
The GBA1 K198E Variant Is Associated with Suppression of Glucocerebrosidase Activity, Autophagy Impairment, Oxidative Stress, Mitochondrial Damage, and Apoptosis in Skin Fibroblasts
 , and
, and
Abstract
:1. Introduction
2. Results
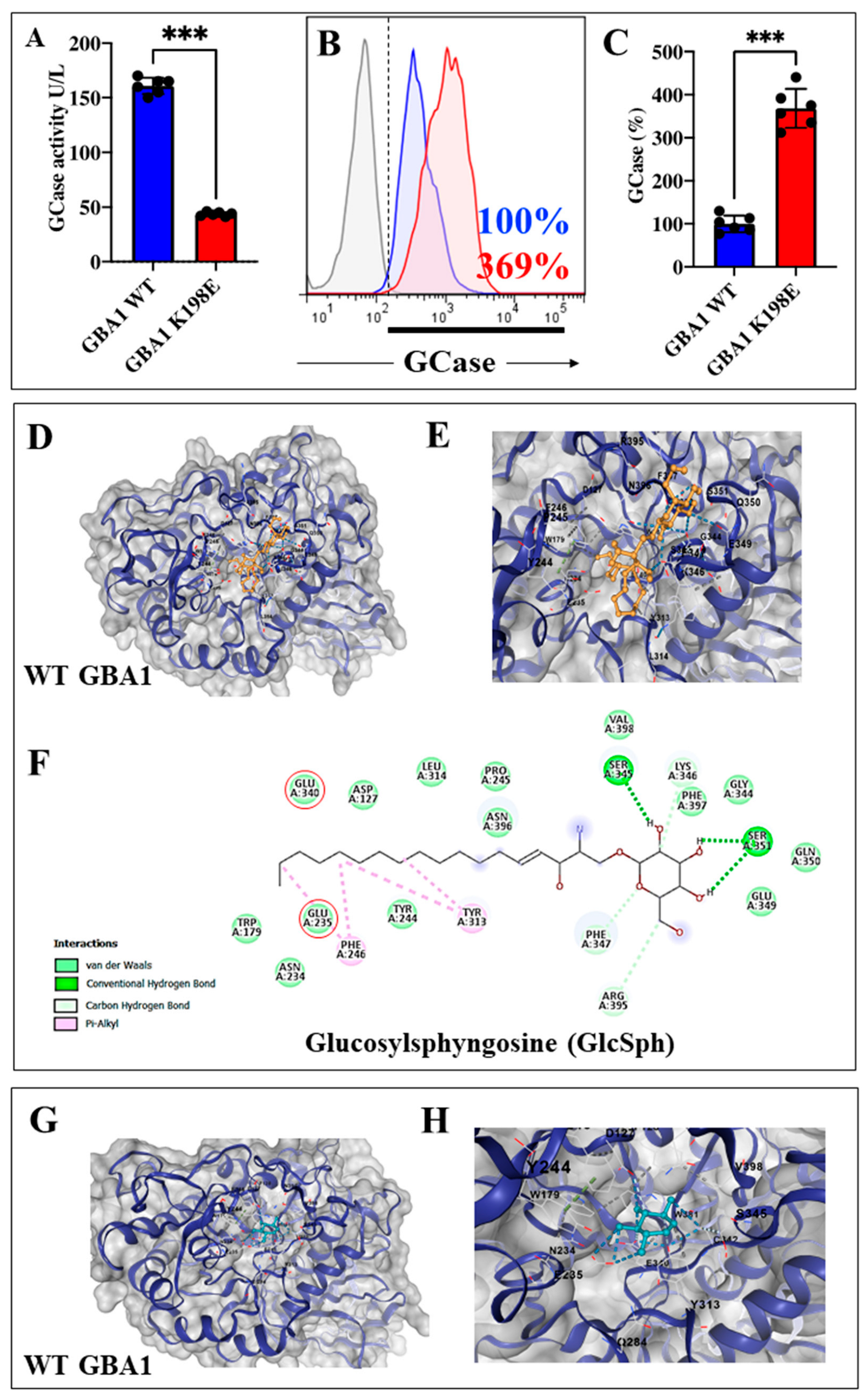
2.1. GBA1 K198E Variant Dramatically Inactivates the Catalytic Enzymatic Activity of GCase, but Did Not Affect the Protein Expression Levels
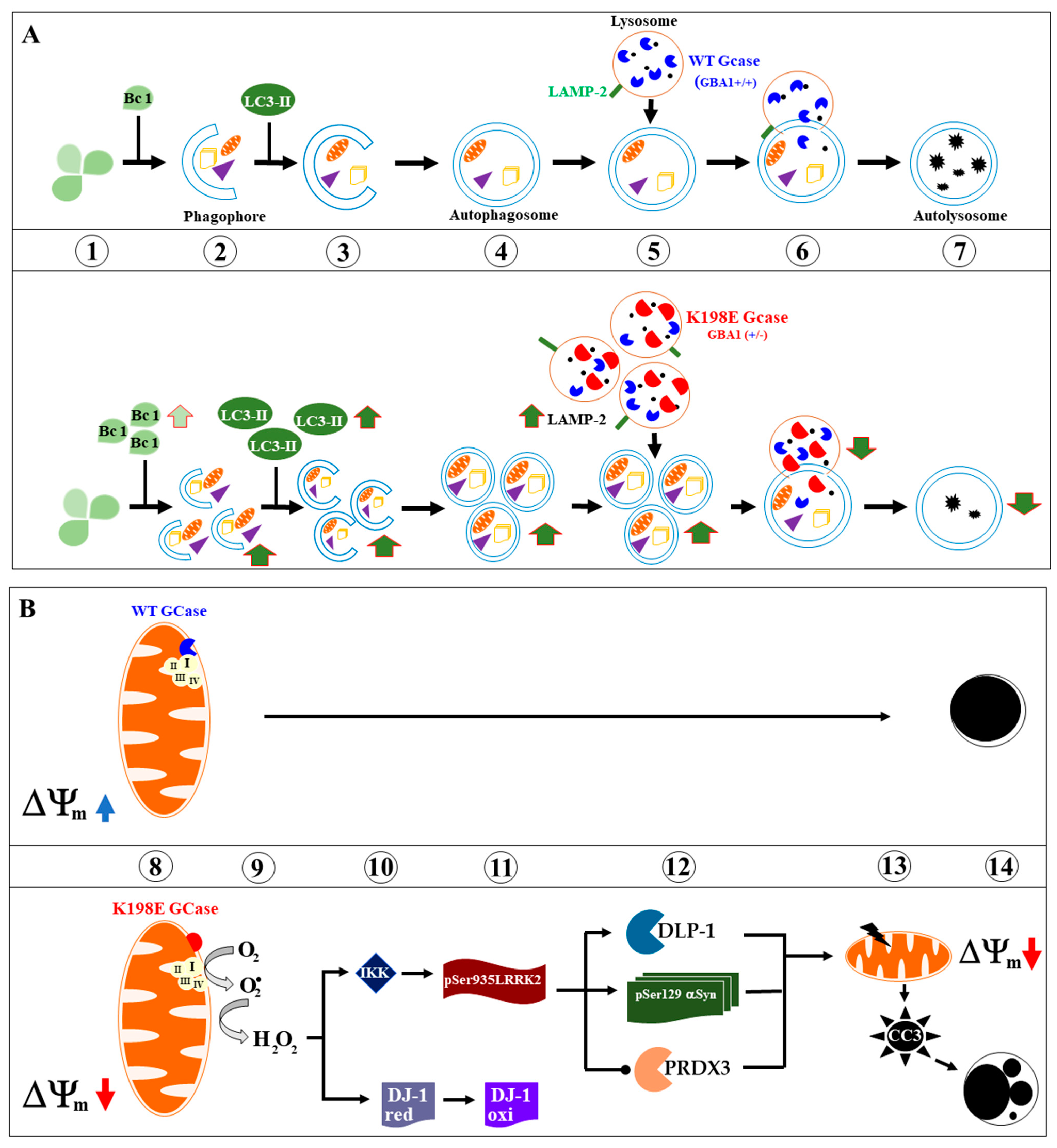
2.2. GBA1 K198E Shows an Acute GCase Deficiency in the Autophagy–Lysosome System
2.3. GBA1 K198E Increases the Expression of Beclin-1, LC3-II, and LAMP-2 in Fibroblasts
2.4. GBA1 K198E Variant Increases the Accumulation of Lysosomes and Decreases the Mitochondrial Membrane Potential (∆Ψm) in Mutant Fibroblasts, while Rotenone Increases the Damage
2.5. GBA1 K198E Fibroblasts Show Parkin Protein Colocalization with Mitochodrial TOM20 Proteins
2.6. GBA1 K198E Fibroblasts Show High Oxidized DJ-1-Cys106-SOH into DJ-1 Cys106SO3
2.7. GBA1 K198E Fibroblasts Show Endogenously High Phosphorylated LRRK2 at Ser935 along with Phosphorylated α-Synuclein (α-Syn) at Pathological Residue Ser129
2.8. GBA1 K198E Fibroblasts Show an Endogenously High Percentage of Cleaved Caspase 3 (CC3) Compared to WT Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Human Dermal Fibroblast Culture
4.2. Analysis of Cells
4.2.1. Assay Protocol
4.2.2. Glucocerebrosidase (GCase) Activity Assay
4.2.3. Autophagy Assay
4.3. Flow Cytometry and Fluorescent Microscopy Immunofluorescence
4.4. Characterization of Lysosomal Complexity
4.5. Analysis of Mitochondrial Membrane Potential (ΔΨm)
4.6. Molecular Docking
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parkinson, J. An Essay on the Shaking Palsy. J. Neuropsychiatry Clin. Neurosci. 2002, 14, 222–223. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, E.R.; Sherer, T.; Okun, M.S.; Bloemd, B.R. The Emerging Evidence of the Parkinson Pandemic. J. Parkinson’s Dis. 2018, 8, S3–S8. [Google Scholar] [CrossRef]
- Dorsey, E.R.; Elbaz, A.; Nichols, E.; Abbasi, N.; Abd-Allah, F.; Abdelalim, A.; Adsuar, J.C.; Ansha, M.G.; Brayne, C.; Choi, J.-Y.J.; et al. Global, Regional, and National Burden of Parkinson’s Disease, 1990–2016: A Systematic Analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Pan, J.; Tang, S.; Duan, D.; Yu, D.; Nong, H.; Wang, Z. Global Trends in the Incidence, Prevalence, and Years Lived with Disability of Parkinson’s Disease in 204 Countries/Territories from 1990 to 2019. Front. Public Health 2021, 9, 776847. [Google Scholar] [CrossRef] [PubMed]
- Váradi, C. Clinical Features of Parkinson’s Disease: The Evolution of Critical Symptoms. Biology 2020, 9, 103. [Google Scholar] [CrossRef]
- Lysia, S. Forno Neuropathology of Parkinson’s Disease. J. Neuropathol. Exp. Neurol. 1996, 55, 259–272. [Google Scholar] [CrossRef]
- Goedert, M.; Spillantini, M.G.; Del Tredici, K.; Braak, H. 100 Years of Lewy Pathology. Nat. Rev. Neurol. 2013, 9, 13–24. [Google Scholar] [CrossRef]
