Abstract
Mycorrhizal fungi, a category of fungi that form symbiotic relationships with plant roots, can participate in the induction of plant disease resistance by secreting phosphatase enzymes. While extensive research exists on the mechanisms by which mycorrhizal fungi induce resistance, the specific contributions of phosphatases to these processes require further elucidation. This article reviews the spectrum of mycorrhizal fungi-induced resistance mechanisms and synthesizes a current understanding of how phosphatases mediate these effects, such as the induction of defense structures in plants, the negative regulation of plant immune responses, and the limitation of pathogen invasion and spread. It explores the role of phosphatases in the resistance induced by mycorrhizal fungi and provides prospective future research directions in this field.
1. Introduction
Plants in the natural world are continually under the threat of pathogens, which can cause diseases that affect crop yield and quality. To counter these biotic stresses, plants have evolved a complex immune system.
The German scholar Frank discovered that the roots of some plants form symbiotic relationships with fungi. These fungi, while obtaining carbohydrates from the plants, help the plants absorb mineral elements and water from the soil. This symbiotic relationship is known as mycorrhiza [1]. Based on morphological characteristics, mycorrhizae can be mainly divided into ectomycorrhizae and endomycorrhizae. Among them, arbuscular mycorrhizae represent a special form of endomycorrhizae which are formed by Glomeromycetes fungi. These fungi play a vital role in enhancing the survival rate of plant seedlings [2], promoting plant growth [3], bioremediation [4], nutrient absorption [5,6], and increasing plant disease resistance [7,8] (Figure 1).
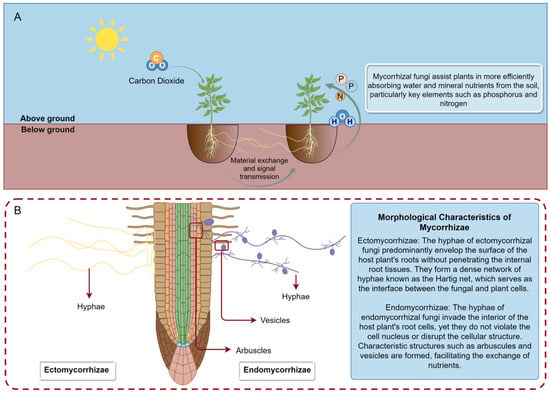
Figure 1.
The functions of mycorrhizal fungi. (A) In the mycorrhizal symbiotic system, the organic carbon produced by plants through photosynthesis is primarily in the form of carbohydrates, such as glucose, fructose, and sucrose, which are transferred to mycorrhizal fungi via plant roots. In return, mycorrhizal fungi assist plants in more effectively absorbing water and mineral nutrients from the soil, particularly key elements like phosphorus and nitrogen. Additionally, mycorrhizal fungi form an extensive underground network through their hyphae, connecting different plant root systems to create a shared mycorrhizal network. Through this network, mycorrhizal fungi can engage in material exchange and signal transmission. (B) Comparative visualization of ectomycorrhizal and endomycorrhizal mycelia: ectomycorrhizal hyphae depicted on the left, with endomycorrhizal hyphae illustrated on the right.
Phosphorus (P), as an essential nutrient element for plant growth, is mainly absorbed and utilized by plants in the form of inorganic phosphates (e.g., HPO42−, H2PO4−) [9]. It also participates in various physiological and biochemical metabolic processes within the plant body, including energy transfer, signal transduction, redox reactions, and photosynthesis [10]. Improving the plant utilization of organic and insoluble inorganic phosphorus from the soil has been a longstanding agricultural challenge. Research indicates that, in addition to modulating the morphology and physiological characteristics of their root systems, plants have evolved a variety of mechanisms to cope with phosphorus deficiency, one of which is the formation of mycorrhizal symbioses to enhance phosphorus uptake from the soil [11]. Mycorrhizal fungi form a physical connection with plant roots through their extramatrical mycelium and extend in the soil to increase the plant’s absorption area for water and nutrients. Among them, the phosphatases secreted by mycorrhizal fungi can transform organic phosphorus in the soil into inorganic phosphorus that can be directly absorbed by plants, thereby improving the nutritional status of the plants. In addition, phosphatases may also enhance the plant’s defense against pathogens [12] by regulating the levels of plant hormones, activating plant defense-related genes [13], and enhancing systemic acquired resistance (SAR) [14,15,16,17].
This review aims to provide a comprehensive analysis of the biological functions of phosphatases in mycorrhizal fungi and their key roles in the plant disease defense mechanism, in order to deeply understand their molecular mechanisms and regulatory networks. By discussing the diversity and specificity of these key enzyme classes, this study aims to provide a theoretical basis and practical guidance for the development of new plant disease management strategies.
2. Mycorrhizal Fungi-Induced Host Plant Resistance to Disease
2.1. The Array of Plant Pathologies That Mycorrhizal Fungi Can Suppress
Plant pathogens encompass a vast array of categories, including Viruses, Prokaryotes, Fungi, Oomycota (such as Albuginales and Peronosporaceae), Nematodes, parasitic plants (like Cuscuta spp.) [18,19,20] (Figure 2).

Figure 2.
Categories of plant pathogens.
It is known that there are more than 30,000 species of Fungi that cause plant diseases, accounting for approximately 70% to 80% of all plant diseases. They are distributed across multiple phyla, classes, orders, families, and genera, such as Ascochyta, Didymella, Epicoccum, and so on. Oomycota belong to the Stramenopila group; although morphologically similar to fungi, they are biologically distinct in terms of classification [21,22]. They are eukaryotic organisms but not classified as fungi and can cause diseases such as late blight caused by Phytophthora spp. [23]. Prokaryotes, including bacterial pathogens, can induce a variety of plant diseases, exemplified by Pseudomonas syringae [24].
Ahmed et al. [25] found that the interaction between mycorrhizal fungi and plants can affect plant–nematode interactions, thereby enhancing the plant’s resistance to nematodes, highlighting the potential of mycorrhizal fungi in improving plant resistance to diseases. Mycorrhizal fungi-induced disease resistance spans a variety of pathogen-induced diseases (Table 1). Mycorrhization by arbuscular mycorrhizal fungi (AMF) can significantly enhance the growth parameters, photosynthetic pigments, and flavonoid content in tomato plants, while reducing the severity and incidence of Tomato mosaic virus (ToMV) infection [26].

Table 1.
Types of plant diseases inhibited by mycorrhizal fungi.
Table 1.
Types of plant diseases inhibited by mycorrhizal fungi.
| Pathogen | Disease Name | Pathogen | Host Plant | Mycorrhizal Fungal Names | Mycorrhizal Types | Mechanism of Disease Resistance |
|---|---|---|---|---|---|---|
| Oomycota | Late blight disease | Phytophthora infestans | Solanum tuberosum | Glomus sp. | Arbuscular mycorrhizae | Inducing systemic acquired resistance by activating plant defense genes (such as PR1 and PR2), reducing the leaf infection index, and enhancing resistance to late blight [23] |
| Fungi | Fusarium wilt | Fusarium oxysporum | Solanum lycopersicum | Glomus mosseae | Arbuscular mycorrhizae | Combating soil-borne pathogens in tomatoes, providing bioprotection effects [27] |
| Fusarium oxysporum | Salvia miltiorrhiza | Glomus versiforme | Arbuscular mycorrhizae | Mycorrrhizal colonization enhances the host plant’s resistance to fungal pathogens by strengthening photosynthesis, root structure, and inducing the expression of defense enzymes and defense-related genes to combat infection [28] | ||
| Fusarium oxysporum | Musa acuminate | Rhizophagus irregularis | Arbuscular mycorrhizae | Promoting the growth of banana plants, inducing the expression of defense-related genes, aiding in the suppression of wilt disease [29] | ||
| Fusarium oxysporum | Citrullus lanatus | Funneliformis mosseae or Glomus versiforme | Arbuscular mycorrhizae | By inducing the root exudation of phthalic esters, altering soil enzyme activity and bacterial community composition, wilt disease in watermelon is mitigated [30] | ||
| White Rot | Erysiphe alphitoides | Quercus robur | The commercially available mycorrhizal inoculant Ectovit®, which contains a variety of mycorrhizal fungi, was used | Multiple mycorrhizal fungi | Mycorrhizal fungi can significantly increase the levels of polyamines, soluble osmotic regulators (such as proline), and phenolic compounds in plant leaves, thereby enhancing the plant’s resistance to powdery mildew [31] | |
| Rust infection | Melampsora larici-populina | Populus trichocarpa × deltoides | Hebeloma mesophaeum | Ectomycorrhizae | By mitigating the reduction in the synthesis of phenolic compounds triggered by rust disease, the negative impact of rust on the host plant is compensated for [32] | |
| Verticillium wilt | Rhizoctonia solani | Cucumis sativus | Glomus mosseae and Glomus clarum | Arbuscular mycorrhizae | Mycorrhizal fungi significantly reduced disease severity and increased plant biomass, potentially through mechanisms such as improving nutritional status, reducing direct competition with pathogens, and inducing plant immunity [33] | |
| Verticillium dahliae | Gossypium hirsutum | Glomus etunicatum, Glomus intraradices, Glomus versiforme | Arbuscular mycorrhizae | Mycorrhizal fungi exhibit competitive interactions with the pathogen V. dahliae, which can alleviate the disease effects of V. dahliae on cotton and enhance the plant’s resistance to the disease [34] | ||
| Verticillium dahliae | Gossypium hirsutum | Rhizophagus irregularis | Arbuscular mycorrhizae | By inducing the expression of plant resistance-related genes and the potential release of volatile compounds by mycorrhizal fungi symbionts, which directly affect the growth of pathogenic fungi [35] | ||
| Early blight disease | Alternaria solani | Solanum lycopersicum | Glomus intraradices | Arbuscular mycorrhizae | Reducing the susceptibility of tomatoes to A. solani, diminishing disease symptoms, is akin to the induction of systemic resistance (ISR) [36] | |
| Damping off | Rhizoctonia solani | Pinus tabulaeformis | Suillus laricinus, S. tomentosus, Amanita vaginata, Gomphidius viscidus | Ectomycorrhizae | Inhibiting the growth of pathogens by producing hydrolytic enzymes (chitinase, β-1,3-glucanase, and β-glucosidase) that participate in the parasitic action on the fungi, altering the morphology of the pathogen [15] | |
| Black pod disease | Phytophthora megakarya | Theobroma cacao | Gigaspora margarita and Acaulospora tuberculata | Arbuscular mycorrhizae | Promoting the growth of cocoa, enhancing resistance to black pod disease, and increasing plant growth parameters, such as height, root, and stem weight, as well as phosphorus uptake [37] | |
| Black foot disease | Cylindrocarpon macrodidymum | Vitis Rupestris | Glomus intraradices | Arbuscular mycorrhizae | Reducing the susceptibility of grapevine roots to black foot disease, enhancing the plant’s resistance to abiotic or biotic stresses, and mitigating the severity of the disease [38] | |
