Chaperones 2016, 21, 1089–1099. [Google Scholar] [CrossRef] [PubMed]
- Ratushnyy, A.; Lobanova, M.; Buravkova, L.B. Expansion of Adipose Tissue-Derived Stromal Cells at “Physiologic” Hypoxia Attenuates Replicative Senescence. Cell Biochem. Funct. 2017, 35, 232–243. [Google Scholar] [CrossRef]
- Xu, Z.; Lin, L.; Fan, Y.; Huselstein, C.; De Isla, N.; He, X.; Chen, Y.; Li, Y. Secretome of Mesenchymal Stem Cells from Consecutive Hypoxic Cultures Promotes Resolution of Lung Inflammation by Reprogramming Anti-Inflammatory Macrophages. Int. J. Mol. Sci. 2022, 23, 4333. [Google Scholar] [CrossRef] [PubMed]
- Shin, L.; Peterson, D.A. Impaired Therapeutic Capacity of Autologous Stem Cells in a Model of Type 2 Diabetes. Stem Cells Transl. Med. 2012, 1, 125–135. [Google Scholar] [CrossRef]
- El-Badawy, A.; El-Badri, N. Clinical Efficacy of Stem Cell Therapy for Diabetes Mellitus: A Meta-Analysis. PLoS ONE 2016, 11, e0151938. [Google Scholar] [CrossRef]
- Inoue, O.; Usui, S.; Takashima, S.I.; Nomura, A.; Yamaguchi, K.; Takeda, Y.; Goten, C.; Hamaoka, T.; Ootsuji, H.; Murai, H.; et al. Diabetes Impairs the Angiogenic Capacity of Human Adipose-Derived Stem Cells by Reducing the CD271+ Subpopulation in Adipose Tissue. Biochem. Biophys. Res. Commun. 2019, 517, 369–375. [Google Scholar] [CrossRef]
- Bigarella, C.L.; Liang, R.; Ghaffari, S. Stem Cells and the Impact of ROS Signaling. Development 2014, 141, 4206–4218. [Google Scholar] [CrossRef]
- Shin, L.; Peterson, D.A. Human Mesenchymal Stem Cell Grafts Enhance Normal and Impaired Wound Healing by Recruiting Existing Endogenous Tissue Stem/Progenitor Cells. Stem Cells Transl. Med. 2013, 2, 33–42. [Google Scholar] [CrossRef]
- Xiao, S.; Liu, Z.; Yao, Y.; Wei, Z.R.; Wang, D.; Deng, C. Diabetic Human Adipose-Derived Stem Cells Accelerate Pressure Ulcer Healing by Inducing Angiogenesis and Neurogenesis. Stem Cells Dev. 2019, 28, 319–328. [Google Scholar] [CrossRef]
- Scruggs, B.A.; Semon, J.A.; Zhang, X.; Zhang, S.; Bowles, A.C.; Pandey, A.C.; Imhof, K.M.P.; Kalueff, A.V.; Gimble, J.M.; Bunnell, B.A. Age of the Donor Reduces the Ability of Human Adipose-Derived Stem Cells to Alleviate Symptoms in the Experimental Autoimmune Encephalomyelitis Mouse Model. Stem Cells Transl. Med. 2013, 2, 797–807. [Google Scholar] [CrossRef]
- Maredziak, M.; Marycz, K.; Tomaszewski, K.A.; Kornicka, K.; Henry, B.M. The Influence of Aging on the Regenerative Potential of Human Adipose Derived Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 2152435. [Google Scholar] [CrossRef] [PubMed]
- Ratushnyy, A.; Ezdakova, M.; Buravkova, L. Secretome of Senescent Adipose-Derived Mesenchymal Stem Cells Negatively Regulates Angiogenesis. Int. J. Mol. Sci. 2020, 21, 1802. [Google Scholar] [CrossRef] [PubMed]
- Efimenko, A.; Starostina, E.; Kalinina, N.; Stolzing, A. Angiogenic Properties of Aged Adipose Derived Mesenchymal Stem Cells after Hypoxic Conditioning. J. Transl. Med. 2011, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.D.; Dehez, S.; Arnaud, E.; Barreau, C.; Cazavet, A.; Perez, G.; Galinier, A.; Casteilla, L.; Planat-Bénard, V. Aging-Related Decrease of Human ASC Angiogenic Potential Is Reversed by Hypoxia Preconditioning through ROS Production. Mol. Ther. 2013, 21, 399–408. [Google Scholar] [CrossRef]
- Wu, H.; Li, J.Z.; Xie, B.D.; Tian, H.; Fang, S.H.; Jiang, S.L.; Kang, K. Lower Senescence of Adipose-Derived Stem Cells than Donor-Matched Bone Marrow Stem Cells for Surgical Ventricular Restoration. Stem Cells Dev. 2018, 27, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Klomjit, N.; Conley, S.M.; Zhu, X.Y.; Sadiq, I.M.; Libai, Y.; Krier, J.D.; Ferguson, C.M.; Jordan, K.L.; Tang, H.; Lerman, A.; et al. Effects of Obesity on Reparative Function of Human Adipose Tissue-Derived Mesenchymal Stem Cells on Ischemic Murine Kidneys. Int. J. Obes. 2022, 46, 1222–1233. [Google Scholar] [CrossRef]