- Engelhardt, E.; Gomes, M.d.M. Lewy and His Inclusion Bodies: Discovery and Rejection. Dement. Neuropsychol. 2017, 11, 198–201. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Crowther, R.A.; Jakes, R.; Hasegawa, M.; Goedert, M. α-Synuclein in Filamentous Inclusions of Lewy Bodies from Parkinson’s Disease and Dementia with Lewy Bodies. Proc. Natl. Acad. Sci. USA 1998, 95, 6469–6473. [Google Scholar] [CrossRef]
- Islam, M.S.; Azim, F.; Saju, H.; Zargaran, A.; Shirzad, M.; Kamal, M.; Fatema, K.; Rehman, S.; Azad, M.A.M.; Ebrahimi-Barough, S. Pesticides and Parkinson’s Disease: Current and Future Perspective. J. Chem. Neuroanat. 2021, 115, 101966. [Google Scholar] [CrossRef]
- Bloem, B.R.; Boonstra, T.A. The Inadequacy of Current Pesticide Regulations for Protecting Brain Health: The Case of Glyphosate and Parkinson’s Disease. Lancet Planet. Health 2023, 7, e948–e949. [Google Scholar] [CrossRef]
- Bogers, J.S.; Bloem, B.R.; Den Heijer, J.M. The Etiology of Parkinson’s Disease: New Perspectives from Gene-Environment Interactions. J. Parkinson’s Dis. 2023, 13, 1281–1288. [Google Scholar] [CrossRef]
- Lim, S.-Y.; Klein, C. Parkinson’s Disease Is Predominantly a Genetic Disease. J. Parkinson’s Dis. 2024, 14, 467–482. [Google Scholar] [CrossRef]
- Pang, S.Y.-Y.; Ho, P.W.-L.; Liu, H.-F.; Leung, C.-T.; Li, L.; Chang, E.E.S.; Ramsden, D.B.; Ho, S.-L. The Interplay of Aging, Genetics and Environmental Factors in the Pathogenesis of Parkinson’s Disease. Transl. Neurodegener. 2019, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Fellner, A.; Kumar, K.R. Monogenic Parkinson’s Disease: Genotype, Phenotype, Pathophysiology, and Genetic Testing. Genes 2022, 13, 471. [Google Scholar] [CrossRef]
- Day, J.O.; Mullin, S. The Genetics of Parkinson’s Disease and Implications for Clinical Practice. Genes 2021, 12, 1006. [Google Scholar] [CrossRef] [PubMed]
- Matsui, H.; Takahashi, R. Current Trends in Basic Research on Parkinson’s Disease: From Mitochondria, Lysosome to α-Synuclein. J. Neural. Transm. 2024, 131, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, J.; Tan, E.K. Parkinson’s Disease: Etiopathogenesis and Treatment. J. Neurol. Neurosurg. Psychiatry 2020, 91, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Dickson, D.W. Parkinson’s Disease: Experimental Models and Reality. Acta Neuropathol. 2018, 135, 13–32. [Google Scholar] [CrossRef]
- Airavaara, M.; Parkkinen, I.; Konovalova, J.; Albert, K.; Chmielarz, P.; Domanskyi, A. Back and to the Future: From Neurotoxin-Induced to Human Parkinson’s Disease Models. Curr. Protoc. Neurosci. 2020, 91, e88. [Google Scholar] [CrossRef]
- Lawana, V.; Cannon, J.R. Rotenone Neurotoxicity: Relevance to Parkinson’s Disease. In Advances in Neurotoxicology; Academic Press: Cambridge, MA, USA, 2020; pp. 209–254. [Google Scholar]
- Teves, J.M.Y.; Bhargava, V.; Kirwan, K.R.; Corenblum, M.J.; Justiniano, R.; Wondrak, G.T.; Anandhan, A.; Flores, A.J.; Schipper, D.A.; Khalpey, Z.; et al. Parkinson’s Disease Skin Fibroblasts Display Signature Alterations in Growth, Redox Homeostasis, Mitochondrial Function, and Autophagy. Front. Neurosci. 2018, 11, 737. [Google Scholar] [CrossRef]
