
Exploring the Effects and Potential Mechanisms of Hesperidin for the Treatment of CPT-11-Induced Diarrhea: Network Pharmacology, Molecular Docking, and Experimental Validation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
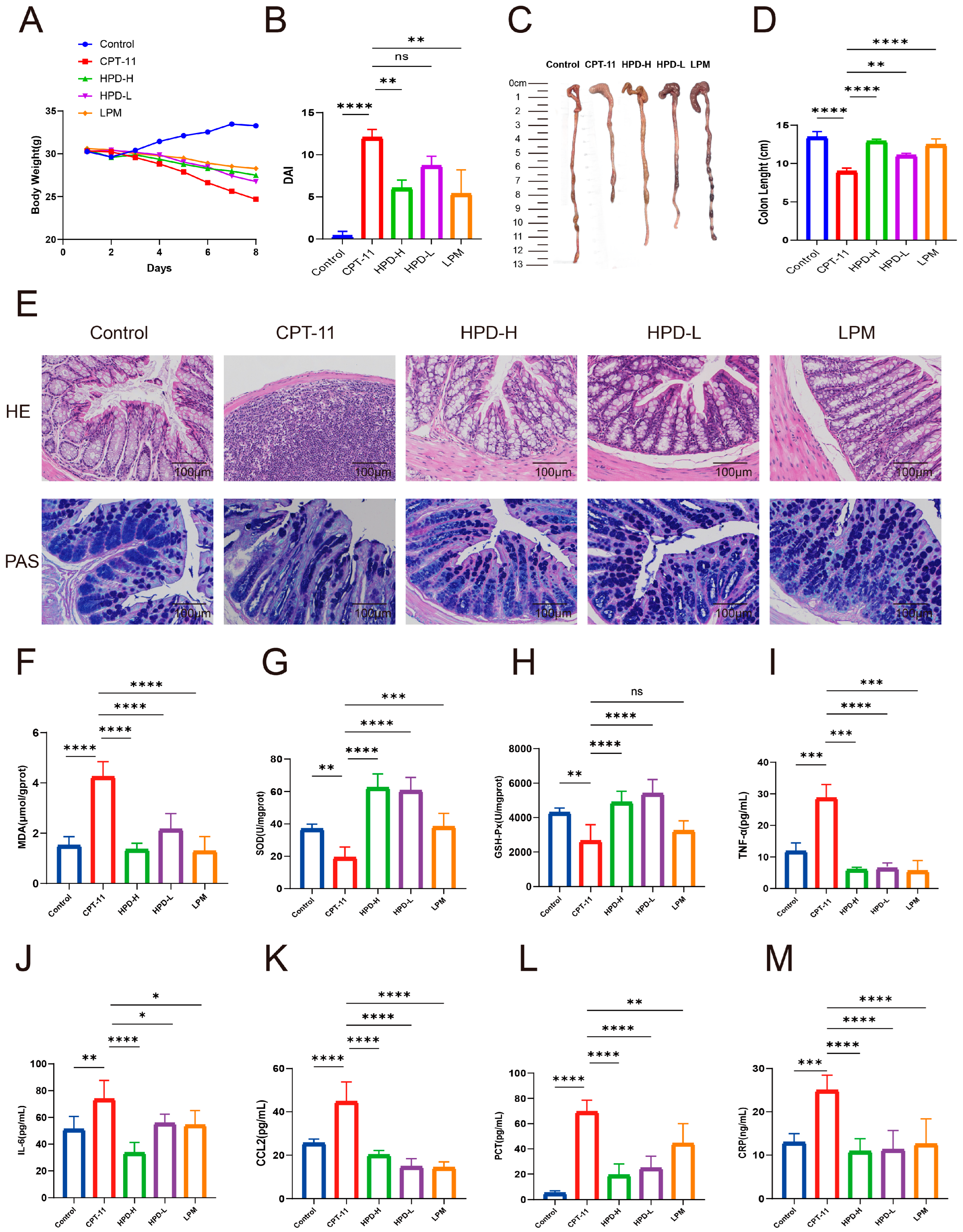
2.1. Amelioration of HPD on CID
2.1.1. HPD Ameliorates Diarrhea in CID Mice
2.1.2. HPD Attenuates Intestinal Pathology in CID Mice
2.1.3. HPD Attenuates Intestinal Oxidative Stress in CID Mice
2.1.4. HPD Attenuates Intestinal Inflammation in CID Mice
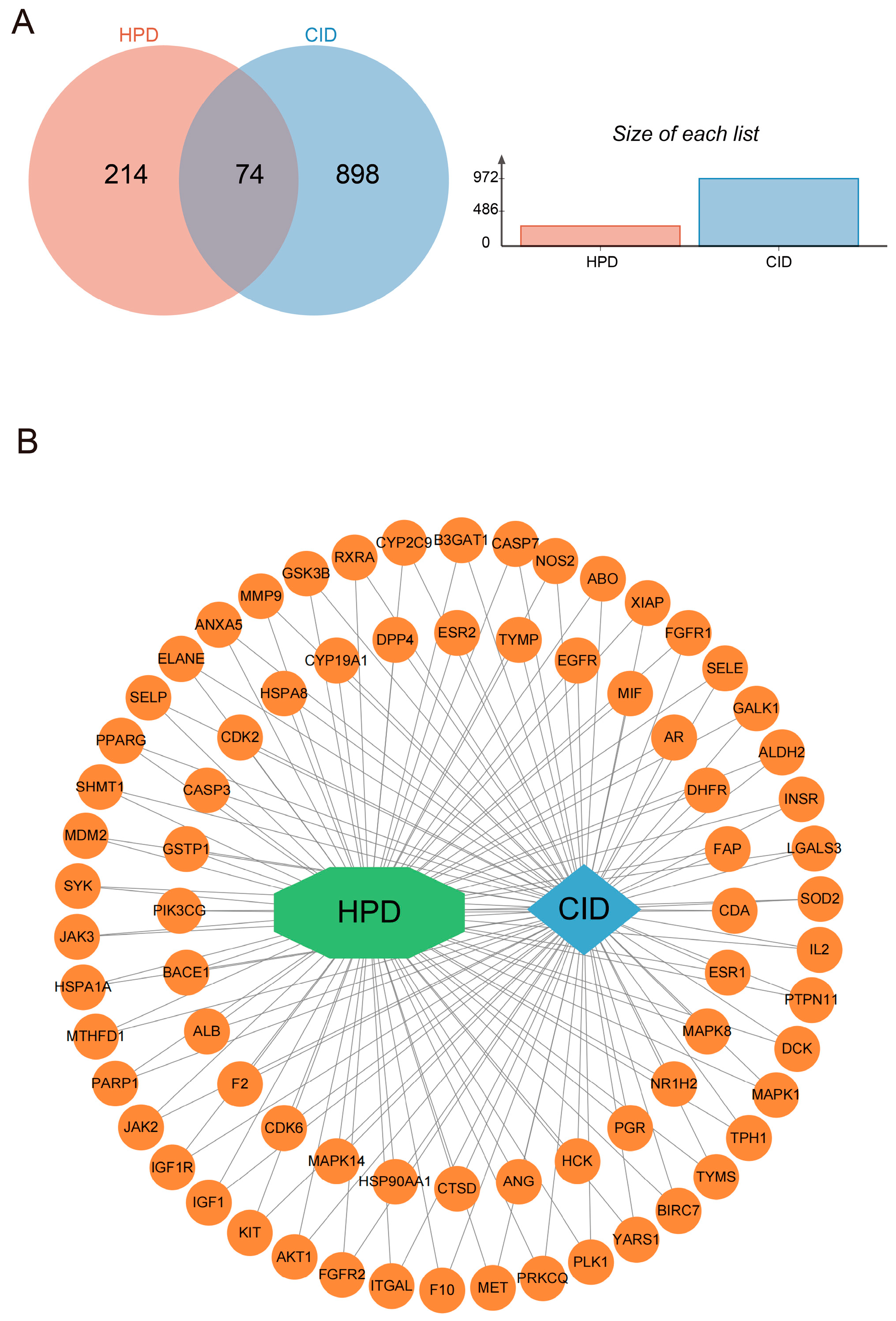
2.2. The Effect of HPD on CID Analyzed by Network Pharmacology
2.2.1. Network Analysis of Targets
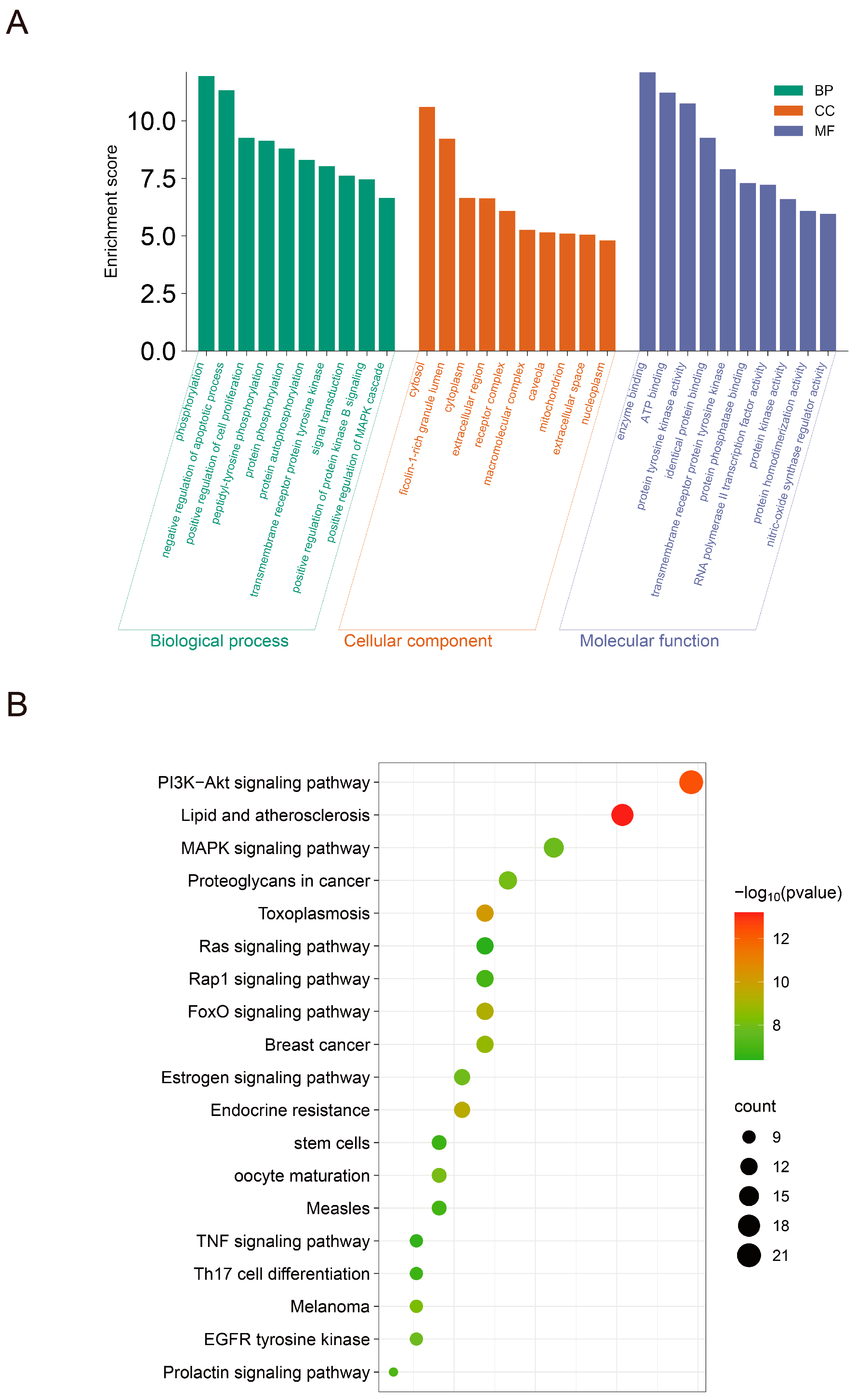
2.2.2. Enrichment Analysis of Overlapped Target
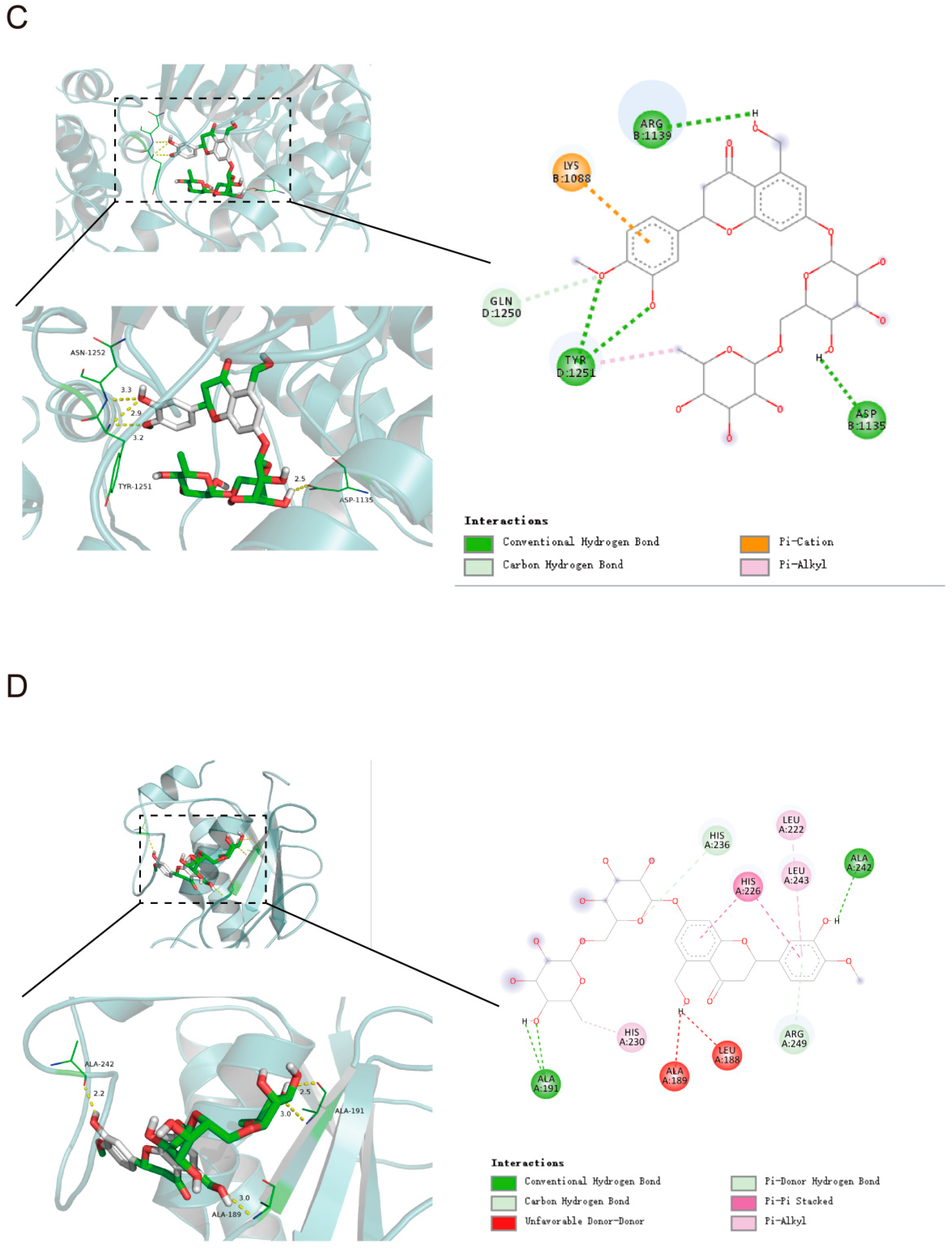
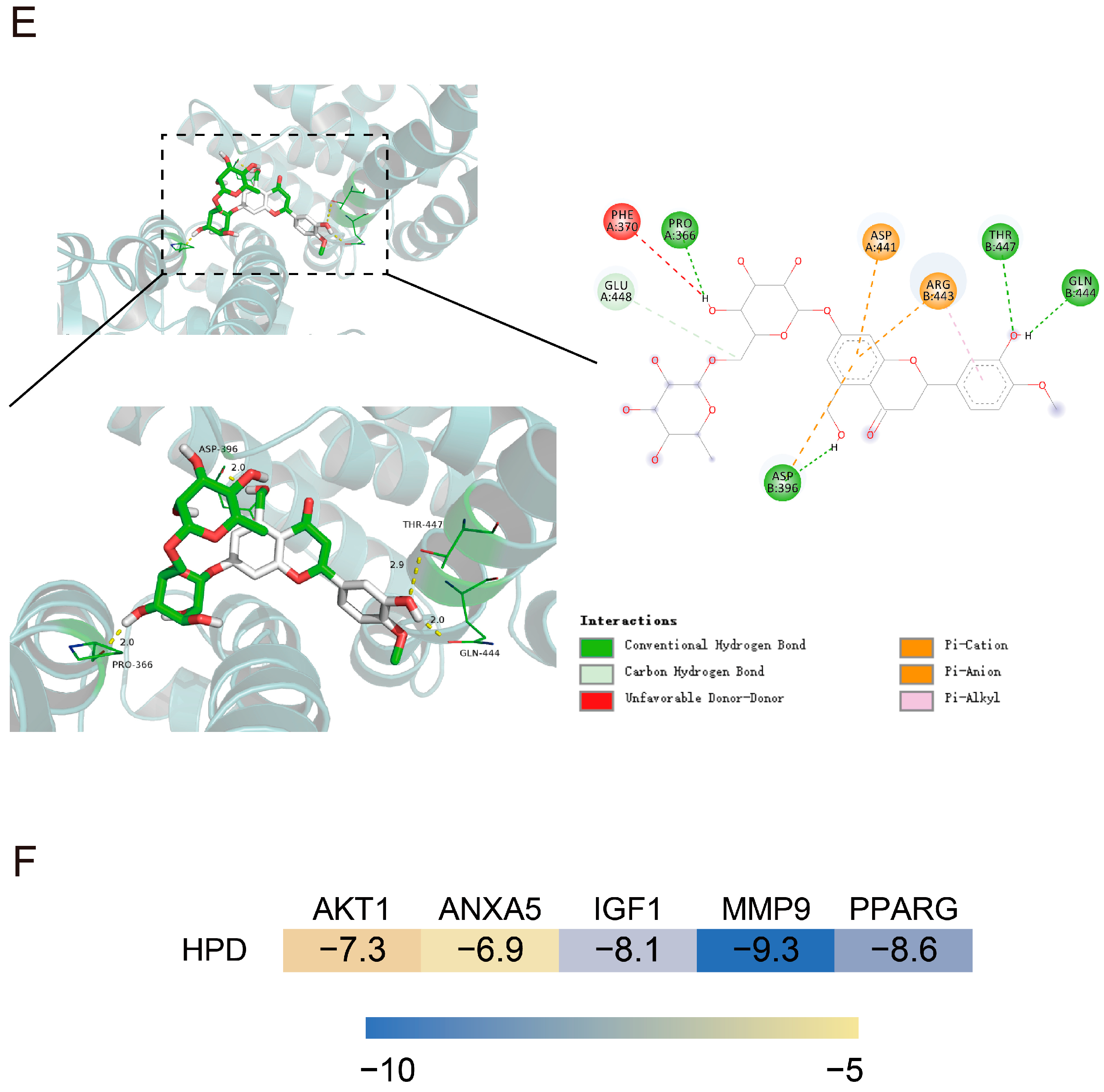
2.3. Molecular Docking Analysis
2.4. Verification of HPD on Alleviating CID
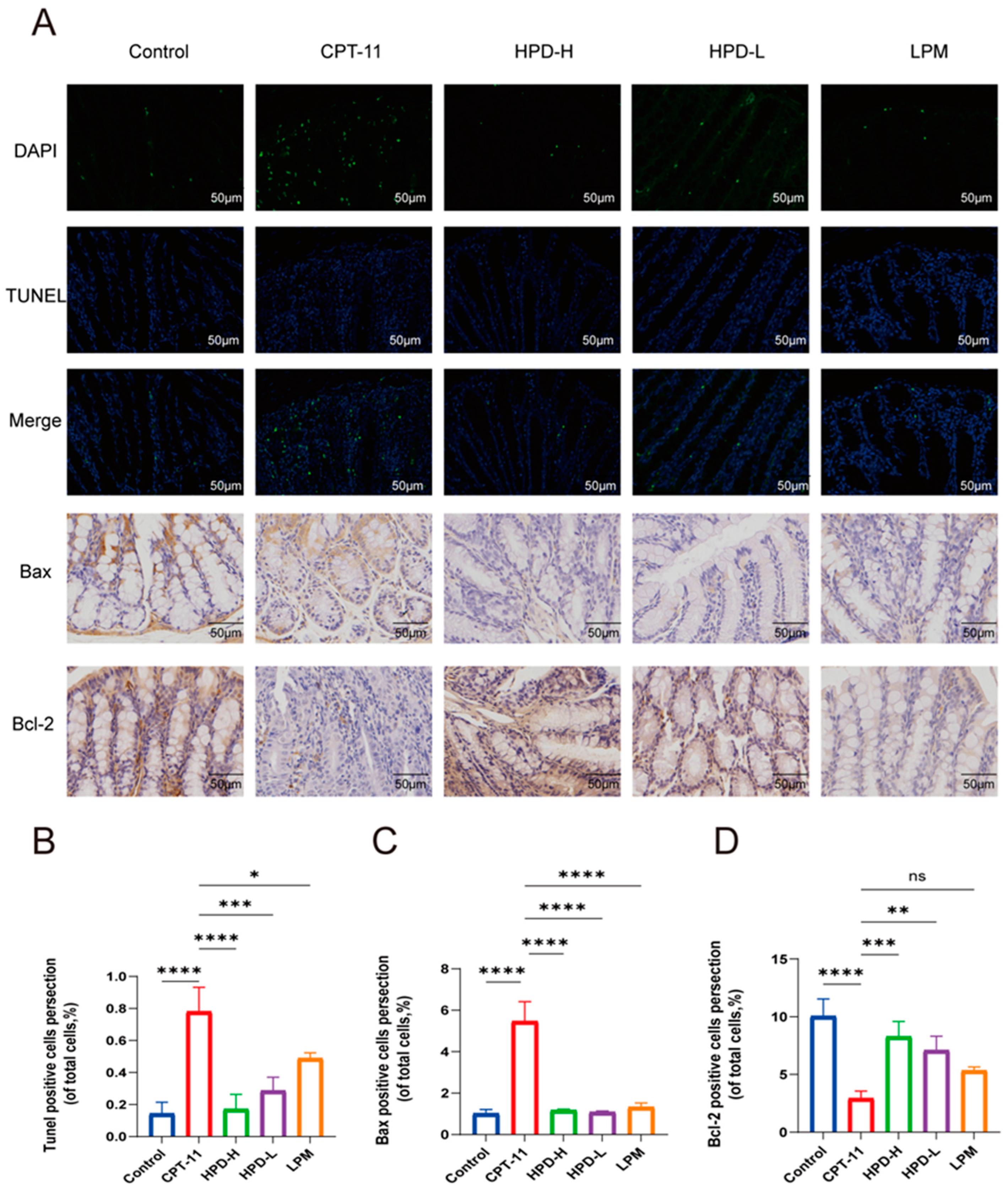
2.4.1. HPD Attenuates Enterocyte Apoptosis in CID Mice
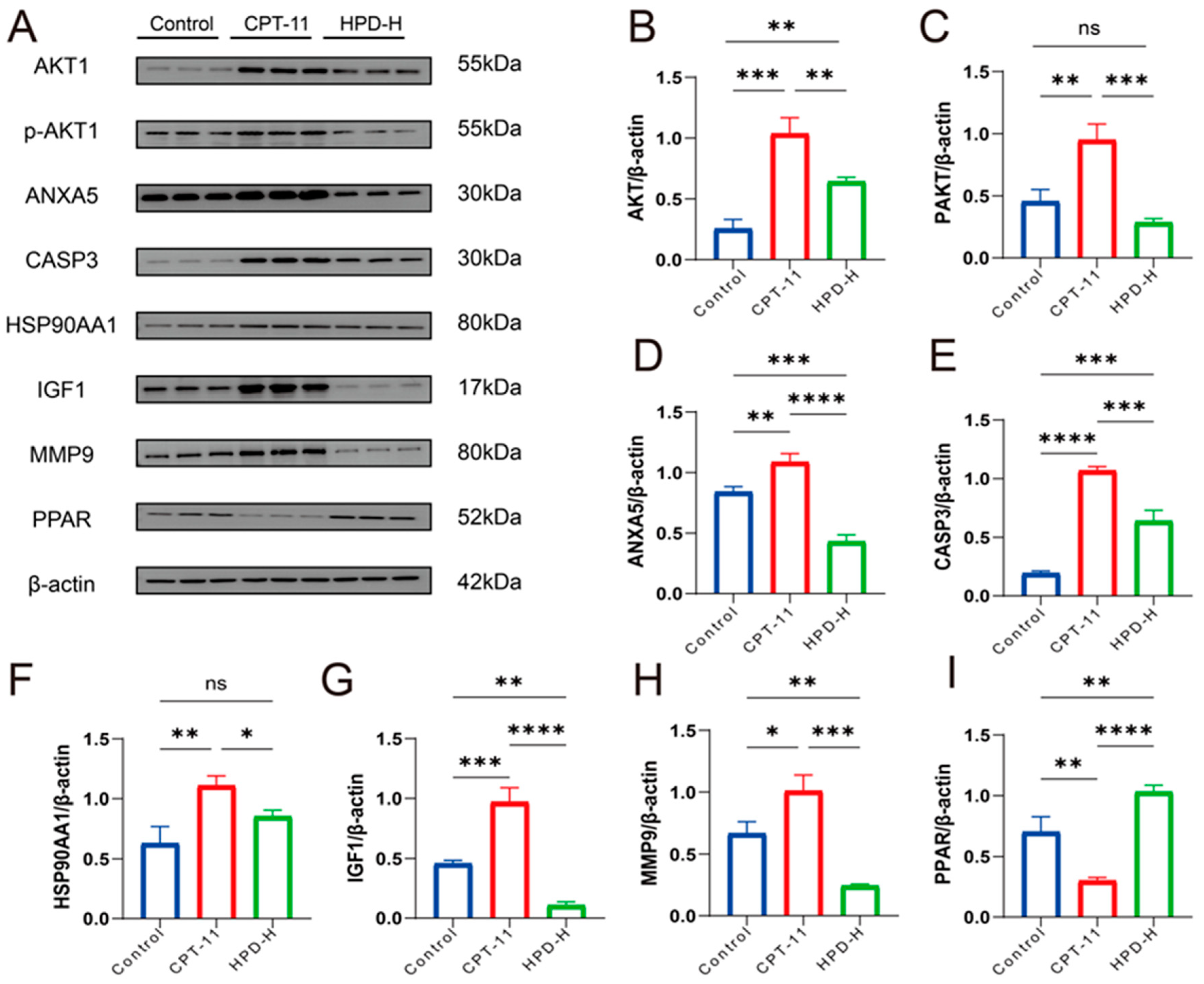
2.4.2. WB Results for 7 Key Proteins
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Animals
4.3. Experimental Design
4.4. Assessment of Severity of CID
4.5. Histopathological Examination
4.6. ELISA Determination
4.7. Network Pharmacology Analysis
4.7.1. HPD Target Prediction and Disease-Related Target Screening
4.7.2. Protein–Protein Interaction Network Analysis
4.7.3. GO and KEGG Pathway Enrichment Analyses
4.8. Molecular Docking
4.9. Immunohistochemical Analysis
4.10. Western Blotting
4.11. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Zraik, I.M.; Heß-Busch, Y. [Management of chemotherapy side effects and their long-term sequelae]. Urologe 2021, 60, 862–871. [Google Scholar] [CrossRef]