| Decline syndrome | Phytophthora cinnamomi | Quercus ilex | Tomentella spp., Russula spp. | Ectomycorrhizae | Affecting the vitality of oak roots and the abundance of mycorrhizal fungi, the interplay of soil properties, topography, and root infection by P. cinnamomi influences the abundance of mycorrhizal fungi [7] | |
| Prokaryotes | Bacterial wilt disease | Ralstonia solanacearum | Solanum lycopersicum | Glomus mosseae, Scutellospora sp., Gigaspora margarita | Arbuscular mycorrhizae | The integration of Glomus mosseae with the pathogen significantly enhanced the height and biomass of tomato plants, with no occurrence of disease symptoms [39] |
| Ralstonia solanacearum | Nicotiana tabacum | Glomus mosseae | Arbuscular mycorrhizae | The combined application of Trichoderma harzianum-amended bio-organic fertilizer and the mycorrhizal fungus Glomus mosseae reduced the abundance of the pathogen, and increased the activities of polyphenol oxidase (PPO), phenylalanine ammonia-lyase (PAL), and peroxidase (POD) in the plants, promoting plant growth [40] | ||
| Bacterial wilt | Ralstonia solanacearum | Solanum lycopersicum | Glomus mosseae | Arbuscular mycorrhizae | Tomatoes inoculated with Glomus mosseae, when combined with the use of organic fertilizers, have exhibited increased plant survival rates and yields [41] | |
| Ralstonia solanacearum | Solanum lycopersicum | Rhizophagus irregularis | Arbuscular mycorrhizae | By activating the plant’s defense mechanisms [42] | ||
| Nematodes | Root-knot nematode disease | Meloidogyne incognita | Solanum lycopersicum | Glomus mosseae | Arbuscular mycorrhizae | Inducing systemic acquired resistance reduced the number of root-knot nematodes, mitigating their damage to tomato root systems [43] |
| Root-lesion nematode disease | Pratylenchus penetrans | Solanum lycopersicum | Glomus mosseae | Arbuscular mycorrhizae | Inducing systemic resistance significantly reduced the number of root-lesion nematodes, decreased their reproduction rate, and lessened the damage to tomato root systems [43] | |
| False root-knot nematode | Nacobbus aberrans | Solanum lycopersicum | Glomus intraradices | Arbuscular mycorrhizae | Reducing root damage caused by nematodes (decreasing the number of root galls) and inhibiting nematode reproduction [44] | |
| Viruses | Tomato yellow leaf curl disease | Tomato yellow leaf curl Sardinia virus (TYLCSV) | Solanum tuberosum | Funneliformis mosseae | Arbuscular mycorrhizae | Mitigating the severity of viral symptoms, reducing the concentration of viral DNA in tomatoes, and enhancing the tolerance of tomatoes to TYLCSV [45] |
Simultaneously, in the context of fungal diseases, mycorrhizal fungi have been observed to enhance the resistance of plants to Fusarium wilt diseases caused by specific pathogenic Fusarium species. These pathogenic species, when infecting plants, can typically result in symptoms such as root rot and desiccation of the plant body [46]. Moreover, mycorrhizal fungi can also strengthen the plant’s defense against root rot caused by Rhizoctonia spp., a disease that is widespread in many crops and causes necrosis of the roots and stunted growth of the plant [47]. Regarding bacterial diseases, mycorrhizal fungi have also been demonstrated to reduce the incidence of leaf spot and canker diseases caused by bacteria such as Pseudomonas syringae [48].
2.2. The Molecular Mechanisms Underlying the Activation of Plant Disease Resistance by Mycorrhizal Fungi
The molecular mechanisms by which mycorrhizal fungi induce plant disease resistance involve a multitude of molecular changes and signal transduction pathways. Upon contact with plant roots, mycorrhizal fungi secrete signaling molecules [49], such as short-chain fatty acids and sugars, which are recognized by receptors on the root surface, initiating internal signal transduction pathways within the plant [50,51]. During this process, changes in intracellular calcium ion concentrations activate calcium-binding protein kinases (CDPKs), which further phosphorylate a series of downstream signaling molecules [52].These signaling molecules may include jasmonate synthases (JAS), which, under the action of CDPKs, convert precursor substances into jasmonic acid. As a signaling molecule, jasmonic acid activates its downstream signaling pathways, including jasmonic acid receptors (e.g., COI1) and related signaling molecules. The activation of the jasmonic acid signaling pathway leads to the phosphorylation and activation of specific transcription factors (e.g., MYC2). The activated transcription factors enter the nucleus and regulate the expression of genes related to disease resistance, including pathogenesis-related proteins (PR proteins) and other defense-related genes [53,54,55,56,57]. The expression of these genes enhances the plant’s defense mechanisms, including the reinforcement of cell walls, the production and activity of pathogen-associated proteins, and the generation of reactive oxygen species.
The presence of mycorrhizal fungi activates the antioxidant system within plants, potentially enhancing the activity of antioxidant enzymes such as cytosolic ascorbate peroxidase (cAPX), superoxide dismutase (SOD), peroxidase (POD), and glutathione reductase (GR) [58]. These enzymes help to eliminate reactive oxygen species produced during pathogen invasion, with cAPX being a primary scavenger of reactive oxygen. cAPX catalyzes the dehydrogenation reaction of H2O2 with ascorbate residues, converting H2O2 into water, thereby protecting plant cells from oxidative damage [59]. Phosphatases further enhance the plant’s antioxidant capacity by regulating the phosphorylation state of these antioxidant enzymes [60]. Hashem et al. [61] demonstrated that AMF can enhance plant disease resistance by promoting the activity of phosphatases. In this study, inoculation with AMF not only increased the activity of plant phosphorus metabolism-related enzymes but also enhanced the defense system by increasing chlorophyll content and improving water status, combating oxidative stress caused by Fusarium oxysporum f. sp. lycopersici (FOL). Furthermore, AMF can elevate the activity of the antioxidant enzyme system, which helps to eliminate reactive oxygen species and protect plants from oxidative damage, ensuring an effective response to pathogen attacks.
Mycorrhizal fungi also increase the absorption area of plant roots for nutrients in the soil through their mycelial network [62], especially key nutrients such as phosphorus and nitrogen, which are essential for the plant immune system [63]. Phosphatases can catalyze the hydrolysis of organic phosphorus compounds in the soil, releasing inorganic phosphate that is readily available for plant uptake, thereby enhancing the nutritional status of the plant [64]. This process not only improves the nutritional status of the plant but may also regulate the levels of plant hormones [65], such as salicylic acid (SA) and jasmonic acid (JA), indirectly enhancing the plant’s disease resistance.
In summary, the molecular mechanisms by which mycorrhizal fungi induce plant disease resistance involve a series of molecular changes at various levels, from signal recognition and transduction to gene expression regulation and the activation of the antioxidant system. Phosphatases play a key regulatory role in this process, enhancing plant resistance by finely tuning signal transduction pathways and gene expression.
3. The Role of Phosphatases in the Induction of Plant Disease Resistance by Mycorrhizal Fungi
Phosphatases play a multifaceted regulatory role in the plant disease resistance induced by mycorrhizal fungi, not only participating in the activation and modulation of signaling pathways, but also affecting the levels of plant hormones and the activity of the antioxidant system, together forming a complex defense network for plants against pathogens.
3.1. The Decomposition of Insoluble and Sparingly Soluble Phosphorus in the Soil
In soil, inorganic phosphorus, particularly that in mineral form, is readily adsorbed and fixed by soil particles, resulting in plant-available free phosphate concentrations typically below 10 µmol/L [66]. In acidic soils, phosphorus predominantly exists as iron-bound, aluminum-bound, and occluded forms, while in calcareous soils, it is mainly present in the form of insoluble calcium phosphates.
Mycorrhizal fungi, through their extensive mycelial networks, can expand the contact area between plant roots and soil [67], enhancing the activation and absorption of soil phosphorus [68,69]. Under acidification conditions, AMF play a crucial role in soil phosphorus cycling and plant nutrition, participating in the regulation of soil pH and the availability of phosphorus through various mechanisms. For instance, AMF can secrete organic acids and other chelating agents that compete with iron and aluminum ions in the soil, reducing the adsorption and fixation of phosphorus, thereby releasing more available phosphorus for plant uptake [70]. Simultaneously, mycorrhizal fungi secrete phosphatases that catalyze the hydrolysis of organic phosphorus compounds in the soil, thereby facilitating the dissolution and mineralization of insoluble phosphorus [71,72,73,74,75,76,77,78]. Consequently, phosphatases are considered key factors affecting the nutritional exchange between mycorrhizal fungi and their plant hosts [79].
Improved phosphorus nutrition promotes the growth of plant root systems, aiding in the adaptation to environmental stresses, such as drought and saline–alkali conditions, and enhances resistance to pathogens [80,81].
3.2. Involvement in the Regulation of Plant Defense and Immune Responses
Pattern recognition receptors (PRRs) on the surface of plant root cells perceive pathogen-associated molecular patterns (MAMPs), triggering an initial immune response known as pattern-triggered immunity (PTI) [82,83].
PTI involves the activation of intracellular signaling pathways, including the opening of calcium ion channels, the activation of protein kinases, and the production of reactive oxygen species (ROS), leading to the upregulation of defense-related gene expression, such as PR proteins and antimicrobial peptides [84,85,86], as well as the significant enhancement of defense-related genes (PR2a, PAL, and AOS) and the key gene BX9 in the DIMBOA (2,4-Dihydroxy-7-methoxy-2H-1,4-benzoxazin-3(4H)-one) biosynthetic pathway [87]. During the process of mycorrhiza formation, plant defense responses and molecular reprogramming are modulated to effectively activate the plant’s immune response and the expression of defense genes, which is highly similar to induced systemic resistance (ISR) [88,89] (Figure 3). This induced resistance is termed mycorrhiza-induced resistance (MIR). Phosphatases play a crucial role in ISR [90,91] and programmed cell death (PCD) by regulating the phosphorylation status of related proteins [92].

Figure 3.