- Deus, C.M.; Pereira, S.P.; Cunha-Oliveira, T.; Pereira, F.B.; Raimundo, N.; Oliveira, P.J. Mitochondrial Remodeling in Human Skin Fibroblasts from Sporadic Male Parkinson’s Disease Patients Uncovers Metabolic and Mitochondrial Bioenergetic Defects. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2020, 1866, 165615. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M.; Wilder, S.; Horowitz, Z.; Reiner, O.; Gelbart, T.; Beutler, E. The Human Glucocerebrosidase Gene and Pseudogene: Structure and Evolution. Genomics 1989, 4, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Dvir, H.; Harel, M.; McCarthy, A.A.; Toker, L.; Silman, I.; Futerman, A.H.; Sussman, J.L. X-ray Structure of Human Acid-β-glucosidase, the Defective Enzyme in Gaucher Disease. EMBO Rep. 2003, 4, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Reczek, D.; Schwake, M.; Schröder, J.; Hughes, H.; Blanz, J.; Jin, X.; Brondyk, W.; Van Patten, S.; Edmunds, T.; Saftig, P. LIMP-2 Is a Receptor for Lysosomal Mannose-6-Phosphate-Independent Targeting of β-Glucocerebrosidase. Cell 2007, 131, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Valeiras, M.; Sidransky, E.; Tayebi, N. Lysosomal Integral Membrane Protein-2: A New Player in Lysosome-Related Pathology. Mol. Genet. Metab. 2014, 111, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Tamargo, R.J.; Velayati, A.; Goldin, E.; Sidransky, E. The Role of Saposin C in Gaucher Disease. Mol. Genet. Metab. 2012, 106, 257–263. [Google Scholar] [CrossRef]
- Atrian, S.; López-Viñas, E.; Gómez-Puertas, P.; Chabás, A.; Vilageliu, L.; Grinberg, D. An Evolutionary and Structure-based Docking Model for Glucocerebrosidase–Saposin C and Glucocerebrosidase–Substrate Interactions—Relevance for Gaucher Disease. Proteins Struct. Funct. Bioinform. 2008, 70, 882–891. [Google Scholar] [CrossRef]
- Kompoliti, K.; Verhagen, L. Encyclopedia of Movement Disorders; Academic Press: Cambridge, MA, USA, 2010; ISBN 9780123741059. [Google Scholar]
- Vieira, S.R.L.; Schapira, A.H.V. Glucocerebrosidase Mutations: A Paradigm for Neurodegeneration Pathways. Free. Radic. Biol. Med. 2021, 175, 42–55. [Google Scholar] [CrossRef]
- Vieira, S.R.L.; Schapira, A.H.V. Glucocerebrosidase Mutations and Parkinson Disease. J. Neural. Transm. 2022, 129, 1105–1117. [Google Scholar] [CrossRef]
- Migdalska-Richards, A.; Schapira, A.H.V. The Relationship between Glucocerebrosidase Mutations and Parkinson Disease. J. Neurochem. 2016, 139, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Menozzi, E.; Schapira, A.H.V. Exploring the Genotype–Phenotype Correlation in GBA-Parkinson Disease: Clinical Aspects, Biomarkers, and Potential Modifiers. Front. Neurol. 2021, 12, 694764. [Google Scholar] [CrossRef]
- Onal, G.; Yalçın-Çakmaklı, G.; Özçelik, C.E.; Boussaad, I.; Şeker, U.Ö.Ş.; Fernandes, H.J.R.; Demir, H.; Krüger, R.; Elibol, B.; Dökmeci, S.; et al. Variant-Specific Effects of GBA1 Mutations on Dopaminergic Neuron Proteostasis. J. Neurochem. 2024. [Google Scholar] [CrossRef]
