
Telomere Length, HLA, and Longevity—Results from a Multicenter Study
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Distribution of HLA Class I and II Alleles in the Studied Populations
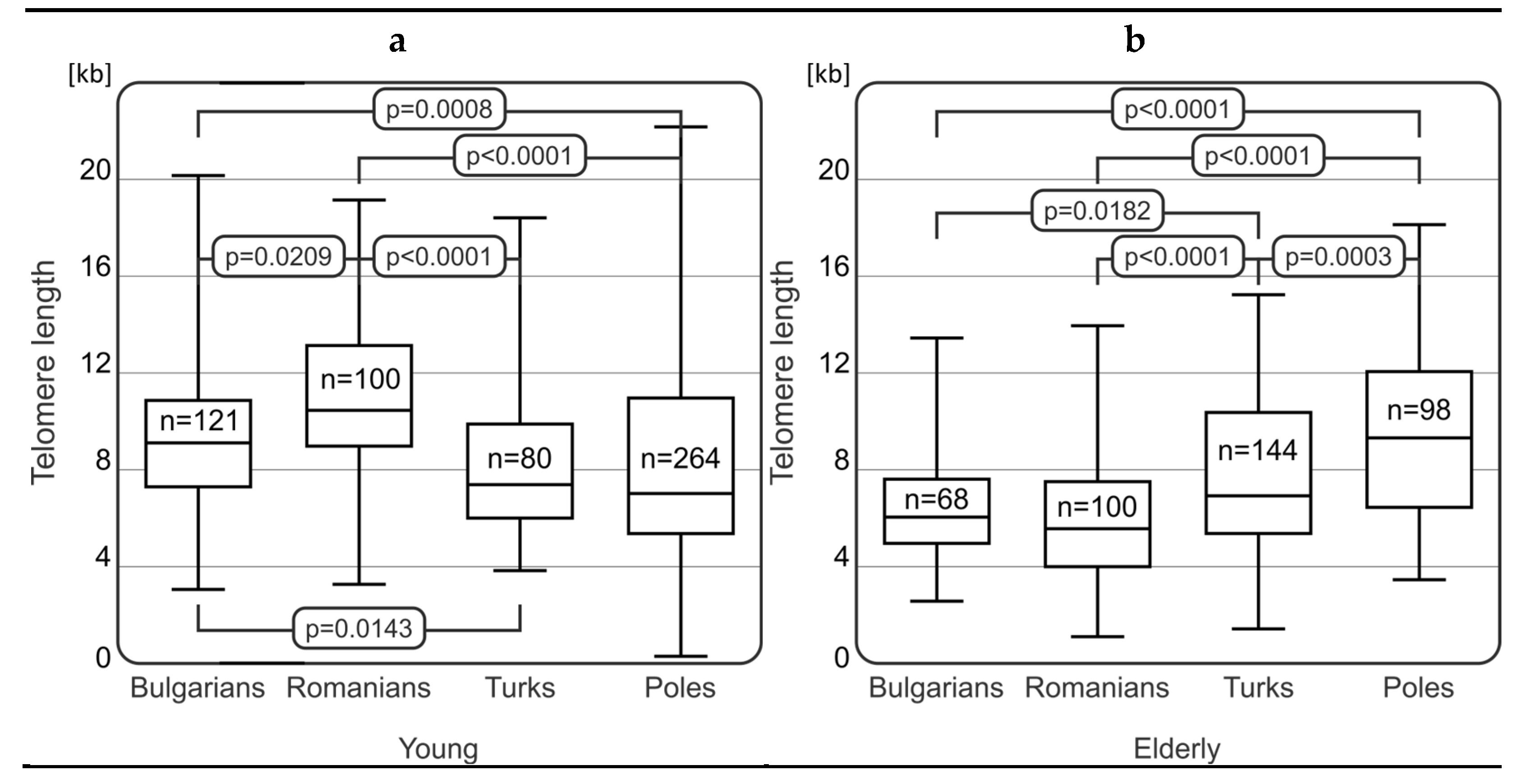
2.2. Comparison of Telomere Length among Elderly and Young Representatives in the Studied Populations
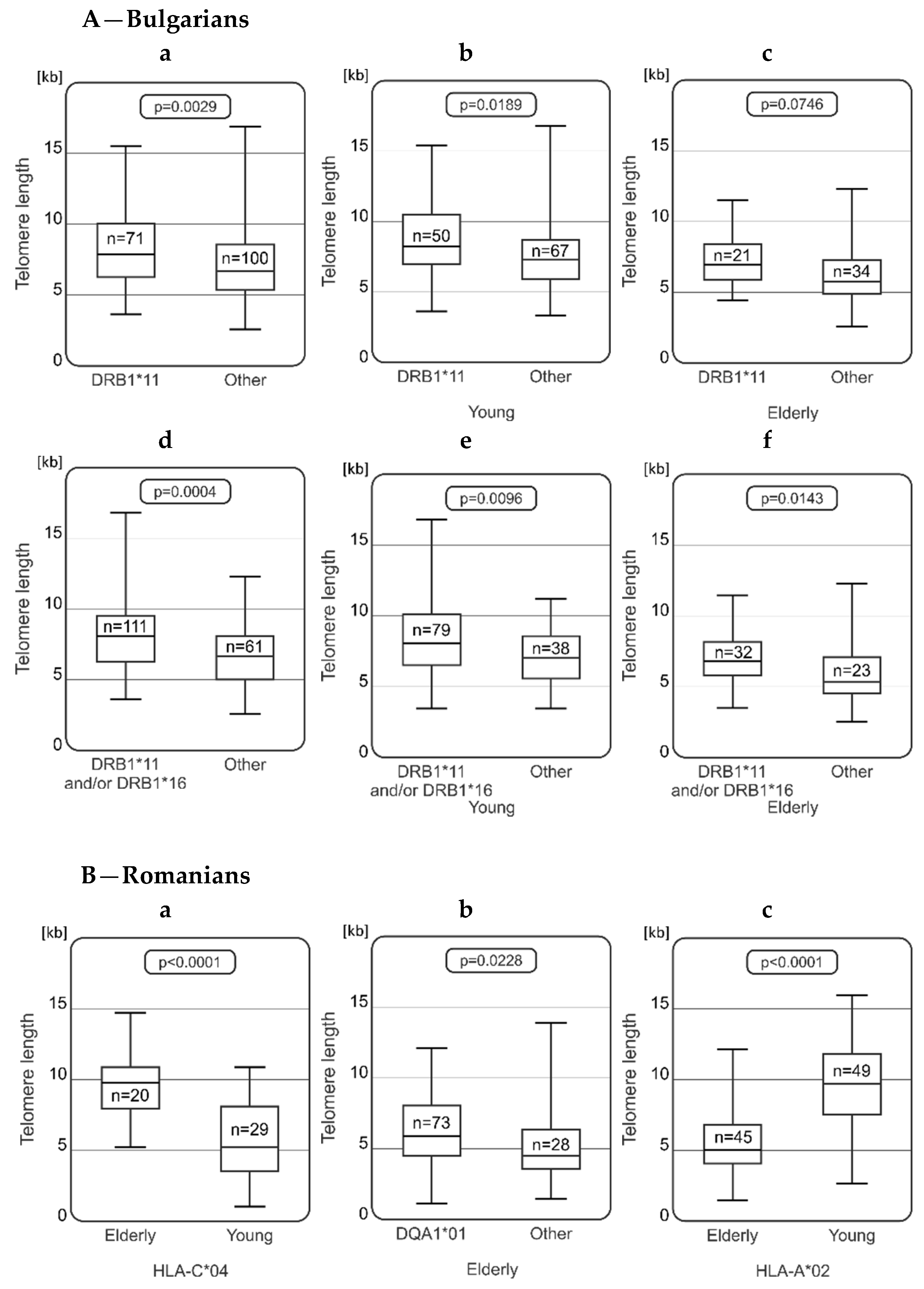
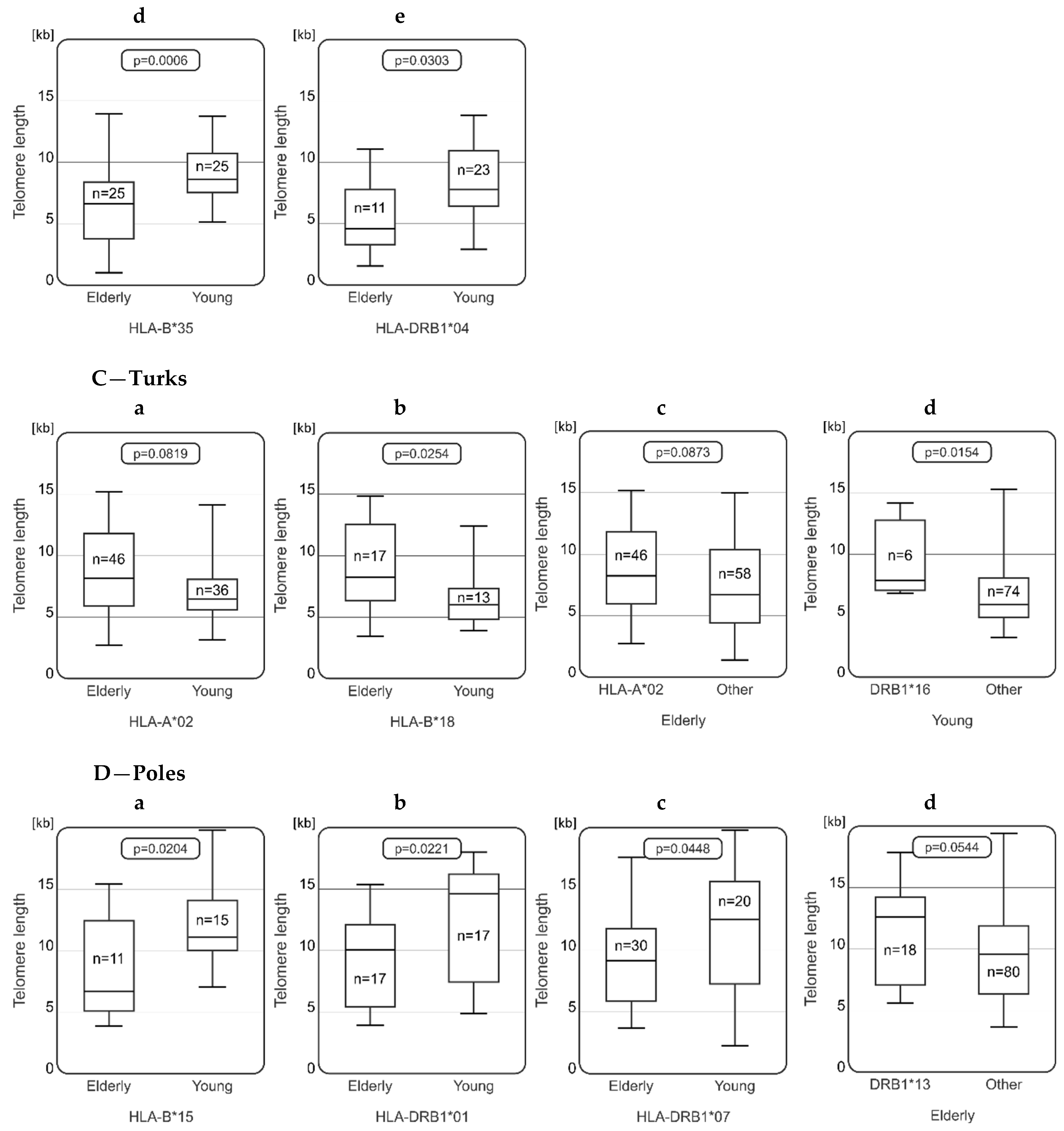
2.3. Relationships between Telomere Length and HLA Genotype

{kind=link}
{kind=link}
{kind=link}
| Haplotype | HF Young (n = 122) | HF Elderly (n = 62) | OR | CI.L | CI.U | p-Value |
|---|---|---|---|---|---|---|
| A*02~B*35~DRB1*16 | 0 | 0.0323 | 18.26 | 0.98 | 341.97 | 0.005 |
| A*02~B*49~DRB1*14 | 0 | 0.0242 | 14.09 | 0.72 | 274.91 | 0.015 |
| A*02~B*51~DRB1*10 | 0 | 0.0161 | 9.98 | 0.48 | 209.50 | 0.047 |
| A*02~B*57~DRB1*07 | 0 | 0.0161 | 9.98 | 0.48 | 209.50 | 0.047 |
| A*24~B*40~DRB1*03 | 0 | 0.0161 | 9.98 | 0.48 | 209.50 | 0.047 |
| A*01~DRB1*11 | 0.0451 | 0.1048 | 2.48 | 0.99 | 6.31 | 0.030 |
| A*02~DRB1*10 | 0.0041 | 0.0323 | 8.10 | 0.90 | 73.27 | 0.030 |
| B*35~DRB1*07 | 0.0041 | 0.0323 | 8.10 | 0.90 | 73.27 | 0.030 |
| C*08~DRB1*01 | 0.0410 | 0 | 0.09 | 0.01 | 1.54 | 0.020 |
| DRB1*04~DQB1*04 | 0 | 0.0242 | 14.09 | 0.72 | 274.91 | 0.015 |
| DRB1*07~DQB1*03 | 0.0041 | 0.0323 | 8.10 | 0.90 | 73.27 | 0.030 |
| Haplotype | HF Young (n = 100) | HF Elderly (n = 100) | OR | CI.L | CI.U | p-Value |
|---|---|---|---|---|---|---|
| A*02~DRB1*12 | 0.03 | 0 | 0.07 | 0.00 | 1.33 | 0.030 |
| A*03~DRB1*01 | 0.005 | 0.04 | 8.29 | 1.03 | 66.93 | 0.040 |
| A*01~B*08 | 0.02 | 0.07 | 3.69 | 1.13 | 15.62 | 0.016 |
| A*02~B*35 | 0.075 | 0.03 | 0.38 | 0.12 | 1.07 | 0.040 |
| A*02~C*07 | 0.14 | 0.07 | 0.46 | 0.22 | 0.95 | 0.020 |
| A*68~C*07 | 0 | 0.04 | 17.71 | 1.01 | 308.89 | 0.007 |
| DRB1*04~DQB1*03 | 0.125 | 0.065 | 0.49 | 0.22 | 1.03 | 0.040 |
| DRB1*04~DQA1*03~DQB1*03 | 0.125 | 0.055 | 0.41 | 0.18 | 0.89 | 0.010 |
| DRB1*14~DQA1*01~DQB1*05 | 0.025 | 0.065 | 2.71 | 0.88 | 9.88 | 0.054 |
| A*01~B*08~DRB1*03 | 0.015 | 0.055 | 3.82 | 0.99 | 21.6 | 0.030 |