Plant root and shoot defense initiation mechanism. (A) The mechanisms underlying induced systemic resistance (ISR) and systemic acquired resistance (SAR) involve distinct yet interconnected pathways. (B) Systemic acquired resistance represents an immune state established throughout the plant following local infection. SAR is primarily activated through the salicylic acid (SA) signaling pathway, endowing the plant with enhanced resistance to subsequent pathogen infections. (C) Induced systemic resistance is an immune response triggered by beneficial microbes or chemical substances perceived by the plant’s roots. This response is activated through signaling pathways mediated by plant hormones such as jasmonic acid (JA) and ethylene (ET), strengthening the plant’s defense against a broad spectrum of pathogens. Abbreviations: EIN3, Ethylene Insensitive 3; JAZ, Jasmonate Zim-Domain Protein; PAMPs, pathogen-associated molecular patterns; PRRs, pattern recognition receptors; JA, jasmonic acid; ET, ethylene; PRs, pathogenesis-related proteins; SA, salicylic acid; NPR1, non-race-specific disease resistance 1.
Phosphatases are primarily derived from fungal hyphae and hyphal sheaths (fungal cellular structures enveloping plant root tips), functioning by removing phosphate groups from signaling molecules [93]. It can modulate the activity and stability of immune-related proteins in plants, control the intensity and duration of signaling, and prevent excessive immune responses, thereby influencing plant’s pattern-triggered immunity (PTI) and effector-triggered immunity (ETI) responses [83].
In the intricate network of plant immunity, the mitogen-activated protein kinase (MAPK) cascade is a central signaling pathway. The activity of the MAPK can promote a local cell death mechanism known as the hypersensitive response (HR), which is part of the plant’s defense strategy [94]. Phosphatases regulate the PTI and ETI responses by dephosphorylating key components of the MAPK cascade, such as MAPKKK, MAPKK, and MAPK (Figure 4). This regulation not only affects the transmission of immune signals but also involves the activity and stability of specific transcription factors, such as WRKY transcription factors, thereby influencing the expression of genes related to SAR [95,96]. After the initial infection, WRKY transcription factors activate downstream resistance genes, helping the plant to establish systemic resistance in tissues distant from the site of primary infection [97]. Among them, the transcription factor WRKY33 is activated by the MPK3/6 kinase and directly binds to the promoter of the ALD1 gene, regulating its expression. The enzyme encoded by the ALD1 gene is responsible for the synthesis of pipecolic acid, a key mobile signal molecule in SAR [98].

Figure 4.
The immune response process of plant root systems against pathogenic agents. (A) Pattern-triggered immunity (PTI) process: Plant cells recognize pathogen-associated molecular patterns (PAMPs) through pattern recognition receptors (PRRs), which initiate a series of downstream signaling pathways. This includes the activation of mitogen-activated protein kinase kinase kinases (MAPKKKs), which in turn activate mitogen-activated protein kinase kinases (MAPKKs) and mitogen-activated protein kinases (MAPKs). The activation of these kinases leads to the conversion of ADP to ATP, providing energy for signal transduction. The signaling pathways activate calcium-dependent protein kinases (CDPKs) and other protein kinases, which further phosphorylate substrate proteins, facilitating intracellular signal transduction. These signals ultimately lead to the activation of transcription factors (TFs) in the nucleus, thereby inducing the expression of pathogenesis-related proteins (PRPs) and enhancing the plant’s defense response. Effector-triggered immunity (ETI) process: To combat PTI, pathogens secrete effector proteins. The recognition of these effectors by NLR proteins leads to their activation, which may involve oligomerization or conformational changes. The activation of NLR proteins initiates a potent immune response, characterized by intracellular signal transduction and changes in gene expression. This results in the binding of the activated NLR proteins to the effectors, inducing programmed cell death, such as the hypersensitive response (HR). (B) In the process of intracellular signal transduction in plant cells, the phosphorylation and dephosphorylation of proteins occur. This process is crucial for the regulation of various cellular functions and responses to environmental stimuli. Abbreviations: ROS, reactive oxygen species; CDPKs, calcium-dependent protein kinases; PPRs, pentatricopeptide repeats; TF, transcription factor; NB, nucleotide-binding site; LRR, leucine-rich repeat; PRPs, pathogenesis-related proteins; NLR, nucleotide-binding leucine-rich repeat; CPKs, calcium-dependent protein kinases; RBOHD, Respiratory Burst Oxidase Homologue D; RLP, receptor-like protein; NLRs, nucleotide-binding leucine-rich repeats.
Wei et al. [99] found that the phosphatase C-terminal domain phosphatase-like 1 (CPL1) plays a negative regulatory role in plant immunity, enhancing the plant’s immune response. CPL1 is localized in the nucleus and can interact with MKK4, MKK5, MPK3, and MPK6, disrupting the interaction between MKK4/MKK5 and MPK3/MPK6, and weakening the transmission of immune signals, thus negatively regulating Arabidopsis resistance to bacteria. This interference depends on the phosphatase activity of CPL1, revealing a new function of phosphatases in the regulatory network of plant immunity.
In addition, 2C-type protein phosphatases (PP2Cs) have been identified as key regulators of the antiviral defense mechanism [100,101,102]. Diao et al. [103] found that PP2C15 acts as a negative regulator of plant immunity by targeting the BRI1-associated receptor kinase 1 (BAK1). Among the 56 PP2Cs, 14 significantly suppressed the immune response induced by flg22, a bacterial pattern recognition molecule, with PP2C15 negatively regulating the immune response by interacting with BAK1 and dephosphorylating it.
Hence, it is clear that phosphatases play a critical role in modulating various aspects of plant defense and immune responses, such as signal transduction, regulation of transcription factor activity, and control of PR gene expression, making them an indispensable component of the plant’s disease resistance regulatory network.
3.3. The Interplay with Hormonal Signaling
Plant hormones such as jasmonic acid (JA), salicylic acid (SA), auxin, and gibberellin (GA) are pivotal signaling molecules in the plant immune system. The transcription factor OsARF17, a key player in the auxin signaling pathway, is implicated in the regulation of plant responses to various biotic stresses, including viral infections [104]. Plants may balance immune responses and growth through such hormonal crosstalk. Specifically, the salicylic acid receptor NPR1, as a subunit of the ubiquitin E3 ligase, can promote the polyubiquitination and degradation of the GA receptor GID1, enhancing the stability of DELLA proteins, which are negative regulators of the GA signaling pathway [105]. Meanwhile, CPL3, a phosphatase associated with the GA response, interacts with DELLA proteins and is involved in the phosphorylation of RNA polymerase II, playing an important role in the regulation of plant growth and development [106]. Phosphatases may also enhance the systemic resistance of plants by regulating the SA and JA signaling pathways [107,108]. For example, by participating in the synthesis, signal transduction, or response of proteins involved in SA, they can affect plant resistance to diseases. This regulatory effect may involve the phosphorylation status of multiple levels of proteins in the SA signaling pathway [109], thereby affecting the plant’s recognition of pathogens, signal amplification, and the activation of the final defense response [103]. In addition to SA, other oxylipins may also be involved in signal transduction during the interaction between mycorrhizal fungi and pathogens in the roots [110].
It can be seen that the roles of plant hormones and phosphatases in the plant immune system are interconnected and interdependent. Together, they form a complex regulatory network that interacts with other hormonal signaling pathways, either directly or indirectly, and participates in the plant’s defense mechanisms against pathogens.
3.4. Restriction the Invasion and Spread of Pathogens
Fernandes et al. [111] have discovered that Fusarium oxysporum can induce rapid alkalinization of the extracellular space in the host by secreting a functional homologue of the plant’s Rapid ALkalinizing Factor (RALF), known as the F-RALF peptide. This alkalization effect is crucial for the pathogen’s infection process, as it not only facilitates the invasive growth of the fungus but also activates cellular signaling pathways associated with pathogenicity, thereby enhancing its virulence.
Specifically, such alkalinization can shift the pH milieu within host tissues, thereby providing a more propitious environment for pathogen proliferation. Additionally, this alkalization may indirectly impact the functionality of the host cells, including intracellular signaling pathways, potentially aiding the efficacy of effector proteins secreted by the pathogen to manipulate the host’s cellular machinery. However, phosphatases secreted by mycorrhizal fungi can dephosphorylate these effector proteins, reducing their activity and thereby limiting the infection and spread of pathogens [112,113,114]. In addition, specific phosphatases secreted by mycorrhizal fungi can hydrolyze polysaccharides in the pathogen cell wall, weakening the pathogen’s ability to infect [115], and activating the plant’s immune response, providing an effective defense mechanism for the plant [116].
Simultaneously, there is competition between mycorrhizal fungi and pathogens for resources around the plant roots, which helps to restrict the growth and spread of pathogens. Mycorrhizal fungi can reduce the chances of pathogen infection through competition and antagonism. Phosphatases may also interfere with the quorum-sensing mechanism of pathogens by hydrolyzing quorum sensing signal molecules, such as AHLs (N-acyl homoserine lactones) [117], reducing the production of virulence factors by pathogens and thus decreasing their pathogenicity to the host.
3.5. The Interaction between Mycorrhizal Fungi and Beneficial Microorganisms
3.5.1. Mycorrhizal Fungi and Beneficial Microbes Synergize to Enhance Crop Disease Resistance
Caravaca et al. [118] found that the Streptomyces AcH505 strain enhances the growth of oak trees by increasing the abundance of saprotrophic and ectomycorrhizal fungi in the rhizosphere (an increase of 158% compared to the control group), which can counteract the damage caused by nematodes by promoting the growth of oak micro-cuttings. This indicates that mycorrhizal fungi form a symbiotic relationship with plant roots, which not only promotes the absorption of nutrients by the plant but also changes the composition of the microbial community in the plant’s rhizosphere [119,120,121,122,123,124]. Through interactions with various beneficial microorganisms, they inhibit the activity of pathogens and jointly enhance the plant’s disease resistance [125,126,127,128] (Table 2).
Beneficial microorganisms can promote the formation of mycorrhizae by secreting compounds that stimulate the growth of mycorrhizal fungi, such as hormones, enzymes, and organic acids [129,130,131]. At the same time, they can also degrade toxic substances in the soil [132], protecting mycorrhizal fungi from the effects of harmful substances. During the interaction process, mycorrhizal fungi and these beneficial microorganisms can inhibit the growth of pathogens by producing antibiotics, siderophores, and SAR signaling molecules, improve the nutritional status of plants, activate various defense-related genes in plants, and change the composition of root exudates to regulate the plant’s sensitivity to pathogens, thereby enhancing the plant’s physiological health and resistance to environmental stress, and thus improving the plant’s resistance to pathogens [133,134,135,136,137,138,139].