- Velez-Pardo, C.; Lorenzo-Betancor, O.; Jimenez-Del-Rio, M.; Moreno, S.; Lopera, F.; Cornejo-Olivas, M.; Torres, L.; Inca-Martinez, M.; Mazzetti, P.; Cosentino, C.; et al. The Distribution and Risk Effect of GBA Variants in a Large Cohort of PD Patients from Colombia and Peru. Park. Relat. Disord. 2019, 63, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Orvisky, E.; Park, J.K.; Parker, A.; Walker, J.M.; Martin, B.M.; Stubblefield, B.K.; Uyama, E.; Tayebi, N.; Sidransky, E. The Identification of Eight Novel Glucocerebrosidase (GBA) Mutations in Patients with Gaucher Disease. Hum. Mutat. 2002, 19, 458–459. [Google Scholar] [CrossRef]
- Chatterjee, D.; Krainc, D. Mechanisms of Glucocerebrosidase Dysfunction in Parkinson’s Disease. J. Mol. Biol. 2023, 435, 168023. [Google Scholar] [CrossRef] [PubMed]
- Perez-Abshana, L.P.; Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Rotenone Blocks the Glucocerebrosidase Enzyme and Induces the Accumulation of Lysosomes and Autophagolysosomes Independently of LRRK2 Kinase in HEK-293 Cells. Int. J. Mol. Sci. 2023, 24, 10589. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, J.C.C.; Mano, G.B.C.; da Cunha Barreto-Vianna, A.R.; Garcia, T.F.M.; de Vasconcelos, A.V.; Sá, C.S.G.; de Souza Santana, S.L.; Farias, A.G.P.; Seimaru, B.; Lima, M.P.P.; et al. The Molecular Impact of Glucosylceramidase Beta 1 (Gba1) in Parkinson’s Disease: A New Genetic State of the Art. Mol. Neurobiol. 2024. [Google Scholar] [CrossRef]
- Zhang, J. Teaching the Basics of Autophagy and Mitophagy to Redox Biologists—Mechanisms and Experimental Approaches. Redox. Biol. 2015, 4, 242–259. [Google Scholar] [CrossRef]
- Lőrincz, P.; Juhász, G. Autophagosome-Lysosome Fusion. J. Mol. Biol. 2020, 432, 2462–2482. [Google Scholar] [CrossRef]
- Wipperman, M.F.; Montrose, D.C.; Gotto, A.M.; Hajjar, D.P. Mammalian Target of Rapamycin. Am. J. Pathol. 2019, 189, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Lippai, M.; Szatmári, Z. Autophagy—From Molecular Mechanisms to Clinical Relevance. Cell. Biol. Toxicol. 2017, 33, 145–168. [Google Scholar] [CrossRef]
- Dikic, I.; Elazar, Z. Mechanism and Medical Implications of Mammalian Autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular Mechanisms of Cell Death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.M.; Drouin-Ouellet, J.; Kuan, W.-L.; Cox, T.; Barker, R.A. Dermal Fibroblasts from Patients with Parkinson’s Disease Have Normal GCase Activity and Autophagy Compared to Patients with PD and GBA Mutations. F1000Research 2017, 6, 1751. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.E.; Newman, K.M.; Scheinker, V.; Berg-Fussman, A.; Grabowski, G.A. Analysis of Human Acid Beta-Glucosidase by Site-Directed Mutagenesis and Heterologous Expression. J. Biol. Chem. 1994, 269, 2283–2291. [Google Scholar] [CrossRef] [PubMed]
- Sawkar, A.R.; Adamski-Werner, S.L.; Cheng, W.-C.; Wong, C.-H.; Beutler, E.; Zimmer, K.-P.; Kelly, J.W. Gaucher Disease-Associated Glucocerebrosidases Show Mutation-Dependent Chemical Chaperoning Profiles. Chem. Biol. 2005, 12, 1235–1244. [Google Scholar] [CrossRef]
- Liou, B.; Kazimierczuk, A.; Zhang, M.; Scott, C.R.; Hegde, R.S.; Grabowski, G.A. Analyses of Variant Acid β-Glucosidases. J. Biol. Chem. 2006, 281, 4242–4253. [Google Scholar] [CrossRef]