| Haplotype | HF Young (n = 158) | HF Elderly (n = 122) | OR | CI.L | CI.U | p-Value |
|---|---|---|---|---|---|---|
| A*02~C*05 | 0.0095 | 0.0410 | 4.46 | 1.13 | 25.42 | 0.010 |
| B*15~C*03 | 0.0253 | 0 | 0.07 | 0.00 | 1.29 | 0.010 |
| A*01~DRB1*11 | 0.0253 | 0.0041 | 0.16 | 0.02 | 1.28 | 0.048 |
| A*02~DRB1*03 | 0.0158 | 0.0492 | 3.22 | 1.04 | 11.8 | 0.020 |
| A*26~DRB1*04 | 0.0190 | 0 | 0.10 | 0.01 | 1.74 | 0.040 |
| A*26~DRB1*07 | 0 | 0.0164 | 11.84 | 0.63 | 221.06 | 0.040 |
| A*03~DRB1*03 | 0 | 0.0164 | 11.84 | 0.63 | 221.06 | 0.040 |
| B*07~DRB1*15 | 0.0285 | 0.0041 | 0.14 | 0.02 | 1.12 | 0.030 |
| B*51~DRB1*15 | 0.0285 | 0.0041 | 0.14 | 0.02 | 1.12 | 0.030 |
| B*44~DRB1*15 | 0.0190 | 0 | 0.10 | 0.005 | 1.74 | 0.040 |
| C*04~DRB1*04 | 0.0411 | 0.0082 | 0.19 | 0.02 | 0.87 | 0.020 |
| C*04~DRB1*11 | 0.0222 | 0.0574 | 2.69 | 0.99 | 7.98 | 0.030 |
| DRB1*10~DQB1*05 | 0.0380 | 0.0082 | 0.21 | 0.05 | 0.94 | 0.025 |
| DRB1*15~DQB1*05 | 0.0285 | 0.0000 | 0.07 | 0.004 | 1.14 | 0.008 |
| A*03~B*35~DRB1*03 | 0 | 0.0164 | 11.84 | 0.63 | 221.06 | 0.040 |
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Quantification of Telomere Length
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef] [PubMed]
- Cooke, H.J.; Smith, B.A. Variability at the telomeres of the human X/Y pseudoautosomal region. Cold Spring Harb. Symp. Quant. Biol. 1986, 51 Pt 1, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, R.C.; Vaziri, H.; Patterson, C.; Goldstein, S.; Younglai, E.V.; Futcher, A.B.; Greider, C.W.; Harley, C.B. Telomere length predicts replicative capacity of human fibroblasts. Proc. Natl. Acad. Sci. USA 1992, 89, 10114–10118. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.B.; Vaziri, H.; Counter, C.M.; Allsopp, R.C. The telomere hypothesis of cellular aging. Exp. Gerontol. 1992, 27, 375–382. [Google Scholar] [CrossRef]
- Njajou, O.T.; Cawthon, R.M.; Damcott, C.M.; Wu, S.H.; Ott, S.; Garant, M.J.; Blackburn, E.H.; Mitchell, B.D.; Shuldiner, A.R.; Hsueh, W.C. Telomere length is paternally inherited and is associated with parental lifespan. Proc. Natl. Acad. Sci. USA 2007, 104, 12135–12139. [Google Scholar] [CrossRef]