Table 2.
Mechanisms by which mycorrhizal fungi synergize with beneficial microbes to enhance crop resistance to diseases.
Table 2.
Mechanisms by which mycorrhizal fungi synergize with beneficial microbes to enhance crop resistance to diseases.
| Disease Name | Pathogenic Microorganism | Beneficial Microbial Species | Host Plant | Mycorrhizal Fungal Names | Mycorrhizal Types | Synergistic Mechanism |
|---|---|---|---|---|---|---|
| Bacterial wilt | Ralstonia solanacearum | Trichoderma harzianum | Nicotiana tabacum | Glomus mosseae | Arbuscular mycorrhizae | The synergistic action of these two factors has led to a reduction in the abundance of soil-borne pathogenic microorganisms and a concomitant enhancement of the activity of plant systemic resistance-related enzymes [40] |
| Ralstonia solanacearum | Bacillus spp., Pseudomonas spp., Azotobacter spp. | Solanum tuberosum | Glomus intraradices, G. etunicatum, G. mosseae | Arbuscular mycorrhizae | The combination of biocontrol agents (BCA) and arbuscular mycorrhizal fungi (AMF) may mitigate disease severity through antagonistic interactions and influence the microbial community by altering root exudates [140] | |
| Spring black stem and leaf spot | Phoma medicaginis | Sinorhizobium medicae | Medicago sativa | Funneliformis mosseae | Arbuscular mycorrhizae | The mutual promotion between mycorrhizal fungi and other microbes enhances the formation of root nodules and mycorrhizal colonization, which in turn boosts the plant’s phosphorus and nitrogen uptake. This interaction also augments the activity of plant defense compounds and enzymes, consequently reducing the disease index [141] |
| Anthracnose | Colletotrichum orbiculare | Phoma sp., Penicillium simplicissimum | Cucumis sativus | Glomus mosseae | Arbuscular mycorrhizae | The interplay between Plant growth-promoting fungi (PGPF) and arbuscular mycorrhizal fungi (AMF) may influence the level of disease protection through competitive interactions for space or nutrients [142] |
| Fusarium wilt | Fusarium oxysporum | Trichoderma harzianum | Cucumis melo | Glomus constrictum, G. mosseae, G. claroideum, G. intraradices | Arbuscular mycorrhizae | Trichoderma harzianum and arbuscular mycorrhizal fungi (AMF) may synergistically control diseases by enhancing nutrient uptake and inducing systemic resistance in plants [143] |
| Bipolaris sorokiniana | Bipolaris sorokiniana | Epichloë festucae | Lolium perenne | Claroideoglomus etunicatum | Arbuscular mycorrhizae | Endophytic fungi and mycorrhizal fungi enhance the resistance of plants to diseases by activating defense-related enzymes, increasing the activity of plant hormones, and elevating the content of lignin [144] |
| Root rot and charcoal rot | Macrophomina phaseolina | Brettanomyces naardensis | Helianthus annuus | Acaulospora bireticulata | Arbuscular mycorrhizae | Yeasts facilitate the development of arbuscular mycorrhizal fungi (AMF) by supplying vitamin B12. The combined action of these two organisms alters the pattern of root exudates, impacting the plant’s rhizosphere microbial community and inhibiting the invasion and growth of pathogenic fungi [145] |
| Late blight | Phytophthora infestans | Pseudomonas sp. | Solanum tuberosum | Rhizophagus irregularis | Arbuscular mycorrhizae | The co-inoculation of plant growth-promoting microbes and arbuscular mycorrhizal fungi may activate the plant’s systemic defense system, leading to the upregulation of ethylene response factor 3 (ERF3) and thereby enhancing the plant’s resistance to diseases [146] |
| Take-all | Gaeumannomyces graminis | Pseudomonas fluorescens | Triticum aestivum | Glomus mossea | Arbuscular mycorrhizae | Mycorrhizal fungi enhance the plant’s resistance to diseases, while beneficial microbes influence plant metabolism or directly inhibit pathogen growth through their metabolic byproducts [147] |
| Root-knot nematode disease | Meloidogyne incognita | Bacillus polymyxa, Bacillus sp. | Solanum lycopersicum | Glomus versiforme, Glomus mosseae | Arbuscular mycorrhizae | Beneficial microbes augment the colonization of arbuscular mycorrhizal (AM) fungi in the roots, and in turn, AM fungi enhance the population of beneficial microbes in the rhizosphere; together, they suppress nematode damage and promote plant growth [148] |
| Sphaeropsis Shoot Blight | Sphaeropsis sapinea | Bacillus pumilus | Pinus thunbergii | Hymenochaete sp. Rl | Ectomycorrhizae | Mycorrhizal fungi elicit systemic defense responses in plants. In concert, beneficial bacteria facilitate the formation of symbiotic structures between the mycorrhizal fungi and their host plants [149] |
| White rot | Sclerotinia sclerotiorum | PGPR | Fragaria | AMF | Arbuscular mycorrhizae | The combined application of mycorrhizal fungi and plant growth-promoting rhizobacteria (PGPR) enhances plant biomass, promotes vegetative growth, and reduces disease indices [150] |
3.5.2. The Role of Phosphatases in the Synergistic Process
Abdel-Fattah et al. [151] found that Sorghum bicolor inoculated with the arbuscular mycorrhizal fungus Glomus intraradices no. LAP8 had significantly higher acid and alkaline phosphatase activities in root extracts compared to non-mycorrhizal plants not inoculated with the fungus. The increase in phosphatase activity leads to the enhanced availability of phosphorus in the soil [152], which can improve soil fertility [153], create more suitable living conditions for beneficial microbes, provide more nutrients, promote the growth and metabolic activities of beneficial microbes, and thus affect the structure and diversity of the soil microbial community [154,155].
Furthermore, the enhancement of phosphatase activity can protect the host plant from changes induced by ionic and osmotic stress [156], and promote the growth and extension of mycorrhizal fungal hyphae [157], thereby increasing the opportunities for contact with other beneficial microbes in the soil, forming a more complex microbial network, and enhancing the plant’s disease resistance.
Thus, the activity of phosphatases is closely related to soil fertility, the symbiotic relationship with mycorrhizal fungi, and the structure and function of the soil microbial community.
4. Perspectives
Current research collectively underscores the pivotal role of mycorrhizal fungi in the absorption and translocation of essential nutrients within symbiotic relationships. Mycorrhizal fungi enhance plant immunity by improving the nutritional status of the host, particularly in terms of phosphorus uptake. Moreover, the mycelial networks formed by mycorrhizal fungi facilitate the exchange of materials and signal transmission among plants, aiding in the collective defense against diseases within plant communities.
The resistance conferred by mycorrhizal fungi through the action of phosphatases is a relatively complex process. Although phosphatases themselves do not directly combat pathogenic organisms, they contribute to the plant defense system by modulating plant hormone signaling pathways, enhancing nutritional status, disrupting the quorum-sensing mechanisms of pathogens, and activating plant immune responses. The role of phosphatases in immune regulation is multifaceted; they are involved not only in the fundamental physiological processes of the plant but also in the response and modulation to pathogen attacks. However, the mechanisms underlying resistance remain contentious, necessitating further integration of molecular techniques and physiological experiments to elucidate the specific mechanisms by which phosphatases enhance plant disease resistance in mycorrhizal associations.
The involvement of phosphatases in induced plant resistance encompasses a variety of signaling molecules and metabolic pathways, providing significant clues for further dissection of the molecular mechanisms underlying plant-microbe interactions. However, the specific roles and molecular mechanisms of phosphatases in different mycorrhizal fungi and plant systems remain to be further elucidated. Additionally, how phosphatases interact with other plant defense signaling pathways and how these interactions influence plant resistance to various pathogens are key points for future research. Furthermore, the impact of environmental factors such as soil pH, nutrient status, and climate change on the secretion of phosphatases by mycorrhizal fungi, and how these factors regulate plant resistance, are also important directions for future investigation.
Further research in this field can pave new avenues and methods for plant disease control and the study of mycorrhizal fungi, offering valuable insights for sustainable strategies to enhance crop productivity.
Author Contributions
Conceptualization, L.C., X.Z., L.Y., Q.L. and B.Z.; Writing—Original Draft Preparation, L.C. and X.Z.; Writing—Review and Editing, L.C., X.Z. and X.L.; Supervision, X.Z., X.Y., Y.H. and X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Sichuan Province Science and Technology Plan Project (2021YFYZ0026, 2024YFHZ0165), the Shandong Province Science and Technology Plan Project(Grant No. 2022TSGC2404) and the Sichuan Mushroom Innovation Team (SCCXTD-2024-07).
Data Availability Statement
All data analyzed during this study are included in this article.
Acknowledgments
The figures were created with figdraw2.0 https://www.figdraw.com (accessed on 14 July 2024).
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this manuscript.