- Maroun, J.A.; Anthony, L.B.; Blais, N.; Burkes, R.; Dowden, S.D.; Dranitsaris, G.; Samson, B.; Shah, A.; Thirlwell, M.P.; Vincent, M.D.; et al. Prevention and management of chemotherapy-induced diarrhea in patients with colorectal cancer: A consensus statement by the Canadian Working Group on Chemotherapy-Induced Diarrhea. Curr. Oncol. 2007, 14, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Jia, Y.; Li, J.; Kuai, W.; Li, Y.; Guo, F.; Xu, X.; Zhao, Z.; Lv, J.; Li, Z. Gegen Qinlian decoction enhances the effect of PD-1 blockade in colorectal cancer with microsatellite stability by remodelling the gut microbiota and the tumour microenvironment. Cell Death Dis. 2019, 10, 415. [Google Scholar] [CrossRef]
- Jansman, F.G.; Sleijfer, D.T.; de Graaf, J.C.; Coenen, J.L.; Brouwers, J.R. Management of chemotherapy-induced adverse effects in the treatment of colorectal cancer. Drug Saf. 2001, 24, 353–367. [Google Scholar] [PubMed]
- Kawasaki, Y.; Kakimoto, K.; Tanaka, Y.; Shimizu, H.; Nishida, K.; Numa, K.; Kinoshita, N.; Tatsumi, Y.; Nakazawa, K.; Koshiba, R.; et al. Relationship between Chemotherapy-Induced Diarrhea and Intestinal Microbiome Composition. Digestion 2023, 104, 357–369. [Google Scholar] [CrossRef]
- Wadler, S.; Benson, A.B., III; Engelking, C.; Catalano, R.; Field, M.; Kornblau, S.M.; Mitchell, E.; Rubin, J.; Trotta, P.; Vokes, E. Recommended guidelines for the treatment of chemotherapy-induced diarrhea. J. Clin. Oncol. 1998, 16, 3169–3178. [Google Scholar] [CrossRef]
- Yu, Y.; Kong, R.; Cao, H.; Yin, Z.; Liu, J.; Nan, X.; Phan, A.T.; Ding, T.; Zhao, H.; Wong, S.T.C. Two birds, one stone: Hesperetin alleviates chemotherapy-induced diarrhea and potentiates tumor inhibition. Oncotarget 2018, 9, 27958. [Google Scholar] [CrossRef]
- Pyrzynska, K.A.-O. Hesperidin: A Review on Extraction Methods, Stability and Biological Activities. Nutrients 2022, 14, 2387. [Google Scholar] [CrossRef]
- Abuelsaad, A.S.; Mohamed, I.; Allam, G.; Al-Solumani, A.A. Antimicrobial and immunomodulating activities of hesperidin and ellagic acid against diarrheic Aeromonas hydrophila in a murine model. Life Sci. 2013, 93, 714–722. [Google Scholar] [CrossRef]