- Vasa-Nicotera, M.; Brouilette, S.; Mangino, M.; Thompson, J.R.; Braund, P.; Clemitson, J.R.; Mason, A.; Bodycote, C.L.; Raleigh, S.M.; Louis, E.; et al. Mapping of a major locus that determines telomere length in humans. Am. J. Hum. Genet. 2005, 76, 147–151, Erratum in Am. J. Hum. Genet. 2005, 76, 373. [Google Scholar] [CrossRef]
- Bekaert, S.; De Meyer, T.; Van Oostveldt, P. Telomere attrition as ageing biomarker. Anticancer. Res. 2005, 25, 3011–3021. [Google Scholar]
- Lulkiewicz, M.; Bajsert, J.; Kopczynski, P.; Barczak, W.; Rubis, B. Telomere length: How the length makes a difference. Mol. Biol. Rep. 2020, 47, 7181–7188. [Google Scholar] [CrossRef]
- Honig, L.S.; Kang, M.S.; Cheng, R.; Eckfeldt, J.H.; Thyagarajan, B.; Leiendecker-Foster, C.; Province, M.A.; Sanders, J.L.; Perls, T.; Christensen, K.; et al. Heritability of telomere length in a study of long-lived families. Neurobiol. Aging 2015, 36, 2785–2790. [Google Scholar] [CrossRef] [PubMed]
- Montiel Rojas, D.; Nilsson, A.; Ponsot, E.; Brummer, R.J.; Fairweather-Tait, S.; Jennings, A.; de Groot, L.C.P.G.M.; Berendsen, A.; Pietruszka, B.; Madej, D.; et al. Short Telomere Length Is Related to Limitations in Physical Function in Elderly European Adults. Front. Physiol. 2018, 9, 1110. [Google Scholar] [CrossRef]
- Zierer, J.; Kastenmüller, G.; Suhre, K.; Gieger, C.; Codd, V.; Tsai, P.C.; Bell, J.; Peters, A.; Strauch, K.; Schulz, H.; et al. Metabolomics profiling reveals novel markers for leukocyte telomere length. Aging 2016, 8, 77–94. [Google Scholar] [CrossRef]
- Kimura, M.; Barbieri, M.; Gardner, J.P.; Skurnick, J.; Cao, X.; van Riel, N.; Rizzo, M.R.; Paoliso, G.; Aviv, A. Leukocytes of exceptionally old persons display ultra-short telomeres. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R2210–R2217. [Google Scholar] [CrossRef] [PubMed]