- Premkumar, L.; Sawkar, A.R.; Boldin-Adamsky, S.; Toker, L.; Silman, I.; Kelly, J.W.; Futerman, A.H.; Sussman, J.L. X-ray Structure of Human Acid-β-Glucosidase Covalently Bound to Conduritol-B-Epoxide. J. Biol. Chem. 2005, 280, 23815–23819. [Google Scholar] [CrossRef]
- Pradas, E.; Martinez-Vicente, M. The Consequences of GBA Deficiency in the Autophagy–Lysosome System in Parkinson’s Disease Associated with GBA. Cells 2023, 12, 191. [Google Scholar] [CrossRef] [PubMed]
- Querfurth, H.; Lee, H.-K. Mammalian/Mechanistic Target of Rapamycin (MTOR) Complexes in Neurodegeneration. Mol. Neurodegener. 2021, 16, 44. [Google Scholar] [CrossRef]
- Wang, R.; Wang, J.; Hassan, A.; Lee, C.-H.; Xie, X.-S.; Li, X. Molecular Basis of V-ATPase Inhibition by Bafilomycin A1. Nat. Commun. 2021, 12, 1782. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.; Fairlie, W.D.; Lee, E.F. BECLIN1: Protein Structure, Function and Regulation. Cells 2021, 10, 1522. [Google Scholar] [CrossRef] [PubMed]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 Conjugation System in Mammalian Autophagy. Int. J. Biochem. Cell Biol. 2004, 36, 2503–2518. [Google Scholar] [CrossRef]
- Eskelinen, E.-L.; Illert, A.L.; Tanaka, Y.; Schwarzmann, G.; Blanz, J.; von Figura, K.; Saftig, P. Role of LAMP-2 in Lysosome Biogenesis and Autophagy. Mol. Biol. Cell 2002, 13, 3355–3368. [Google Scholar] [CrossRef] [PubMed]
- van Meel, E.; Bos, E.; van der Lienden, M.J.C.; Overkleeft, H.S.; van Kasteren, S.I.; Koster, A.J.; Aerts, J.M.F.G. Localization of Active Endogenous and Exogenous Β-glucocerebrosidase by Correlative Light-electron Microscopy in Human Fibroblasts. Traffic 2019, 20, 346–356. [Google Scholar] [CrossRef]
- Baden, P.; Perez, M.J.; Raji, H.; Bertoli, F.; Kalb, S.; Illescas, M.; Spanos, F.; Giuliano, C.; Calogero, A.M.; Oldrati, M.; et al. Glucocerebrosidase Is Imported into Mitochondria and Preserves Complex I Integrity and Energy Metabolism. Nat. Commun. 2023, 14, 1930. [Google Scholar] [CrossRef]
- Read, A.D.; Bentley, R.E.T.; Archer, S.L.; Dunham-Snary, K.J. Mitochondrial Iron–Sulfur Clusters: Structure, Function, and an Emerging Role in Vascular Biology. Redox. Biol. 2021, 47, 102164. [Google Scholar] [CrossRef]
- Pereira, C.S.; Teixeira, M.H.; Russell, D.A.; Hirst, J.; Arantes, G.M. Mechanism of Rotenone Binding to Respiratory Complex I Depends on Ligand Flexibility. Sci. Rep. 2023, 13, 6738. [Google Scholar] [CrossRef]
- Wang, X.-L.; Feng, S.-T.; Wang, Z.-Z.; Yuan, Y.-H.; Chen, N.-H.; Zhang, Y. Parkin, an E3 Ubiquitin Ligase, Plays an Essential Role in Mitochondrial Quality Control in Parkinson’s Disease. Cell. Mol. Neurobiol. 2021, 41, 1395–1411. [Google Scholar] [CrossRef]
- Araiso, Y.; Endo, T. Structural Overview of the Translocase of the Mitochondrial Outer Membrane Complex. Biophys. Physicobiol. 2022, 19, e190022. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, N.; Chisci, E.; Giovannoni, R. The Role of Hydrogen Peroxide in Redox-Dependent Signaling: Homeostatic and Pathological Responses in Mammalian Cells. Cells 2018, 7, 156. [Google Scholar] [CrossRef]