- Habauzit, V.; Morand, C. Evidence for a protective effect of polyphenols-containing foods on cardiovascular health: An update for clinicians. Ther. Adv. Chronic Dis. 2012, 3, 87–106. [Google Scholar] [CrossRef]
- Xu, L.; Yang, Z.L.; Li, P.; Zhou, Y.Q. Modulating effect of Hesperidin on experimental murine colitis induced by dextran sulfate sodium. Phytomedicine 2009, 16, 989–995. [Google Scholar] [CrossRef]
- Shafik, N.A.-O.; Gaber, R.A.; Mohamed, D.A.; Ebeid, A.M. Hesperidin modulates dextran sulfate sodium-induced ulcerative colitis in rats: Targeting sphingosine kinase-1- sphingosine 1 phosphate signaling pathway, mitochondrial biogenesis, inflammation, and apoptosis. J. Biochem. Mol. Toxicol. 2019, 33, e22312. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Zhou, Z.; Xiong, P.; Cheng, L.; Ma, J.; Wen, Y.; Shen, T.; He, X.; Wang, L.; et al. Integrated network pharmacology, metabolomics, and transcriptomics of Huanglian-Hongqu herb pair in non-alcoholic fatty liver disease. J. Ethnopharmacol. 2024, 10, 117828. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, B.; Chen, S.; Lin, M.; Chen, Y.; Jin, S.; Chen, W.; Zhang, Y. Applications of Network Pharmacology in Traditional Chinese Medicine Research. Evid. Based Complement. Altern. Med. 2020, 2020, 1646905. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, H.; Li, N.; Chen, J.; Xu, H.; Wang, Y.; Liang, Q. Network pharmacology, a promising approach to reveal the pharmacology mechanism of Chinese medicine formula. J. Ethnopharmacol. 2023, 309, 116306. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Shen, W.; Wang, Y.; Cao, Y.; Nuerbulati, N.; Chen, W.; Lu, G.; Xiao, W.; Qi, R. Grape Seed Polyphenols Ameliorated Dextran Sulfate Sodium-Induced Colitis via Suppression of Inflammation and Apoptosis. Pharmacology 2020, 105, 9–18. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, W.; Wu, Z.; Tian, X.; Xiang, J.; Li, L.; Li, Z.; Peng, X.; Wei, S.; Ma, X.; et al. Baicalin and the liver-gut system: Pharmacological bases explaining its therapeutic effects. Pharmacol. Res. 2021, 165, 105444. [Google Scholar] [CrossRef]