References
- Frank, B. Über die auf Wurzelsymbiose beruhende Ernährung gewisser Bäume durch unterirdische Pilze. Plant Biol. 1885, 3, 128–145. [Google Scholar]
- Dickie, I.A.; Koide, R.T.; Steiner, K.C. Influences of established trees on mycorrhizas, nutrition, and growth of Quercus rubra seedlings. Ecol. Monogr. 2002, 72, 505–521. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Yamanaka, T.; Yoshimura, K.; Kosugi, Y. Ectomycorrhizal-fungal colonization induces physio-morphological changes in Quercus serrata leaves and roots. J. Plant Nutr. Soil Sci. 2012, 175, 900–906. [Google Scholar] [CrossRef]
- Guo, Y.; Martin, K.; Hrynkiewicz, K.; Rasche, F. Arbuscular mycorrhizal fungi-based bioremediation of mercury: Insights from zinc and cadmium transporter studies. Int. J. Environ. Sci. Technol. 2024, 21, 3475–3488. [Google Scholar] [CrossRef]
- Stallmann, J.; Schweiger, R. Effects of arbuscular mycorrhiza on primary metabolites in phloem exudates of Plantago major and Poa annua and on a generalist aphid. Int. J. Mol. Sci. 2021, 22, 13086. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Corcobado, T.; Vivas, M.; Moreno, G.; Solla, A. Ectomycorrhizal symbiosis in declining and non-declining Quercus ilex trees infected with or free of Phytophthora cinnamomi. For. Ecol. Manag. 2014, 324, 72–80. [Google Scholar] [CrossRef]
- Perrin, R. Interactions between mycorrhizae and diseases caused by soil-borne fungi. Soil Use Manag. 1990, 6, 189–194. [Google Scholar] [CrossRef]
- Raymond, N.S.; Gómez-Muñoz, B.; van der Bom, F.J.; Nybroe, O.; Jensen, L.S.; Müller-Stöver, D.S.; Oberson, A.; Richardson, A.E. Phosphate-solubilising microorganisms for improved crop productivity: A critical assessment. New Phytol. 2021, 229, 1268–1277. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, G.; Declerck, S. Signal beyond nutrient, fructose, exuded by an arbuscular mycorrhizal fungus triggers phytate mineralization by a phosphate solubilizing bacterium. ISME J. 2018, 12, 2339–2351. [Google Scholar] [CrossRef]
- Crombez, H.; Motte, H.; Beeckman, T. Tackling plant phosphate starvation by the roots. Dev. Cell 2019, 48, 599–615. [Google Scholar] [CrossRef] [PubMed]
- Bücking, H.; Liepold, E.; Ambilwade, P. The role of the mycorrhizal symbiosis in nutrient uptake of plants and the regulatory mechanisms underlying these transport processes. Plant Sci. 2012, 4, 108–132. [Google Scholar]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic press: Cambridge, MA, USA, 2010. [Google Scholar]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef]
- Tang, M.; Zhang, R.-Q.; Chen, H.; Zhang, H.-H.; Tian, Z.-Q. Induced hydrolytic enzymes of ectomycorrhizal fungi against pathogen Rhizoctonia solani. Biotechnol. Lett. 2008, 30, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Dreischhoff, S.; Das, I.S.; Jakobi, M.; Kasper, K.; Polle, A. Local responses and systemic induced resistance mediated by ectomycorrhizal fungi. Front. Plant Sci. 2020, 11, 590063. [Google Scholar] [CrossRef] [PubMed]
- Ortas, I.; Rafique, M.; Çekiç, F. Do Mycorrhizal Fungi enable plants to cope with abiotic stresses by overcoming the detrimental effects of salinity and improving drought tolerance? In Symbiotic Soil Microorganisms: Biology and Applications; Springer International Publishing: Berlin/Heidelberg, Germany, 2021; pp. 391–428. [Google Scholar]
- Tapia-Vázquez, I.; Montoya-Martínez, A.C.; De los Santos-Villalobos, S.; Ek-Ramos, M.J.; Montesinos-Matías, R.; Martínez-Anaya, C. Root-knot nematodes (Meloidogyne spp.) a threat to agriculture in Mexico: Biology, current control strategies, and perspectives. World J. Microbiol. Biotechnol. 2022, 38, 26. [Google Scholar] [CrossRef]
- Stukenbrock, E.; McDonald, B. The origins of plant pathogens in agro-ecosystems. Annu. Rev. Phytopathol. 2008, 46, 75–100. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; Hamss, H.E.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Mian, G.; Zuiderduin, K.; Barnes, L.S.; Loketsatian, S.; Bell, L.; Ermacora, P.; Cipriani, G. In vitro application of Eruca vesicaria subsp. sativa leaf extracts and associated metabolites reduces the growth of Oomycota species involved in Kiwifruit Vine Decline Syndrome. Front. Plant Sci. 2023, 14, 1292290. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Boguś, M. Fungi of entomopathogenic potential in Chytridiomycota and Blastocladiomycota, and in fungal allies of the Oomycota and Microsporidia. IMA Fungus 2021, 12, 29. [Google Scholar] [CrossRef]
- Gallou, A.; Mosquera, H.P.L.; Cranenbrouck, S.; Suárez, J.P.; Declerck, S. Mycorrhiza induced resistance in potato plantlets challenged by Phytophthora infestans. Physiol. Mol. Plant Pathol. 2011, 76, 20–26. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.H.; Abdelgani, M.E.; Yassin, A. Effects of interaction between Vesicular-Arbuscular Mycorrhizal (VAM) fungi and root-knot nematodes on Dolichos bean (Lablab niger Medik.) plants. Am.-Eurasian J. Sustain. Agric. 2009, 3, 678–683. [Google Scholar]
- Aseel, D.G.; Rashad, Y.M.; Hammad, S.M. Arbuscular mycorrhizal fungi trigger transcriptional expression of flavonoid and chlorogenic acid biosynthetic pathways genes in tomato against Tomato Mosaic Virus. Sci. Rep. 2019, 9, 9692. [Google Scholar] [CrossRef] [PubMed]
- Steinkellner, S.; Hage-Ahmed, K.; García-Garrido, J.M.; Illana, A.; Ocampo, J.A.; Vierheilig, H. A comparison of wild-type, old and modern tomato cultivars in the interaction with the arbuscular mycorrhizal fungus Glomus mosseae and the tomato pathogen Fusarium oxysporum f. sp. lycopersici. Mycorrhiza 2012, 22, 189–194. [Google Scholar] [CrossRef]
- Pu, C.; Ge, Y.; Yang, G.; Zheng, H.; Guan, W.; Chao, Z.; Shen, Y.; Liu, S.; Chen, M.; Huang, L. Arbuscular mycorrhizal fungi enhance disease resistance of Salvia miltiorrhiza to Fusarium wilt. Front. Plant Sci. 2022, 13, 975558. [Google Scholar] [CrossRef]
- Lin, P.; Zhang, M.; Wang, M.; Li, Y.; Liu, J.; Chen, Y. Inoculation with arbuscular mycorrhizal fungus modulates defense-related genes expression in banana seedlings susceptible to wilt disease. Plant Signal. Behav. 2021, 16, 1884782. [Google Scholar] [CrossRef]
- Li, W.; Zhu, C.; Song, Y.; Yuan, Y.; Li, M.; Sun, Y. Arbuscular mycorrhizal fungi by inducing watermelon roots secretion phthalates, altering soil enzyme activity and bacterial community composition to alleviate the watermelon wilt. BMC Plant Biol. 2024, 24, 593. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Zlatković, M.; Stojnic, S.; Čapelja, E.; Zorić, M.; Kiprovski, B.; Budakov, D.; Orlović, S. Ectomycorrhizal fungi modulate biochemical response against powdery mildew disease in Quercus robur L. Forests 2022, 13, 1491. [Google Scholar] [CrossRef]
- Pfabel, C.; Eckhardt, K.-U.; Baum, C.; Struck, C.; Frey, P.; Weih, M. Impact of ectomycorrhizal colonization and rust infection on the secondary metabolism of poplar (Populus trichocarpa × deltoides). Tree Physiol. 2012, 32, 1357–1364. [Google Scholar] [CrossRef]
- D Aljawasim, B.; M Khaeim, H.; A Manshood, M. Assessment of arbuscular mycorrhizal fungi (Glomus spp.) as potential biocontrol agents against damping-off disease Rhizoctonia solani on cucumber. J. Crop Prot. 2020, 9, 141–147. [Google Scholar]
- Kobra, N.; Jalil, K.; Youbert, G. Arbuscular mycorrhizal fungi and biological control of Verticillium-wilted cotton plants. Arch. Phytopathol. Plant Prot. 2011, 44, 933–942. [Google Scholar] [CrossRef]
- Zhang, Q.; Gao, X.; Ren, Y.; Ding, X.; Qiu, J.; Li, N.; Zeng, F.; Chu, Z. Improvement of Verticillium wilt resistance by applying arbuscular mycorrhizal fungi to a cotton variety with high symbiotic efficiency under field conditions. Int. J. Mol. Sci. 2018, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.; Jakobsen, I.; Lyngkjær, M.F.; Thordal-Christensen, H.; Pons-Kühnemann, J. Arbuscular mycorrhiza reduces susceptibility of tomato to Alternaria solani. Mycorrhiza 2006, 16, 413–419. [Google Scholar] [CrossRef]
- Tchameni, S.N.; Ngonkeu, M.; Begoude, B.; Nana, L.W.; Fokom, R.; Owona, A.; Mbarga, J.; Tchana, T.; Tondje, P.; Etoa, F. Effect of Trichoderma asperellum and arbuscular mycorrhizal fungi on cacao growth and resistance against black pod disease. Crop Prot. 2011, 30, 1321–1327. [Google Scholar] [CrossRef]
- Petit, E.; Gubler, W.D. Influence of Glomus intraradices on black foot disease caused by Cylindrocarpon macrodidymum on Vitis rupestris under controlled conditions. Plant Dis. 2006, 90, 1481–1484. [Google Scholar] [CrossRef]
- Tahat, M.M.; Sijam, K.; Othman, R. The potential of endomycorrhizal fungi in controlling tomato bacterial wilt Ralstonia solanacearum under glasshouse conditions. Afr. J. Biotechnol. 2012, 11, 13085–13094. [Google Scholar]
- Yuan, S.; Li, M.; Fang, Z.; Liu, Y.; Shi, W.; Pan, B.; Wu, K.; Shi, J.; Shen, B.; Shen, Q. Biological control of tobacco bacterial wilt using Trichoderma harzianum amended bioorganic fertilizer and the arbuscular mycorrhizal fungi Glomus mosseae. Biol. Control 2016, 92, 164–171. [Google Scholar] [CrossRef]