- Kinumi, T.; Kimata, J.; Taira, T.; Ariga, H.; Niki, E. Cysteine-106 of DJ-1 Is the Most Sensitive Cysteine Residue to Hydrogen Peroxide-Mediated Oxidation in Vivo in Human Umbilical Vein Endothelial Cells. Biochem. Biophys. Res. Commun. 2004, 317, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, Y.; Saito, Y. Oxidative Stress Biomaker and Its Application to Health Maintainance. J. Clin. Biochem. Nutr. 2014, 54, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Qing, H.; Wong, W.; McGeer, E.G.; McGeer, P.L. Lrrk2 Phosphorylates Alpha Synuclein at Serine 129: Parkinson Disease Implications. Biochem. Biophys. Res. Commun. 2009, 387, 149–152. [Google Scholar] [CrossRef]
- Sotomayor-Vivas, C.; Hernández-Lemus, E.; Dorantes-Gilardi, R. Linking Protein Structural and Functional Change to Mutation Using Amino Acid Networks. PLoS ONE 2022, 17, e0261829. [Google Scholar] [CrossRef]
- Rahman, A.A.; Morrison, B.E. Contributions of VPS35 Mutations to Parkinson’s Disease. Neuroscience 2019, 401, 1–10. [Google Scholar] [CrossRef]
- Bravo-San Pedro, J.M.; Niso-Santano, M.; Gómez-Sánchez, R.; Pizarro-Estrella, E.; Aiastui-Pujana, A.; Gorostidi, A.; Climent, V.; López de Maturana, R.; Sanchez-Pernaute, R.; López de Munain, A.; et al. The LRRK2 G2019S Mutant Exacerbates Basal Autophagy through Activation of the MEK/ERK Pathway. Cell. Mol. Life Sci. 2013, 70, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Menozzi, E.; Chau, K.; Schapira, A.H.V. Glucocerebrosidase 1 and Leucine-rich Repeat Kinase 2 in Parkinson Disease and Interplay between the Two Genes. J. Neurochem. 2021, 159, 826–839. [Google Scholar] [CrossRef]
- Kluss, J.H.; Beilina, A.; Williamson, C.D.; Lewis, P.A.; Cookson, M.R.; Bonet-Ponce, L. Lysosomal Positioning Regulates Rab10 Phosphorylation at LRRK2+ Lysosomes. Proc. Natl. Acad. Sci. USA 2022, 119, e2205492119. [Google Scholar] [CrossRef]
- Ysselstein, D.; Nguyen, M.; Young, T.J.; Severino, A.; Schwake, M.; Merchant, K.; Krainc, D. LRRK2 Kinase Activity Regulates Lysosomal Glucocerebrosidase in Neurons Derived from Parkinson’s Disease Patients. Nat. Commun. 2019, 10, 5570. [Google Scholar] [CrossRef] [PubMed]
- Funayama, M.; Nishioka, K.; Li, Y.; Hattori, N. Molecular Genetics of Parkinson’s Disease: Contributions and Global Trends. J. Hum. Genet. 2023, 68, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.A.; Fraile-Martinez, O.; de Leon-Oliva, D.; Boaru, D.L.; Lopez-Gonzalez, L.; García-Montero, C.; Alvarez-Mon, M.A.; Guijarro, L.G.; Torres-Carranza, D.; Saez, M.A.; et al. Autophagy in Its (Proper) Context: Molecular Basis, Biological Relevance, Pharmacological Modulation, and Lifestyle Medicine. Int. J. Biol. Sci. 2024, 20, 2532–2554. [Google Scholar] [CrossRef]
- Cleeter, M.W.J.; Chau, K.-Y.; Gluck, C.; Mehta, A.; Hughes, D.A.; Duchen, M.; Wood, N.W.; Hardy, J.; Mark Cooper, J.; Schapira, A.H. Glucocerebrosidase Inhibition Causes Mitochondrial Dysfunction and Free Radical Damage. Neurochem. Int. 2013, 62, 1–7. [Google Scholar] [CrossRef]
- Nechushtai, L.; Frenkel, D.; Pinkas-Kramarski, R. Autophagy in Parkinson’s Disease. Biomolecules 2023, 13, 1435. [Google Scholar] [CrossRef]