- Arbuckle, R.B.; Huber, S.L.; Zacker, C. The consequences of diarrhea occurring during chemotherapy for colorectal cancer: A retrospective study. Oncologist 2000, 5, 250–259. [Google Scholar] [CrossRef]
- Richardson, G.; Dobish, R. Chemotherapy induced diarrhea. J. Oncol. Pharm. Pract. 2007, 13, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Ha, L.; Arimori, K.; Latham, P.; Kobayashi, K.; Ceryak, S.; Matsuzaki, Y.; Bouscarel, B. Intestinal alkalization as a possible preventive mechanism in irinotecan (CPT-11)-induced diarrhea. Cancer Res. 2002, 62, 179–187. [Google Scholar] [PubMed]
- Huang, S.Y.; Zou, X. Advances and challenges in protein-ligand docking. Int. J. Mol. Sci. 2010, 11, 3016–3034. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zan, J.; Song, Y.; Yang, G.; Shang, H.; Zhao, W. Evaluation of intestinal injury, inflammatory response and oxidative stress following intracerebral hemorrhage in mice. Int. J. Mol. Med. 2018, 42, 2120–2128. [Google Scholar] [CrossRef] [PubMed]
- Izadparast, F.; Riahi-Zajani, B.; Yarmohammadi, F.; Hayes, A.W.; Karimi, G. Protective effect of berberine against LPS-induced injury in the intestine: A review. Cell Cycle. 2022, 21, 2365–2378. [Google Scholar] [CrossRef]
- Zhang, M.; Xia, F.; Xia, S.; Zhou, W.; Zhang, Y.; Han, X.; Zhao, K.; Feng, L.; Dong, R.; Tian, D.; et al. NSAID-Associated Small Intestinal Injury: An Overview from Animal Model Development to Pathogenesis, Treatment, and Prevention. Front. Pharmacol. 2022, 13, 818877. [Google Scholar] [CrossRef]
- Brandt, A.; Baumann, A.; Hernández-Arriaga, A.; Jung, F.; Nier, A.; Staltner, R.; Rajcic, D.; Schmeer, C.; Witte, O.W.; Wessner, B.; et al. Impairments of intestinal arginine and NO metabolisms trigger aging-associated intestinal barrier dysfunction and ‘inflammaging’. Redox Biol. 2022, 58, 102528. [Google Scholar] [CrossRef]