- Taiwo, L.; Adebayo, D.; Adebayo, O.; Adediran, J. Compost and Glomus mosseae for management of bacterial and Fusarium wilts of tomato. Int. J. Veg. Sci. 2007, 13, 49–61. [Google Scholar] [CrossRef]
- Chave, M.; Crozilhac, P.; Deberdt, P.; Plouznikoff, K.; Declerck, S. Rhizophagus irregularis MUCL 41833 transitorily reduces tomato bacterial wilt incidence caused by Ralstonia solanacearum under in vitro conditions. Mycorrhiza 2017, 27, 719–723. [Google Scholar] [CrossRef]
- Vos, C.; Tesfahun, A.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi induce systemic resistance in tomato against the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans. Appl. Soil Ecol. 2012, 61, 1–6. [Google Scholar] [CrossRef]
- Lax, P.; Becerra, A.G.; Soteras, F.; Cabello, M.; Doucet, M.E. Effect of the arbuscular mycorrhizal fungus Glomus intraradices on the false root-knot nematode Nacobbus aberrans in tomato plants. Biol. Fertil. Soils 2011, 47, 591–597. [Google Scholar] [CrossRef]
- Maffei, G.; Miozzi, L.; Fiorilli, V.; Novero, M.; Lanfranco, L.; Accotto, G.P. The arbuscular mycorrhizal symbiosis attenuates symptom severity and reduces virus concentration in tomato infected by Tomato yellow leaf curl Sardinia virus (TYLCSV). Mycorrhiza 2014, 24, 179–186. [Google Scholar] [CrossRef]
- Sharma, R.; Rajak, R.C.; Pandey, A.K. Evidence of antagonistic interactions between rhizosphere and mycorrhizal fungi associated with Dendrocalamus strictus (Bamboo). J. Yeast Fungal Res. 2010, 1, 112–117. [Google Scholar]
- Loit, K.; Soonvald, L.; Adamson, K.; Runno-Paurson, E.; Tedersoo, L.; Astover, A. Assessing the effect of commercial arbuscular mycorrhizal fungal inoculum on potato plant disease incidence, yield and the indigenous root fungal community composition. BioControl 2023, 68, 537–547. [Google Scholar] [CrossRef]
- Garcia-Garrido, J.; Ocampo, J. Effect of VA mycorrhizal infection of tomato on damage caused by Pseudomonas syringae. Soil Biol. Biochem. 1989, 21, 165–167. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-induced resistance and priming of plant defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Miransari, M. Plant, mycorrhizal fungi, and bacterial network. In Plant Signaling: Understanding the Molecular Crosstalk; Springer: Berlin/Heidelberg, Germany, 2014; pp. 315–325. [Google Scholar]
- He, J.; Dai, H.; Zhang, X.; Wang, E. Mycorrhizal signals promote root development dependent on LysM-receptor like kinases in rice. New Crops 2024, 1, 100009. [Google Scholar] [CrossRef]
- Liese, A.; Eichstädt, B.; Lederer, S.; Schulz, P.; Oehlschläger, J.; Matschi, S.; Feijó, J.A.; Schulze, W.X.; Konrad, K.R.; Romeis, T. Imaging of plant calcium-sensor kinase conformation monitors real time calcium-dependent decoding in planta. Plant Cell 2024, 36, 276–297. [Google Scholar] [CrossRef]
- Li, S.; Xiong, Y.; Ralf, M.-X.; Xing, Q. Distinct expression patterns of WRKY 6 and PR 1 in Arabidopsis stress memory assays. Bull. Bot. Res. 2019, 39, 752–759. [Google Scholar]
- Jain, D.; Khurana, J.P. Role of pathogenesis-related (PR) proteins in plant defense mechanism. In Molecular Aspects of Plant-Pathogen Interaction; Springer: Berlin/Heidelberg, Germany, 2018; pp. 265–281. [Google Scholar]
- Malviya, D.; Singh, P.; Singh, U.B.; Paul, S.; Kumar Bisen, P.; Rai, J.P.; Verma, R.L.; Fiyaz, R.A.; Kumar, A.; Kumari, P. Arbuscular mycorrhizal fungi-mediated activation of plant defense responses in direct seeded rice (Oryza sativa L.) against root-knot nematode Meloidogyne graminicola. Front. Microbiol. 2023, 14, 1104490. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.; Cai, X.; Han, Z.; Si, J.; Chen, D. Jasmonate signaling pathway modulates plant defense, growth, and their trade-offs. Int. J. Mol. Sci. 2022, 23, 3945. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; San Segundo, B. Systemic induction of phosphatidylinositol-based signaling in leaves of arbuscular mycorrhizal rice plants. Sci. Rep. 2020, 10, 15896. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-J.; Tong, C.-L.; Wang, Q.-S.; Bie, S. Mycorrhizas Affect Physiological Performance, Antioxidant System, Photosynthesis, Endogenous Hormones, and Water Content in Cotton under Salt Stress. Plants 2024, 13, 805. [Google Scholar] [CrossRef] [PubMed]
- Camillo, L.R.; Filadelfo, C.R.; Monzani, P.S.; Corrêa, R.X.; Gramacho, K.P.; Micheli, F.; Pirovani, C.P. Tc-cAPX, a cytosolic ascorbate peroxidase of Theobroma cacao L. engaged in the interaction with Moniliophthora perniciosa, the causing agent of witches’ broom disease. Plant Physiol. Biochem. 2013, 73, 254–265. [Google Scholar] [CrossRef][Green Version]
- Rahikainen, M.; Pascual, J.; Alegre, S.; Durian, G.; Kangasjärvi, S. PP2A phosphatase as a regulator of ROS signaling in plants. Antioxidants 2016, 5, 8. [Google Scholar] [CrossRef]
- Hashem, A.; Akhter, A.; Alqarawi, A.A.; Singh, G.; Almutairi, K.F.; Abd_Allah, E.F. Mycorrhizal fungi induced activation of tomato defense system mitigates Fusarium wilt stress. Saudi J. Biol. Sci. 2021, 28, 5442–5450. [Google Scholar] [CrossRef]
- Xu, F.-Q.; Meng, L.-L.; Kuča, K.; Wu, Q.-S. The mechanism of arbuscular mycorrhizal fungi-alleviated manganese toxicity in plants: A review. Plant Physiol. Biochem. 2024, 213, 108808. [Google Scholar] [CrossRef]
- Wang, G.; Jin, Z.; George, T.S.; Feng, G.; Zhang, L. Arbuscular mycorrhizal fungi enhance plant phosphorus uptake through stimulating hyphosphere soil microbiome functional profiles for phosphorus turnover. New Phytol. 2023, 238, 2578–2593. [Google Scholar] [CrossRef]
- George, T.S.; Giles, C.D.; Menezes-Blackburn, D.; Condron, L.M.; Gama-Rodrigues, A.C.; Jaisi, D.; Lang, F.; Neal, A.L.; Stutter, M.I.; Almeida, D.S. Organic phosphorus in the terrestrial environment: A perspective on the state of the art and future priorities. Plant Soil. 2018, 427, 191–208. [Google Scholar] [CrossRef]
- Li, P.; Liu, J. Protein phosphorylation in plant cell signaling. Plant Phosphoproteomics Methods Protoc. 2021, 2358, 45–71. [Google Scholar]
- Becquer, A.; Garcia, K.; Amenc, L.; Rivard, C.; Doré, J.; Trives-Segura, C.; Szponarski, W.; Russet, S.; Baeza, Y.; Lassalle-Kaiser, B. The Hebeloma cylindrosporum HcPT2 Pi transporter plays a key role in ectomycorrhizal symbiosis. New Phytol. 2018, 220, 1185–1199. [Google Scholar] [CrossRef]
- Wang, Y.; He, X.; Yu, F. Non-host plants: Are they mycorrhizal networks players? Plant Divers. 2022, 44, 127–134. [Google Scholar] [CrossRef]
- Kafle, A.; Cope, K.R.; Raths, R.; Krishna Yakha, J.; Subramanian, S.; Bücking, H.; Garcia, K. Harnessing soil microbes to improve plant phosphate efficiency in cropping systems. Agronomy 2019, 9, 127. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, B.; Zheng, S.; Zhang, X.; Wang, X.; Dong, W.; Xie, Q.; Wang, G.; Xiao, Y.; Chen, F. A phosphate starvation response-centered network regulates mycorrhizal symbiosis. Cell 2021, 184, 5527–5540.e18. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Hui, D.; Wang, Y.P.; Li, J.; Chen, J.; Chen, G.; Zhu, Y.; Zhang, L.; Zhang, D. Mycorrhizal fungi alleviate acidification-induced phosphorus limitation: Evidence from a decade-long field experiment of simulated acid deposition in a tropical forest in south China. Glob. Change Biol. 2022, 28, 3605–3619. [Google Scholar] [CrossRef]
- Louche, J.; Ali, M.A.; Cloutier-Hurteau, B.; Sauvage, F.X.; Quiquampoix, H.; Plassard, C. Efficiency of acid phosphatases secreted from the ectomycorrhizal fungus Hebeloma cylindrosporum to hydrolyse organic phosphorus in podzols. FEMS Microbiol. Ecol. 2010, 73, 323–335. [Google Scholar] [CrossRef]
- Courty, P.-E.; Labbé, J.; Kohler, A.; Marçais, B.; Bastien, C.; Churin, J.-L.; Garbaye, J.; Le Tacon, F. Effect of poplar genotypes on mycorrhizal infection and secreted enzyme activities in mycorrhizal and non-mycorrhizal roots. J. Exp. Bot. 2011, 62, 249–260. [Google Scholar] [CrossRef]
- Garcia, K.; Delteil, A.; Conéjéro, G.; Becquer, A.; Plassard, C.; Sentenac, H.; Zimmermann, S. Potassium nutrition of ectomycorrhizal Pinus pinaster: Overexpression of the H ebeloma cylindrosporum H c T rk1 transporter affects the translocation of both K+ and phosphorus in the host plant. New Phytol. 2014, 201, 951–960. [Google Scholar] [CrossRef]
- Sardans, J.; Lambers, H.; Preece, C.; Alrefaei, A.F.; Penuelas, J. Role of mycorrhizas and root exudates in plant uptake of soil nutrients (calcium, iron, magnesium, and potassium): Has the puzzle been completely solved? Plant J. 2023, 114, 1227–1242. [Google Scholar] [CrossRef]
- Baghel, R.; Sharma, R.; Pandey, A. Activity of acid phosphatase in the ectomycorrhizal fungus Cantharellus tropicalis under controlled conditions. J. Trop. For. Sci. 2009, 21, 218–222. [Google Scholar]
- Meeds, J.A.; Kranabetter, J.M.; Zigg, I.; Dunn, D.; Miros, F.; Shipley, P.; Jones, M.D. Phosphorus deficiencies invoke optimal allocation of exoenzymes by ectomycorrhizas. ISME J. 2021, 15, 1478–1489. [Google Scholar] [CrossRef] [PubMed]
- Plassard, C.; Fransson, P. Regulation of low-molecular weight organic acid production in fungi. Fungal Biol. Rev. 2009, 23, 30–39. [Google Scholar] [CrossRef]