- Gardner, M.; Bann, D.; Wiley, L.; Cooper, R.; Hardy, R.; Nitsch, D.; Martin-Ruiz, C.; Shiels, P.; Sayer, A.A.; Barbieri, M.; et al. Gender and telomere length: Systematic review and meta-analysis. Exp. Gerontol. 2014, 51, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.E.; Hunt, S.C.; Stone, R.C.; Horvath, K.; Herbig, U.; Ranciaro, A.; Hirbo, J.; Beggs, W.; Reiner, A.P.; Wilson, J.G.; et al. Shorter telomere length in Europeans than in Africans due to polygenetic adaptation. Hum. Mol. Genet. 2016, 25, 2324–2330. [Google Scholar] [CrossRef] [PubMed]
- Bosquet Enlow, M.; Bollati, V.; Sideridis, G.; Flom, J.D.; Hoxha, M.; Hacker, M.R.; Wright, R.J. Sex differences in effects of maternal risk and protective factors in childhood and pregnancy on newborn telomere length. Psychoneuroendocrinology 2018, 95, 74–85. [Google Scholar] [CrossRef]
- Candore, G.; Balistreri, C.R.; Listì, F.; Grimaldi, M.P.; Vasto, S.; Colonna-Romano, G.; Franceschi, C.; Lio, D.; Caselli, G.; Caruso, C. Immunogenetics, gender, and longevity. Ann. N. Y. Acad. Sci. 2006, 1089, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Wick, G.; Huber, L.A.; Xu, Q.B.; Jarosch, E.; Schönitzer, D.; Jürgens, G. The decline of the immune response during aging: The role of an altered lipid metabolism. Ann. N. Y. Acad. Sci. 1991, 621, 277–290. [Google Scholar] [CrossRef]
- Lindmeier, C. “Ageing Well” Must Be a Global Priority. Available online: https://www.who.int/news/item/06-11-2014--ageing-well-must-be-a-global-priority (accessed on 22 July 2024).
- OECD. Trends in life expectancy. In Health at a Glance 2021; OECD Indicators; OECD Publishing: Paris, France, 2021; Available online: https://www.oecd-ilibrary.org/social-issues-migration-health/health-at-a-glance-2021_e0d509f9-en (accessed on 22 July 2024).
- United Nations Department of Economic and Social Affairs, Population Division. World Population Ageing 2020 Highlights: Living Arrangements of Older Persons (ST/ESA/SER.A/451). 2020. Available online: https://www.un.org/development/desa/pd/sites/www.un.org.development.desa.pd/files/undesa_pd-2020_world_population_ageing_highlights.pdf (accessed on 22 July 2024).