- Hasegawa, T.; Mizugaki, A.; Inoue, Y.; Kato, H.A.-O.X.; Murakami, H. Cystine reduces tight junction permeability and intestinal inflammation induced by oxidative stress in Caco-2 cells. Amino Acids 2021, 53, 1021–1032. [Google Scholar] [CrossRef]
- Ahmadi, A.; Shadboorestan, A. Oxidative stress and cancer; the role of hesperidin, a citrus natural bioflavonoid, as a cancer chemoprotective agent. Nutr. Cancer 2016, 68, 29–39. [Google Scholar] [CrossRef]
- Bailly, C. Irinotecan: 25 years of cancer treatment. Pharmacol. Res. 2019, 148, 104398. [Google Scholar] [CrossRef]
- Di Tommaso, N.; Gasbarrini, A.; Ponziani, F.A.-O. Intestinal Barrier in Human Health and Disease. Int. J. Environ. Res. Public Health 2021, 18, 12836. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, M.; Chen, R.; Xu, Z.; Yang, X.; Gu, H.; Zhang, L.; Fu, C.; Zhang, J.; Wu, Y. Gegen Qinlian standard decoction alleviated irinotecan-induced diarrhea via PI3K/AKT/NF-κB axis by network pharmacology prediction and experimental validation combination. Chin. Med. 2023, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Song, Y.; Li, G.; Xiao, P.; Liu, Y.; Xue, Y.; Cao, Q.; Tu, X.; Pan, T.; Jiang, Z.; et al. Fbxw7 increases CCL2/7 in CX3CR1hi macrophages to promote intestinal inflammation. J. Clin. Invest. 2019, 129, 3877–3893. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Calder, P.C. Effects of Citrus Fruit Juices and Their Bioactive Components on Inflammation and Immunity: A Narrative Review. Front. Immunol. 2021, 12, 712608. [Google Scholar] [CrossRef] [PubMed]
- Pierrakos, C.; Velissaris, D.; Bisdorff, M.; Marshall, J.C.; Vincent, J.A.-O. Biomarkers of sepsis: Time for a reappraisal. Crit. Care 2020, 24, 287. [Google Scholar] [CrossRef]
- Liu, X.; Fan, Y.; Du, L.; Mei, Z.; Fu, Y. In Silico and In Vivo Studies on the Mechanisms of Chinese Medicine Formula (Gegen Qinlian Decoction) in the Treatment of Ulcerative Colitis. Front. Pharmacol. 2021, 12, 665102. [Google Scholar] [CrossRef]