- De Lange, P.; Lombardi, A.; Silvestri, E.; Cioffi, F.; Giacco, A.; Iervolino, S.; Petito, G.; Senese, R.; Lanni, A.; Moreno, M. Physiological Approaches Targeting Cellular and Mitochondrial Pathways Underlying Adipose Organ Senescence. Int. J. Mol. Sci. 2023, 24, 11676. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Xu, N.; Xu, D.; Dong, M.; Shi, X.; Wang, Y.; Hao, Z.; Zhu, S.; Zhao, D.; Jin, H.; et al. Comparison of Senescence-Related Changes between Three-and Two-Dimensional Cultured Adipose-Derived Mesenchymal Stem Cells. Stem Cell Res. Ther. 2020, 11, 226. [Google Scholar] [CrossRef] [PubMed]
- Truong, N.C.; Bui, K.H.T.; Van Pham, P. Characterization of Senescence of Human Adipose-Derived Stem Cells After Long-Term Expansion. Adv. Exp. Med. Biol. 2019, 1084, 109–128. [Google Scholar] [CrossRef]
- Mullen, M.; Nelson, A.L.; Goff, A.; Billings, J.; Kloser, H.; Huard, C.; Mitchell, J.; Hambright, W.S.; Ravuri, S.; Huard, J. Fisetin Attenuates Cellular Senescence Accumulation During Culture Expansion of Human Adipose-Derived Stem Cells. Stem Cells 2023, 41, 698–710. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics Improve Physical Function and Increase Lifespan in Old Age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Yousefzadeh, M.J.; Zhu, Y.; McGowan, S.J.; Angelini, L.; Fuhrmann-Stroissnigg, H.; Xu, M.; Ling, Y.Y.; Melos, K.I.; Pirtskhalava, T.; Inman, C.L.; et al. Fisetin Is a Senotherapeutic That Extends Health and Lifespan. EBioMedicine 2018, 36, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.J.; Liu, X.; Hu, M.; Zhang, Y.; Shi, P.Z.; Wang, J.W.; Lu, X.H.; Cheng, X.F.; Tao, Y.P.; Feng, X.M.; et al. Quercetin Ameliorates Oxidative Stress-Induced Senescence in Rat Nucleus Pulposus-Derived Mesenchymal Stem Cells via the MiR-34a-5p/SIRT1 Axis. World J. Stem Cells 2023, 15, 842–865. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T.; Tuday, E.; Allen, S.; Kim, J.; Trott, D.W.; Holland, W.L.; Donato, A.J.; Lesniewski, L.A. Senolytic Drugs, Dasatinib and Quercetin, Attenuate Adipose Tissue Inflammation, and Ameliorate Metabolic Function in Old Age. Aging Cell 2023, 22, e13767. [Google Scholar] [CrossRef] [PubMed]
- Hickson, L.T.J.; Langhi Prata, L.G.P.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Senolytics Decrease Senescent Cells in Humans: Preliminary Report from a Clinical Trial of Dasatinib plus Quercetin in Individuals with Diabetic Kidney Disease. EBioMedicine 2019, 47, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Doshida, Y.; Sano, H.; Iwabuchi, S.; Aigaki, T.; Yoshida, M.; Hashimoto, S.; Ishigami, A. Age-Associated Changes in the Transcriptomes of Non-Cultured Adipose-Derived Stem Cells from Young and Old Mice Assessed via Single-Cell Transcriptome Analysis. PLoS ONE 2020, 15, e0242171. [Google Scholar] [CrossRef]
- Suvakov, S.; Cubro, H.; White, W.M.; Butler Tobah, Y.S.; Weissgerber, T.L.; Jordan, K.L.; Zhu, X.Y.; Woollard, J.R.; Chebib, F.T.; Milic, N.M.; et al. Targeting Senescence Improves Angiogenic Potential of Adipose-Derived Mesenchymal Stem Cells in Patients with Preeclampsia. Biol. Sex Differ. 2019, 10, 49. [Google Scholar] [CrossRef]
- Lv, M.; Zhang, S.; Jiang, B.; Cao, S.; Dong, Y.; Cao, L.; Guo, S. Adipose-Derived Stem Cells Regulate Metabolic Homeostasis and Delay Aging by Promoting Mitophagy. FASEB J. 2021, 35, e21709. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Authors and Year of Publication | Type of Cells | Experimental Model | Key Findings | Experimental Settings | Reference |