- Zhu, H.; Belcher, M.; van der Harst, P. Healthy aging and disease: Role for telomere biology? Clin. Sci. 2011, 120, 427–440. [Google Scholar] [CrossRef]
- Kim, S.; Bi, X.; Czarny-Ratajczak, M.; Dai, J.; Welsh, D.A.; Myers, L.; Welsch, M.A.; Cherry, K.E.; Arnold, J.; Poon, L.W.; et al. Telomere maintenance genes SIRT1 and XRCC6 impact age-related decline in telomere length but only SIRT1 is associated with human longevity. Biogerontology 2012, 13, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Atzmon, G.; Cho, M.; Cawthon, R.M.; Budagov, T.; Katz, M.; Yang, X.; Siegel, G.; Bergman, A.; Huffman, D.M.; Schechter, C.B.; et al. Evolution in health and medicine Sackler colloquium: Genetic variation in human telomerase is associated with telomere length in Ashkenazi centenarians. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. 1), 1710–1717. [Google Scholar] [CrossRef]
- Halaschek-Wiener, J.; Vulto, I.; Fornika, D.; Collins, J.; Connors, J.M.; Le, N.D.; Lansdorp, P.M.; Brooks-Wilson, A. Reduced telomere length variation in healthy oldest old. Mech. Ageing Dev. 2008, 129, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Arai, Y.; Martin-Ruiz, C.M.; Takayama, M.; Abe, Y.; Takebayashi, T.; Koyasu, S.; Suematsu, M.; Hirose, N.; von Zglinicki, T. Inflammation, But Not Telomere Length, Predicts Successful Ageing at Extreme Old Age: A Longitudinal Study of Semi-supercentenarians. EBioMedicine 2015, 2, 1549–1558. [Google Scholar] [CrossRef] [PubMed]
- Terry, D.F.; Nolan, V.G.; Andersen, S.L.; Perls, T.T.; Cawthon, R. Association of longer telomeres with better health in centenarians. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 809–812. [Google Scholar] [CrossRef] [PubMed]
- Tedone, E.; Arosio, B.; Gussago, C.; Casati, M.; Ferri, E.; Ogliari, G.; Ronchetti, F.; Porta, A.; Massariello, F.; Nicolini, P.; et al. Leukocyte telomere length and prevalence of age-related diseases in semisupercentenarians, centenarians and centenarians’ offspring. Exp. Gerontol. 2014, 58, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Tedone, E.; Huang, E.; O’Hara, R.; Batten, K.; Ludlow, A.T.; Lai, T.P.; Arosio, B.; Mari, D.; Wright, W.E.; Shay, J.W. Telomere length and telomerase activity in T cells are biomarkers of high-performing centenarians. Aging Cell. 2019, 18, e12859. [Google Scholar] [CrossRef]