- Zhao, X.; Kang, B.; Lu, C.; Liu, S.; Wang, H.; Yang, X.; Chen, Y.; Jiang, B.; Zhang, J.; Lu, Y.; et al. Evaluation of p38 MAPK pathway as a molecular signature in ulcerative colitis. J. Proteome Res. 2011, 10, 2216–2225. [Google Scholar] [CrossRef]
- Wei, Y.; Fan, Y.; Huang, S.; Lv, J.; Zhang, Y.; Hao, Z. Baizhu shaoyao decoction restores the intestinal barrier and brain-gut axis balance to alleviate diarrhea-predominant irritable bowel syndrome via FoxO1/FoxO3a. Phytomedicine 2024, 122, 155163. [Google Scholar] [CrossRef]
- Huang, M.Y.; Pan, H.; Liang, Y.D.; Wei, H.X.; Xu, L.H.; Zha, Q.B.; He, X.H.; Ouyang, D.Y. Chemotherapeutic agent CPT-11 eliminates peritoneal resident macrophages by inducing apoptosis. Apoptosis 2016, 21, 130–142. [Google Scholar] [CrossRef]
- Hinz, N.; Jücker, M.A.-O. Distinct functions of AKT isoforms in breast cancer: A comprehensive review. Cell Commun. Signal 2019, 17, 154. [Google Scholar] [CrossRef]
- Li, Y.Z.; Wang, Y.Y.; Huang, L.; Zhao, Y.Y.; Chen, L.H.; Zhang, C. Annexin A protein family in atherosclerosis. Clin. Chim. Acta 2022, 531, 406–417. [Google Scholar] [CrossRef]
- Lei, K.; Wei, Q.; Cheng, Y.; Wang, Z.; Wu, H.; Zhao, F.; Ding, W.; Shi, F. OONO-/MMP2/MMP9 pathway-mediated apoptosis of porcine granulosa cells is associated with DNA damage. Reproduction 2023, 165, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Li, Y.; Yang, B.; Long, F.; Yang, Z.; Tang, D. Exploring the mechanism of action of Yiyi Fuzi Baijiang powder in colorectal cancer based on network pharmacology and molecular docking studies. Biotechnol. Genet. Eng. Rev. 2023, 1–21. [Google Scholar] [CrossRef]
- Gobé, G.; Zhang, X.J.; Cuttle, L.; Pat, B.; Willgoss, D.; Hancock, J.; Barnard, R.; Endre, R.B. Bcl-2 genes and growth factors in the pathology of ischaemic acute renal failure. Immunol. Cell Biol. 1999, 77, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Yang, J. PPAR-γ silencing inhibits the apoptosis of A549 cells by upregulating Bcl-2. Chin. J. Lung Cancer 2013, 16, 125–130. [Google Scholar]