- Rubilar, J.C.; Outeiro, T.F.; Klein, A.D. The Lysosomal β-Glucocerebrosidase Strikes Mitochondria: Implications for Parkinson’s Therapeutics. Brain 2024, 2024, awae070. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.; Dawson, V.L.; Dawson, T.M. PINK1 and Parkin Mitochondrial Quality Control: A Source of Regional Vulnerability in Parkinson’s Disease. Mol. Neurodegener. 2020, 15, 20. [Google Scholar] [CrossRef]
- Stone, J.R.; Yang, S. Hydrogen Peroxide: A Signaling Messenger. Antioxid. Redox. Signal 2006, 8, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Kamikawaji, S.; Ito, G.; Iwatsubo, T. Identification of the Autophosphorylation Sites of LRRK2. Biochemistry 2009, 48, 10963–10975. [Google Scholar] [CrossRef]
- West, A.B.; Moore, D.J.; Choi, C.; Andrabi, S.A.; Li, X.; Dikeman, D.; Biskup, S.; Zhang, Z.; Lim, K.-L.; Dawson, V.L.; et al. Parkinson’s Disease-Associated Mutations in LRRK2 Link Enhanced GTP-Binding and Kinase Activities to Neuronal Toxicity. Hum. Mol. Genet. 2007, 16, 223–232. [Google Scholar] [CrossRef]
- Li, X.; Moore, D.J.; Xiong, Y.; Dawson, T.M.; Dawson, V.L. Reevaluation of Phosphorylation Sites in the Parkinson Disease-Associated Leucine-Rich Repeat Kinase 2. J. Biol. Chem. 2010, 285, 29569–29576. [Google Scholar] [CrossRef] [PubMed]
- Dzamko, N.; Inesta-Vaquera, F.; Zhang, J.; Xie, C.; Cai, H.; Arthur, S.; Tan, L.; Choi, H.; Gray, N.; Cohen, P.; et al. The IkappaB Kinase Family Phosphorylates the Parkinson’s Disease Kinase LRRK2 at Ser935 and Ser910 during Toll-Like Receptor Signaling. PLoS ONE 2012, 7, e39132. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, H.; Hasegawa, M.; Dohmae, N.; Kawashima, A.; Masliah, E.; Goldberg, M.S.; Shen, J.; Takio, K.; Iwatsubo, T. α-Synuclein Is Phosphorylated in Synucleinopathy Lesions. Nat. Cell Biol. 2002, 4, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Wang, L.; Liu, W.; Zhu, G.; Chen, Y.; Zhang, J. Biomarkers and the Role of α-Synuclein in Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 645996. [Google Scholar] [CrossRef]
- Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Neuroprotective Effect of the LRRK2 Kinase Inhibitor PF-06447475 in Human Nerve-like Differentiated Cells Exposed to Oxidative Stress Stimuli: Implications for Parkinson’s Disease. Neurochem. Res. 2016, 41, 2675–2692. [Google Scholar] [CrossRef]
- Giraldo-Berrio, D.; Mendivil-Perez, M.; Velez-Pardo, C.; Jimenez-Del-Rio, M. Rotenone Induces a Neuropathological Phenotype in Cholinergic-like Neurons Resembling Parkinson’s Disease Dementia (PDD). Neurotox. Res. 2024, 42, 28. [Google Scholar] [CrossRef]
- Quintero-Espinosa, D.A.; Sanchez-Hernandez, S.; Velez-Pardo, C.; Martin, F.; Jimenez-Del-Rio, M. LRRK2 Knockout Confers Resistance in HEK-293 Cells to Rotenone-Induced Oxidative Stress, Mitochondrial Damage, and Apoptosis. Int. J. Mol. Sci. 2023, 24, 10474. [Google Scholar] [CrossRef]
- Hartmann, A.; Hunot, S.; Michel, P.P.; Muriel, M.-P.; Vyas, S.; Faucheux, B.A.; Mouatt-Prigent, A.; Turmel, H.; Srinivasan, A.; Ruberg, M.; et al. Caspase-3: A Vulnerability Factor and Final Effector in Apoptotic Death of Dopaminergic Neurons in Parkinson’s Disease. Proc. Natl. Acad. Sci. USA 2000, 97, 2875–2880. [Google Scholar] [CrossRef]