- Caruso, C.; Candore, G.; Romano, G.C.; Lio, D.; Bonafè, M.; Valensin, S.; Franceschi, C. Immunogenetics of longevity. Is major histocompatibility complex polymorphism relevant to the control of human longevity? A review of literature data. Mech. Ageing Dev. 2001, 122, 445–462. [Google Scholar] [CrossRef]
- Pawelec, G.; Wagner, W.; Adibzadeh, M.; Engel, A. T cell immunosenescence in vitro and in vivo. Exp. Gerontol. 1999, 34, 419–429. [Google Scholar] [CrossRef]
- Arnaiz-Villena, A.; Reguera, R.; Parga-Lozano, C.; Abd-El-Fatah, S.; Monleon, L.; Barbolla, L.; Gomez-Prieto, P.; Martínez-Laso, J.; Silvera, C. HLA Genes in Afro-American Colombians (San Basilio de Palenque): The First Free Africans in America. Open Immunol. J. 2009, 2, 59–66. [Google Scholar] [CrossRef]
- Papasteriades, C.; Boki, K.; Pappa, H.; Aedonopoulos, S.; Papasteriadis, E.; Economidou, J. HLA phenotypes in healthy aged subjects. Gerontology 1997, 43, 176–181. [Google Scholar] [CrossRef]
- Ivanova, M.; Creary, L.E.; Al Hadra, B.; Lukanov, T.; Mazzocco, M.; Sacchi, N.; Ameen, R.; Al-Shemmari, S.; Moise, A.; Ursu, L.D.; et al. 17th IHIW component “Immunogenetics of Ageing”—New NGS data. Hum. Immunol. 2019, 80, 703–713. [Google Scholar] [CrossRef]
- Naumova, E.; Pawelec, G.; Ivanova, M.; Constantinescu, I.; Bogunia-Kubik, K.; Lange, A.; Qguz, F.; Carin, M. 14th International HLA and Immunogenetics Workshop: Report on the immunogenetics of aging. Tissue Antigens. 2007, 69 (Suppl. 1), 304–310. [Google Scholar] [CrossRef] [PubMed]
- Schreuder, G.M.; Hurley, C.K.; Marsh, S.G.; Lau, M.; Maiers, M.; Kollman, C.; Noreen, H.J. The HLA Dictionary 2001: A summary of HLA-A, -B, -C, -DRB1/3/4/5 and -DQB1 alleles and their association with serologically defined HLA-A, -B, -C, -DR and -DQ antigens. Eur. J. Immunogenet. 2001, 28, 565–596. [Google Scholar] [CrossRef] [PubMed]