- Rúa, M.A.; Moore, B.; Hergott, N.; Van, L.; Jackson, C.R.; Hoeksema, J.D. Ectomycorrhizal fungal communities and enzymatic activities vary across an ecotone between a forest and field. J. Fungi 2015, 1, 185–210. [Google Scholar] [CrossRef]
- Miyauchi, S.; Kiss, E.; Kuo, A.; Drula, E.; Kohler, A.; Sánchez-García, M.; Morin, E.; Andreopoulos, B.; Barry, K.W.; Bonito, G. Large-scale genome sequencing of mycorrhizal fungi provides insights into the early evolution of symbiotic traits. Nat. Commun. 2020, 11, 5125. [Google Scholar] [CrossRef]
- Hai-Yan, F.; Gu, F.; Jian-Lan, S.; Jing-Guo, W.; Xiao-Lin, L. Studies on the relationship between the activity of alkaline phosphatase in intraradical hyphae of arbuscular mycorrhizae fungi and efficiency of mycorrhizal symbiosis. Chin. J. Eco-Agric. 2004, 12, 124–127. [Google Scholar]
- Ang, Y.-f.; Guo, Y.; Chen, H.-y.; Liu, R.-y.; Zhu, Q.-q.; Wang, L.; Zhu, Y.-y.; Yi, K.-k.; Zeng, H.-q. Interaction between plant phosphorus nutrition and abiotic stress responses and its potential application in agricultural production. J. Plant Nutr. Fertil. 2023, 29, 2345–2359. [Google Scholar]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.-M.; He, S.Y.; Xin, X.-F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 2015, 6, 786. [Google Scholar] [CrossRef]
- Campos-Soriano, L.; García-Martínez, J.; Segundo, B.S. The arbuscular mycorrhizal symbiosis promotes the systemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection. Mol. Plant Pathol. 2012, 13, 579–592. [Google Scholar] [CrossRef]
- Pruitt, R.N.; Locci, F.; Wanke, F.; Zhang, L.; Saile, S.C.; Joe, A.; Karelina, D.; Hua, C.; Fröhlich, K.; Wan, W.-L. The EDS1–PAD4–ADR1 node mediates Arabidopsis pattern-triggered immunity. Nature 2021, 598, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Cao, M.; Xie, L.J.; Liang, X.T.; Zeng, R.S.; Su, Y.J.; Huang, J.H.; Wang, R.L.; Luo, S.M. Induction of DIMBOA accumulation and systemic defense responses as a mechanism of enhanced resistance of mycorrhizal corn (Zea mays L.) to sheath blight. Mycorrhiza 2011, 21, 721–731. [Google Scholar] [CrossRef]
- Cameron, D.D.; Neal, A.L.; van Wees, S.C.; Ton, J. Mycorrhiza-induced resistance: More than the sum of its parts? Trends Plant Sci. 2013, 18, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hao, Z.; Zhang, X.; Xie, W.; Chen, B. Arbuscular mycorrhizal fungi induced plant resistance against fusarium wilt in jasmonate biosynthesis defective mutant and wild type of tomato. J. Fungi 2022, 8, 422. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Guo, M.; Song, J.; Ma, Y.; Xu, Z. Signals in systemic acquired resistance of plants against microbial pathogens. Mol. Biol. Rep. 2021, 48, 3747–3759. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcón-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- He, X.; Anderson, J.C.; Pozo, O.d.; Gu, Y.Q.; Tang, X.; Martin, G.B. Silencing of subfamily I of protein phosphatase 2A catalytic subunits results in activation of plant defense responses and localized cell death. Plant J. 2004, 38, 563–577. [Google Scholar] [CrossRef]
- Liu, Y.; Shepherd, E.G.; Nelin, L.D. MAPK phosphatases—Regulating the immune response. Nat. Rev. Immunol. 2007, 7, 202–212. [Google Scholar] [CrossRef]
- Vilela, B.; Pagès, M.; Lumbreras, V. Regulation of MAPK signaling and cell death by MAPK phosphatase MKP2. Plant Signal. Behav. 2010, 5, 1497–1500. [Google Scholar] [CrossRef]
- Barman, A.; Ray, S.K. Protein phosphatase mediated responses in plant host-pathogen interactions. In Protein Phosphatases and Stress Management in Plants: Functional Genomic Perspective; Springer: Cham, Switzerland, 2020; pp. 289–330. [Google Scholar]
- Kachroo, A.; Liu, H.; Yuan, X.; Kurokawa, T.; Kachroo, P. Systemic acquired resistance-associated transport and metabolic regulation of salicylic acid and glycerol-3-phosphate. Essays Biochem. 2022, 66, 673–681. [Google Scholar] [PubMed]
- Zhao, S.; Li, M.; Ren, X.; Wang, C.; Sun, X.; Sun, M.; Yu, X.; Wang, X. Enhancement of broad-spectrum disease resistance in wheat through key genes involved in systemic acquired resistance. Front. Plant Sci. 2024, 15, 1355178. [Google Scholar] [CrossRef]
- Wang, Y.; Schuck, S.; Wu, J.; Yang, P.; Döring, A.-C.; Zeier, J.; Tsuda, K. A MPK3/6-WRKY33-ALD1-pipecolic acid regulatory loop contributes to systemic acquired resistance. Plant Cell 2018, 30, 2480–2494. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Sun, W.; Zheng, X.; Qiu, S.; Jiao, S.; Babilonia, K.; Koiwa, H.; He, P.; Shan, L.; Sun, W. Arabidopsis RNA polymerase II C-terminal domain phosphatase-like 1 targets mitogen-activated protein kinase cascades to suppress plant immunity. J. Integr. Plant Biol. 2023, 65, 2380–2394. [Google Scholar] [CrossRef] [PubMed]
- Sobol, G.; Chakraborty, J.; Martin, G.B.; Sessa, G. The emerging role of PP2C phosphatases in tomato immunity. Mol. Plant-Microbe Interact. 2022, 35, 737–747. [Google Scholar] [CrossRef]
- Seo, J.-K.; Kwon, S.-J.; Cho, W.K.; Choi, H.-S.; Kim, K.-H. Type 2C protein phosphatase is a key regulator of antiviral extreme resistance limiting virus spread. Sci. Rep. 2014, 4, 5905. [Google Scholar] [CrossRef]
- Chen, Y.-r.; Mao, Z.-j.; Li, Z.-w.; Fan, K. Research status and progress in structure and function of protein phosphatase 2C in plants. J. Zhejiang Univ. (Agric. Life Sci.) 2021, 47, 11–20. [Google Scholar]
- Diao, Z.; Yang, R.; Wang, Y.; Cui, J.; Li, J.; Wu, Q.; Zhang, Y.; Yu, X.; Gong, B.; Huang, Y. Functional screening of the Arabidopsis 2C protein phosphatases family identifies PP2C15 as a negative regulator of plant immunity by targeting BRI1-associated receptor kinase 1. Mol. Plant Pathol. 2024, 25, e13447. [Google Scholar] [CrossRef]
- Zhang, H.H.; Li, L.L.; He, Y.Q.; Qin, Q.Q.; Chen, C.H.; Wei, Z.Y.; Tan, X.X.; Xie, K.L.; Zhang, R.F.; Hong, G.J.; et al. Distinct modes of manipulation of rice auxin response factor OsARF17 by different plant RNA viruses for infection. Proc. Natl. Acad. Sci. USA 2020, 117, 9112–9121. [Google Scholar] [CrossRef]
- Yu, X.; Cui, X.; Wu, C.; Shi, S.; Yan, S. Salicylic acid inhibits gibberellin signaling through receptor interactions. Mol. Plant 2022, 15, 1759–1771. [Google Scholar] [CrossRef]
- Li, T.; Wang, Y.; Natran, A.; Zhang, Y.; Wang, H.; Du, K.; Qin, P.; Yuan, H.; Chen, W.; Tu, B. C-TERMINAL DOMAIN PHOSPHATASE-LIKE 3 contributes to GA-mediated growth and flowering by interaction with DELLA proteins. New Phytol. 2024, 242, 2555–2569. [Google Scholar] [CrossRef]
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, W.; Dai, L. The progress of the cross-talk among the signaling pathways of phytohormones in plant defense responses. Chin. Agric. Sci. Bull. 2013, 29, 153–157. [Google Scholar]
- Kaur, K.; Tripathy, M.K.; Pandey, G.K. Role of dual specificity phosphatase in stress and starch metabolism. In Protein Phosphatases and Stress Management in Plants: Functional Genomic Perspectiv; Springers: Berlin/Heidelberg, Germany, 2020; pp. 331–351. [Google Scholar]
- Hayek, S.; Gianinazzi-Pearson, V.; Gianinazzi, S.; Franken, P. Elucidating mechanisms of mycorrhiza-induced resistance against Thielaviopsis basicola via targeted transcript analysis of Petunia hybrida genes. Physiol. Mol. Plant Pathol. 2014, 88, 67–76. [Google Scholar] [CrossRef]
- Fernandes, T.R.; Segorbe, D.; Prusky, D.; Di Pietro, A. How alkalinization drives fungal pathogenicity. PLoS Pathog. 2017, 13, e1006621. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Niu, Y.; Huang, X.; He, Z.; Zhang, Q.; Meng, H.; Shi, H.; Feng, B.; Zhou, Y.; Zhang, J.; Lu, G. Phosphorylation of OsTGA5 by casein kinase II compromises its suppression of defense-related gene transcription in rice. Plant Cell 2022, 34, 3425–3442. [Google Scholar] [CrossRef]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ECM fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef]
- Guo, Y.; Li, F.; Li, Y.; Duan, T. Progress in the elucidation of the mechanisms of arbuscular mycorrhizal fungi in promotion of phosphorus uptake and utilization by plants. Pratacult. Sci. 2016, 10, 2379–2390. [Google Scholar]
- Uroz, S.; Heinonsalo, J. Degradation of N-acyl homoserine lactone quorum sensing signal molecules by forest root-associated fungi. FEMS Microbiol. Ecol. 2008, 65, 271–278. [Google Scholar] [CrossRef]
- Caravaca, F.; Maboreke, H.; Kurth, F.; Herrmann, S.; Tarkka, M.T.; Ruess, L. Synergists and antagonists in the rhizosphere modulate microbial communities and growth of Quercus robur L. Soil Biol. Biochem. 2015, 82, 65–73. [Google Scholar] [CrossRef]
- Berrios, L.; Yeam, J.; Holm, L.; Robinson, W.; Pellitier, P.T.; Chin, M.L.; Henkel, T.W.; Peay, K.G. Positive interactions between mycorrhizal fungi and bacteria are widespread and benefit plant growth. Curr. Biol. 2023, 33, 2878–2887.e2874. [Google Scholar] [CrossRef]
- Huang, J.-H.; Zeng, R.-S.; Luo, S.-M. On disease resistance of maize toward sheath blight induced by arbuscular mycorrhizal fungi. Chin. J. Eco-Agric. 2006, 14, 167–169. [Google Scholar]