- Muhammad, T.A.-O.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a Citrus Flavonoid, Attenuates LPS-Induced Neuroinflammation, Apoptosis and Memory Impairments by Modulating TLR4/NF-κB Signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef]
- Ashkenazi, A.; Fairbrother, W.J.; Leverson, J.D.; Souers, A.J. From basic apoptosis discoveries to advanced selective BCL-2 family inhibitors. Nat. Rev. Drug Discov. 2017, 16, 273–284. [Google Scholar] [CrossRef]
- Wang, J.; Feng, W.; Zhang, S.; Chen, L.; Tang, F.; Sheng, Y.; Ao, H.; Peng, C. Gut microbial modulation in the treatment of chemotherapy-induced diarrhea with Shenzhu Capsule. BMC Complement. Altern. Med. 2019, 19, 126. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Y.; Wang, Q.; Wang, Y.; Fan, Y.; Shang, E.; Jiang, S.; Duan, J. 6-Gingerol ameliorates ulcerative colitis by inhibiting ferroptosis based on the integrative analysis of plasma metabolomics and network pharmacology. Food Funct. 2024, 15, 6054–6067. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef] [PubMed]
- Rebhan, M.; Chalifa-Caspi, V.; Prilusky, J.; Lancet, D. GeneCards: Integrating information about genes, proteins and diseases. Trends Genet. 1997, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Amberger, J.S.; Bocchini, C.A.; Scott, A.F.; Hamosh, A. OMIM.org: Leveraging knowledge across phenotype-gene relationships. Nucleic Acids Res. 2019, 47, D1038–D1043. [Google Scholar] [CrossRef] [PubMed]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, C.; Dong, L. Exploring the Mechanism of White Peony in the Treatment of Lupus Nephritis Based on Network Pharmacology and Molecular Docking. Arch. Esp. Urol. 2023, 76, 123–131. [Google Scholar] [CrossRef]