- Schönland, S.O.; Lopez, C.; Widmann, T.; Zimmer, J.; Bryl, E.; Goronzy, J.J.; Weyand, C.M. Premature telomeric loss in rheumatoid arthritis is genetically determined and involves both myeloid and lymphoid cell lineages. Proc. Natl. Acad. Sci. USA 2003, 100, 13471–13476. [Google Scholar] [CrossRef]
- Akisaka, M.; Suzuki, M.; Inoko, H. Molecular genetic studies on DNA polymorphism of the HLA class II genes associated with human longevity. Tissue Antigens 1997, 50, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, R.; Hénon, N.; Lepage, V.; Charron, D.; Vicaut, E.; Schächter, F. HLA-DR alleles display sex-dependent effects on survival and discriminate between individual and familial longevity. Hum. Mol. Genet. 1998, 7, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kaul, R.; Kaul, A.; Khan, K. A comparative review of HLA associations with hepatitis B and C viral infections across global populations. World J. Gastroenterol. 2007, 13, 1770–1787. [Google Scholar] [CrossRef]
- Kunizheva, S.S.; Volobaev, V.P.; Plotnikova, M.Y.; Kupriyanova, D.A.; Kuznetsova, I.L.; Tyazhelova, T.V.; Rogaev, E.I. Current Trends and Approaches to the Search for Genetic Determinants of Aging and Longevity. Russ. J. Genet. 2022, 58, 1427–1443. [Google Scholar] [CrossRef]
- Belsky, D.W.; Caspi, A.; Houts, R.; Cohen, H.J.; Corcoran, D.L.; Danese, A.; Harrington, H.; Israel, S.; Levine, M.E.; Schaefer, J.D.; et al. Quantification of biological aging in young adults. Proc. Natl. Acad. Sci. USA 2015, 112, E4104–E4110. [Google Scholar] [CrossRef] [PubMed]
- Ligthart, G.J.; Corberand, J.X.; Fournier, C.; Galanaud, P.; Hijmans, W.; Kennes, B.; Müller-Hermelink, H.K.; Steinmann, G.G. Admission criteria for immunogerontological studies in man: The SENIEUR protocol. Mech. Ageing Dev. 1984, 28, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Dratwa, M.; Wysoczanska, B.; Turlej, E.; Anisiewicz, A.; Maciejewska, M.; Wietrzyk, J.; Bogunia-Kubik, K. Heterogeneity of telomerase reverse transcriptase mutation and expression, telomerase activity and telomere length across human cancer cell lines cultured in vitro. Exp. Cell Res. 2020, 396, 112298. [Google Scholar] [CrossRef] [PubMed]