|---|---|---|---|---|---|
| Yousefzadeh MJ, et al. 2018 | Murine and human fibroblasts | Various flavonoid polyphenols were tested for their senolytic activity on senescent fibroblasts from mice and humans, which had been induced by oxidative and genotoxic stress. | Fisetin has been shown to possess senotherapeutic properties, demonstrating effectiveness in reducing senescence in both mouse models and human tissue samples. | In vitro | [96] |
| Mullen M, et al. 2023 | hADSCs | Fisetin usage to Attenuates Cellular Senescence Accumulation During Culture Expansion | Fisetin has been effective in eliminating senescent cell populations from both early and late stage cell cultures without compromising their ability to differentiate into multiple cell types. | In vitro | [94] |
| Zhao WJ, et al. 2023 | Rat-ADSCs | Usage of Quercetin in treating oxidative stress-induced senescence in rat cells | Quercetin ameliorates oxidative stress-induced senescence in rat nucleus pulposus-derived mesenchymal stem cells via the miR-34a-5p/SIRT1 axis | In vitro | [97] |
| Hickson LJ, et al. 2019 | hADSCs | Usage of Dasatinib plus Quercetin | Dasatinib plus Quercetin usage decrease senescent cells in individuals with diabetic kidney disease | In vivo | [99] |
| Islam MT, et al. 2023 | Mice ADSCs | Twenty-one-month-old mice were administered Dasatinib and Quercetin through oral gavage for three consecutive days every two weeks over a period of three months. | Dasatinib and quercetin exhibit senolytic properties, effectively reducing the age-related accumulation of senescence markers. | In vitro | [98] |
| Suvakov S, et al. 2019 | hADSCs | hASCs exposure to TNF-alpha in vitro | Targeting senescence improves angiogenic potential of adipose-derived mesenchymal stem cells in patients with preeclampsia. | In vitro | [101] |
| Glasstetter LM, et al. 2023 | Swine and Human ADSCs | 5hmC profiles were examined through hydroxymethylated DNA immunoprecipitation sequencing followed by an integrative gene set enrichment analysis. | 467 hyper- and 591 hypo-hydroxymethylated loci in swine Obese- versus Lean-ASCs were found and were partly reversed in swine Obese-MSCs treated with vitamin-C. | In vitro | [69] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foti, R.; Storti, G.; Palmesano, M.; Scioli, M.G.; Fiorelli, E.; Terriaca, S.; Cervelli, G.; Kim, B.S.; Orlandi, A.; Cervelli, V. Senescence in Adipose-Derived Stem Cells: Biological Mechanisms and Therapeutic Challenges. Int. J. Mol. Sci. 2024, 25, 8390. https://doi.org/10.3390/ijms25158390
Foti R, Storti G, Palmesano M, Scioli MG, Fiorelli E, Terriaca S, Cervelli G, Kim BS, Orlandi A, Cervelli V. Senescence in Adipose-Derived Stem Cells: Biological Mechanisms and Therapeutic Challenges. International Journal of Molecular Sciences. 2024; 25(15):8390. https://doi.org/10.3390/ijms25158390
Chicago/Turabian StyleFoti, Riccardo, Gabriele Storti, Marco Palmesano, Maria Giovanna Scioli, Elena Fiorelli, Sonia Terriaca, Giulio Cervelli, Bong Sung Kim, Augusto Orlandi, and Valerio Cervelli. 2024. "Senescence in Adipose-Derived Stem Cells: Biological Mechanisms and Therapeutic Challenges" International Journal of Molecular Sciences 25, no. 15: 8390. https://doi.org/10.3390/ijms25158390
APA StyleFoti, R., Storti, G., Palmesano, M., Scioli, M. G., Fiorelli, E., Terriaca, S., Cervelli, G., Kim, B. S., Orlandi, A., & Cervelli, V. (2024). Senescence in Adipose-Derived Stem Cells: Biological Mechanisms and Therapeutic Challenges. International Journal of Molecular Sciences, 25(15), 8390. https://doi.org/10.3390/ijms25158390