- Monteiro, L.d.B.; Davanzo, G.G.; de Aguiar, C.F.; Moraes-Vieira, P.M.M. Using Flow Cytometry for Mitochondrial Assays. MethodsX 2020, 7, 100938. [Google Scholar] [CrossRef]
- Lazic, S.E.; Clarke-Williams, C.J.; Munafò, M.R. What Exactly Is ‘N’ in Cell Culture and Animal Experiments? PLoS Biol. 2018, 16, e2005282. [Google Scholar] [CrossRef]
- Maayan Eshed, G.; Alcalay, R.N. GBA1-and LRRK2-Directed Treatments: The Way Forward. Park. Relat. Disord. 2024, 122, 106039. [Google Scholar] [CrossRef] [PubMed]
- Auburger, G.; Klinkenberg, M.; Drost, J.; Marcus, K.; Morales-Gordo, B.; Kunz, W.S.; Brandt, U.; Broccoli, V.; Reichmann, H.; Gispert, S.; et al. Primary Skin Fibroblasts as a Model of Parkinson’s Disease. Mol. Neurobiol. 2012, 46, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Corenblum, M.; McRobbie-Johnson, A.; Carruth, E.; Bernard, K.; Luo, M.; Mandarino, L.; Peterson, S.; Sans-Fuentes, M.; Billheimer, D.; Maley, T.; et al. Parallel Neurodegenerative Phenotypes in Sporadic Parkinson’s Disease Fibroblasts and Midbrain Dopamine Neurons. Prog. Neurobiol. 2023, 229, 102501. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Submitted Protein * | Submitted Ligand ** | Vina Score *** | Cavity Volume (Å3) | Center (x, y, z) | Docking Size (x, y, z) | Contact Residues |
|---|---|---|---|---|---|---|
| WT GCase | Glucosylsphingosine CID: 5280570 | −6.1 | 447 | 9, −12, 0 | 28, 28, 28 | Chain A: ASP127 TRP179 ASN234 GLU235 TYR244 PRO245 PHE246 TYR313 LEU314 GLU340 GLY344 SER345 LYS346 PHE347 GLU349 GLN350 SER351 TRP381 ARG395 ASN396 PHE397 VAL398 |
| K198E GCase | Glucosylsphingosine CID: 5280570 | - | - | - | - | - |
| WT GCase | Conduritol B epoxide CID: 136345 | −5.7 | 447 | 9, −12, 0 | 16, 16, 16 | Chain A: ASP127 PHE128 TRP179 ASN234 GLU235 TYR244 PHE246 GLN284 TYR313 GLU340 CYS342 SER345 TRP381 ASN396 VAL398 |
| K198E GCase | Conduritol-B-epoxide CID: 136345 | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Abshana, L.P.; Mendivil-Perez, M.; Jimenez-Del-Rio, M.; Velez-Pardo, C. The GBA1 K198E Variant Is Associated with Suppression of Glucocerebrosidase Activity, Autophagy Impairment, Oxidative Stress, Mitochondrial Damage, and Apoptosis in Skin Fibroblasts. Int. J. Mol. Sci. 2024, 25, 9220. https://doi.org/10.3390/ijms25179220
Perez-Abshana LP, Mendivil-Perez M, Jimenez-Del-Rio M, Velez-Pardo C. The GBA1 K198E Variant Is Associated with Suppression of Glucocerebrosidase Activity, Autophagy Impairment, Oxidative Stress, Mitochondrial Damage, and Apoptosis in Skin Fibroblasts. International Journal of Molecular Sciences. 2024; 25(17):9220. https://doi.org/10.3390/ijms25179220
Chicago/Turabian StylePerez-Abshana, Laura Patricia, Miguel Mendivil-Perez, Marlene Jimenez-Del-Rio, and Carlos Velez-Pardo. 2024. "The GBA1 K198E Variant Is Associated with Suppression of Glucocerebrosidase Activity, Autophagy Impairment, Oxidative Stress, Mitochondrial Damage, and Apoptosis in Skin Fibroblasts" International Journal of Molecular Sciences 25, no. 17: 9220. https://doi.org/10.3390/ijms25179220