- Yang, M.; Zou, J.; Liu, C.; Xiao, Y.; Zhang, X.; Yan, L.; Ye, L.; Tang, P.; Li, X. Chinese white truffles shape the ectomycorrhizal microbial communities of Corylus avellana. Ann. Microbiol. 2019, 69, 553–565. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Yang, M.; Yan, L.; Kang, Z.; Xiao, Y.; Tang, P.; Ye, L.; Zhang, B.; Zou, J. Tuber borchii shapes the ectomycorrhizosphere microbial communities of Corylus avellana. Mycobiology 2019, 47, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.; Zou, J.; Huang, Y.; Zhang, X.; Ye, L.; Zhang, B.; Li, X. Tuber melanosporum shapes nirS-type denitrifying and ammonia-oxidizing bacterial communities in Carya illinoinensis ectomycorrhizosphere soils. PeerJ 2020, 8, e9457. [Google Scholar] [CrossRef]
- Zhang, X.; Ye, L.; Kang, Z.; Zou, J.; Li, X. Mycorrhization of Quercus acutissima with Chinese black truffle significantly altered the host physiology and root-associated microbiomes. PeerJ 2019, 7, e6421. [Google Scholar] [CrossRef]
- Nurlaeny, N.; Marschner, H.; George, E. Effects of liming and mycorrhizal colonization on soil phosphate depletion and phosphate uptake by maize (Zea mays L.) and soybean (Glycine max L.) grown in two tropical acid soils. Plant Soil 1996, 181, 275–285. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Carvalhais, L.C.; Percy, C.D.; Prakash Verma, J.; Schenk, P.M.; Singh, B.K. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytol. 2021, 229, 2873–2885. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of arbuscular mycorrhizal fungi, phosphate–solubilizing bacteria, and silicon to P uptake by plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, Z.; Li, X.; Tan, J.; Liu, F.; Wu, J. Mycorrhizal fungi alter root exudation to cultivate a beneficial microbiome for plant growth. Funct. Ecol. 2023, 37, 664–675. [Google Scholar] [CrossRef]
- Seipke, R.F.; Kaltenpoth, M.; Hutchings, M.I. Streptomyces as symbionts: An emerging and widespread theme? FEMS Microbiol. Rev. 2012, 36, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Lehr, N.A.; Adomas, A.; Asiegbu, F.O.; Hampp, R.; Tarkka, M.T. WS-5995 B, an antifungal agent inducing differential gene expression in the conifer pathogen Heterobasidion annosum but not in Heterobasidion abietinum. Appl. Microbiol. Biotechnol. 2009, 85, 347–358. [Google Scholar] [CrossRef]
- Riedlinger, J.; Schrey, S.D.; Tarkka, M.T.; Hampp, R.; Kapur, M.; Fiedler, H.-P. Auxofuran, a novel metabolite that stimulates the growth of fly agaric, is produced by the mycorrhiza helper bacterium Streptomyces strain AcH 505. Appl. Environ. Microbiol. 2006, 72, 3550–3557. [Google Scholar] [CrossRef]
- Duponnois, R.; Garbaye, J. Some mechanisms involved in growth stimulation of ectomycorrhizal fungi by bacteria. Can. J. Bot. 1990, 68, 2148–2152. [Google Scholar] [CrossRef]
- Wang, X.; Ding, T.; Li, Y.; Guo, Y.; Li, Y.; Duan, T. Dual inoculation of alfalfa (Medicago sativa L.) with Funnelliformis mosseae and Sinorhizobium medicae can reduce Fusarium wilt. J. Appl. Microbiol. 2020, 129, 665–679. [Google Scholar] [CrossRef]
- Sanchez, L.; Courteaux, B.; Hubert, J.; Kauffmann, S.; Renault, J.-H.; Clément, C.; Baillieul, F.; Dorey, S. Rhamnolipids elicit defense responses and induce disease resistance against biotrophic, hemibiotrophic, and necrotrophic pathogens that require different signaling pathways in Arabidopsis and highlight a central role for salicylic acid. Plant Physiol. 2012, 160, 1630–1641. [Google Scholar] [CrossRef]
- Jain, A.; Das, S. Insight into the interaction between plants and associated fluorescent Pseudomonas spp. Int. J. Agron. 2016, 2016, 4269010. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Garbaye, J.; Tarkka, M. The mycorrhiza helper bacteria revisited. New Phytol. 2007, 176, 22–36. [Google Scholar] [CrossRef]
- Founoune, H.; Duponnois, R.; Ba, A.; Sall, S.; Branget, I.; Lorquin, J.; Neyra, M.; Chotte, J.-L. Mycorrhiza helper bacteria stimulate ectomycorrhizal symbiosis of Acacia holosericea with Pisolithus alba. New Phytol. 2002, 153, 81–89. [Google Scholar] [CrossRef]
- Nishad, R.; Ahmed, T.; Rahman, V.J.; Kareem, A. Modulation of plant defense system in response to microbial interactions. Front. Microbiol. 2020, 11, 1298. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zou, J.; Kang, Z.; Zhang, X.; Penttinen, P.; Li, X. Effects of truffle inoculation on a nursery culture substrate environment and seedling of Carya illinoinensis. Fungal Biol. 2021, 125, 576–584. [Google Scholar] [CrossRef]
- Aguk, J.; Karanja, N.; Schulte-Geldermann, E.; Bruns, C.; Kinyua, Z.; Parker, M. Control of bacterial wilt (Ralstonia solanacearum) in potato (Solanum tuberosum) using rhizobacteria and arbuscular mycorrhiza fungi. Afr. J. Food Agric. Nutr. Dev. 2018, 18, 13371–13387. [Google Scholar]
- Gao, P.; Li, Y.; Guo, Y.; Duan, T. Co-inoculation of lucerne (Medicago sativa) with an AM fungus and a rhizobium reduces occurrence of spring black stem and leaf spot caused by Phoma medicaginis. Crop Pasture Sci. 2018, 69, 933–943. [Google Scholar] [CrossRef]
- Chandanie, W.; Kubota, M.; Hyakumachi, M. Interactions between plant growth promoting fungi and arbuscular mycorrhizal fungus Glomus mosseae and induction of systemic resistance to anthracnose disease in cucumber. Plant Soil 2006, 286, 209–217. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Roldán, A.; Pascual, J.A. Interaction between arbuscular mycorrhizal fungi and Trichoderma harzianum under conventional and low input fertilization field condition in melon crops: Growth response and Fusarium wilt biocontrol. Appl. Soil Ecol. 2011, 47, 98–105. [Google Scholar] [CrossRef]
- Guo, Y.; Gao, P.; Li, F.; Duan, T. Effects of AM fungi and grass endophytes on perennial ryegrass Bipolaris sorokiniana leaf spot disease under limited soil nutrients. Eur. J. Plant Pathol. 2019, 154, 659–671. [Google Scholar] [CrossRef]
- Nafady, N.A.; Hashem, M.; Hassan, E.A.; Ahmed, H.A.; Alamri, S.A. The combined effect of arbuscular mycorrhizae and plant-growth-promoting yeast improves sunflower defense against Macrophomina phaseolina diseases. Biol. Control 2019, 138, 104049. [Google Scholar] [CrossRef]
- Velivelli, S.L.; Lojan, P.; Cranenbrouck, S.; de Boulois, H.D.; Suarez, J.P.; Declerck, S.; Franco, J.; Prestwich, B.D. The induction of Ethylene response factor 3 (ERF3) in potato as a result of co-inoculation with Pseudomonas sp. R41805 and Rhizophagus irregularis MUCL 41833–a possible role in plant defense. Plant Signal. Behav. 2015, 10, e988076. [Google Scholar] [CrossRef]
- Behn, O. Influence of Pseudomonas fluorescens and arbuscular mycorrhiza on the growth, yield, quality and resistance of wheat infected with Gaeumannomyces graminis/Einfluss von Pseudomonas fluorescens und Mykorrhiza auf Wachstum, Ertrag, Qualität und Resistenz von Weizen bei Befall mit Gaeumannomyces graminis. J. Plant Dis. Prot. 2008, 115, 4–8. [Google Scholar]
- Liu, R.; Dai, M.; Wu, X.; Li, M.; Liu, X. Suppression of the root-knot nematode [Meloidogyne incognita (Kofoid & White) Chitwood] on tomato by dual inoculation with arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria. Mycorrhiza 2012, 22, 289–296. [Google Scholar] [PubMed]
- Wang, Y.-H.; Dai, Y.; Kong, W.-L.; Zhu, M.-L.; Wu, X.-Q. Improvement of sphaeropsis Shoot Blight Disease resistance by applying the ectomycorrhizal fungus Hymenochaete sp. Rl and mycorrhizal helper Bacterium Bacillus pumilus HR10 to Pinus thunbergii. Phytopathology® 2022, 112, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.; Toro, M.; Memenza-Zegarra, M.; Zúñiga-Dávila, D. Control of White Rot Caused by Sclerotinia sclerotiorum in Strawberry Using Arbuscular Mycorrhizae and Plant-Growth-Promoting Bacteria. Sustainability 2023, 15, 2901. [Google Scholar] [CrossRef]
- Abdel-Fattah, G.; Mohamedin, A. Interactions between a vesicular-arbuscular mycorrhizal fungus (Glomus intraradices) and Streptomyces coelicolor and their effects on sorghum plants grown in soil amended with chitin of brawn scales. Biol. Fertil. Soils 2000, 32, 401–409. [Google Scholar] [CrossRef]
- Banerjee, A.; Sanyal, S.; Sen, S. Soil phosphatase activity of agricultural land: A possible index of soil fertility. Agric. Sci. Res. J. 2012, 2, 412–419. [Google Scholar]
- Cavagnaro, T.R.; Bender, S.F.; Asghari, H.R.; van der Heijden, M.G. The role of arbuscular mycorrhizas in reducing soil nutrient loss. Trends Plant Sci. 2015, 20, 283–290. [Google Scholar] [CrossRef]
- Hunter, P.J.; Teakle, G.R.; Bending, G.D. Root traits and microbial community interactions in relation to phosphorus availability and acquisition, with particular reference to Brassica. Front. Plant Sci. 2014, 5, 27. [Google Scholar] [CrossRef]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of Arbuscular Mycorrhizal Fungi on Soil Fertility: Contribution in the Improvement of Physical, Chemical, and Biological Properties of the Soil. Front. Fungal Biol. 2022, 3, 723892. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, 1089. [Google Scholar] [CrossRef]
- Cornejo, P.; Borie, F.; Rubio, R.; Azcón, R. Influence of nitrogen source on the viability, functionality and persistence of Glomus etunicatum fungal propagules in an Andisol. Appl. Soil Ecol. 2007, 35, 423–431. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).