- Pappas, D.J.; Marin, W.; Hollenbach, J.A.; Mack, S.J. Bridging ImmunoGenomic Data Analysis Workflow Gaps (BIGDAWG): An integrated case-control analysis pipeline. Hum. Immunol. 2016, 77, 283–287. [Google Scholar] [CrossRef] [PubMed]
| Elderly Individuals | Young Individuals | |
|---|---|---|
| Bulgaria | N = 68 | N = 121 |
| Male, n (%) | 24 (35%) | 43 (36%) |
| Female, n (%) | 44 (65%) | 78 (64%) |
| Mean age (years ± SD) | 71 ± 6.19 | 27 ± 4.03 |
| Romania | N = 100 | N = 100 |
| Male, n (%) | 49 (49%) | 55 (55%) |
| Female, n (%) | 51 (51%) | 45 (45%) |
| Mean age (years ± SD) | 71 ± 5.74 | 30 ± 3.40 |
| Turkey | N = 125 | N = 80 |
| Male, n (%) | 38 (30%) | 38 (48%) |
| Female, n (%) | 87 (70%) | 42 (52%) |
| Mean age (years ± SD) | 85 ± 6.99 | 37.79 ± 6.44 |
| Poland | N = 99 | N = 264 |
| Male, n (%) | 37 (37%) | 134 (51%) |
| Female, n (%) | 62 (63%) | 130 (49%) |
| Mean age (years ± SD) | 74 ± 6.19 | 34 ± 9.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dratwa-Kuzmin, M.; Hadra, B.A.; Oguz, F.; Ogret, Y.; Constantinescu, I.; Apostol, D.; Talangescu, A.; Constantinescu, A.-E.; Maruntelu, I.; Kościńska, K.; et al. Telomere Length, HLA, and Longevity—Results from a Multicenter Study. Int. J. Mol. Sci. 2024, 25, 9457. https://doi.org/10.3390/ijms25179457
Dratwa-Kuzmin M, Hadra BA, Oguz F, Ogret Y, Constantinescu I, Apostol D, Talangescu A, Constantinescu A-E, Maruntelu I, Kościńska K, et al. Telomere Length, HLA, and Longevity—Results from a Multicenter Study. International Journal of Molecular Sciences. 2024; 25(17):9457. https://doi.org/10.3390/ijms25179457
Chicago/Turabian StyleDratwa-Kuzmin, Marta, Bushra Al Hadra, Fatma Oguz, Yeliz Ogret, Ileana Constantinescu, Dimitri Apostol, Adriana Talangescu, Alexandra-Elena Constantinescu, Ion Maruntelu, Katarzyna Kościńska, and et al. 2024. "Telomere Length, HLA, and Longevity—Results from a Multicenter Study" International Journal of Molecular Sciences 25, no. 17: 9457. https://doi.org/10.3390/ijms25179457