- Tang, D.; Chen, M.; Huang, X.; Zhang, G.; Zeng, L.; Zhang, G.; Wu, S.; Wang, Y. SRplot: A free online platform for data visualization and graphing. PLoS ONE 2023, 18, e0294236. [Google Scholar] [CrossRef]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Wang, R.; Fang, X.; Lu, Y.; Wang, S. The PDBbind database: Collection of binding affinities for protein-ligand complexes with known three-dimensional structures. J. Med. Chem. 2017, 48, 3126–3139. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Grove, L.E.; Hall, D.R.; Bohnuud, T.; Mottarella, S.E.; Luo, L.; Xia, B.; Beglov, D.; Vajda, S. The FTMap family of web servers for determining and characterizing ligand-binding hot spots of proteins. Nat. Protoc. 2015, 10, 733–755. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, X.; Xu, R.; Xiong, P.; Liu, J.; Zhou, Z.; Shen, T.; Zhang, X. Exploring the Effects and Potential Mechanisms of Hesperidin for the Treatment of CPT-11-Induced Diarrhea: Network Pharmacology, Molecular Docking, and Experimental Validation. Int. J. Mol. Sci. 2024, 25, 9309. https://doi.org/10.3390/ijms25179309
Shu X, Xu R, Xiong P, Liu J, Zhou Z, Shen T, Zhang X. Exploring the Effects and Potential Mechanisms of Hesperidin for the Treatment of CPT-11-Induced Diarrhea: Network Pharmacology, Molecular Docking, and Experimental Validation. International Journal of Molecular Sciences. 2024; 25(17):9309. https://doi.org/10.3390/ijms25179309
Chicago/Turabian StyleShu, Xinyao, Ruitong Xu, Peiyu Xiong, Junyu Liu, Zubing Zhou, Tao Shen, and Xiaobo Zhang. 2024. "Exploring the Effects and Potential Mechanisms of Hesperidin for the Treatment of CPT-11-Induced Diarrhea: Network Pharmacology, Molecular Docking, and Experimental Validation" International Journal of Molecular Sciences 25, no. 17: 9309. https://doi.org/10.3390/ijms25179309
APA StyleShu, X., Xu, R., Xiong, P., Liu, J., Zhou, Z., Shen, T., & Zhang, X. (2024). Exploring the Effects and Potential Mechanisms of Hesperidin for the Treatment of CPT-11-Induced Diarrhea: Network Pharmacology, Molecular Docking, and Experimental Validation. International Journal of Molecular Sciences, 25(17), 9309. https://doi.org/10.3390/ijms25179309