
Metabolic and Antioxidant Responses of Different Control Methods to the Interaction of Sorghum sudangrass hybrids-Colletotrichum boninense

Abstract
:1. Introduction
2. Results
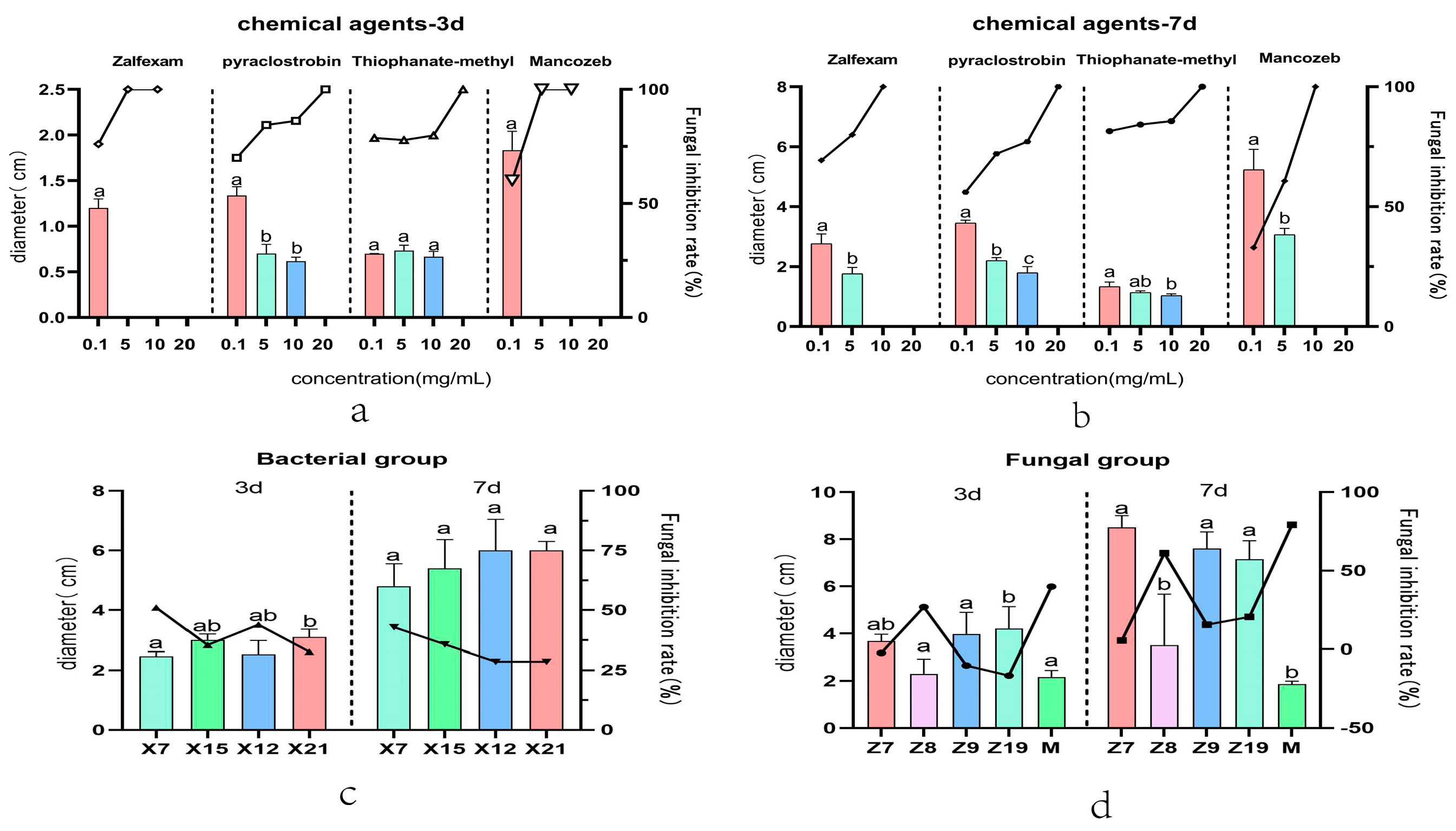
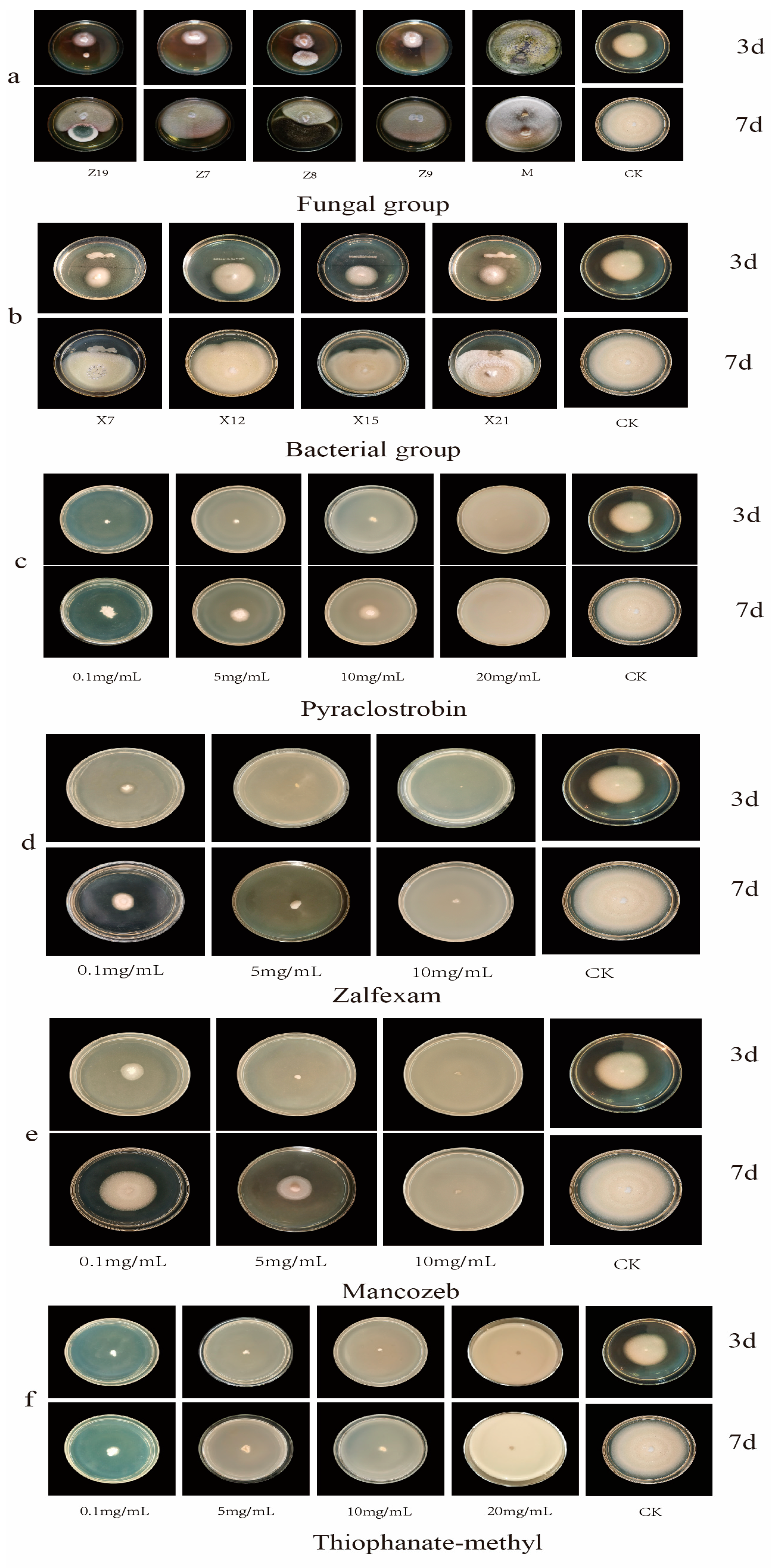
2.1. Initial Screening of Chemical Fungicides
2.2. Screening Results of Bacterial Biocides
2.3. Screening Results of Fungicides
2.4. Measurement of Biomass of Different Treatment Groups Treating Sorghum sudangrass hybrids Seedlings
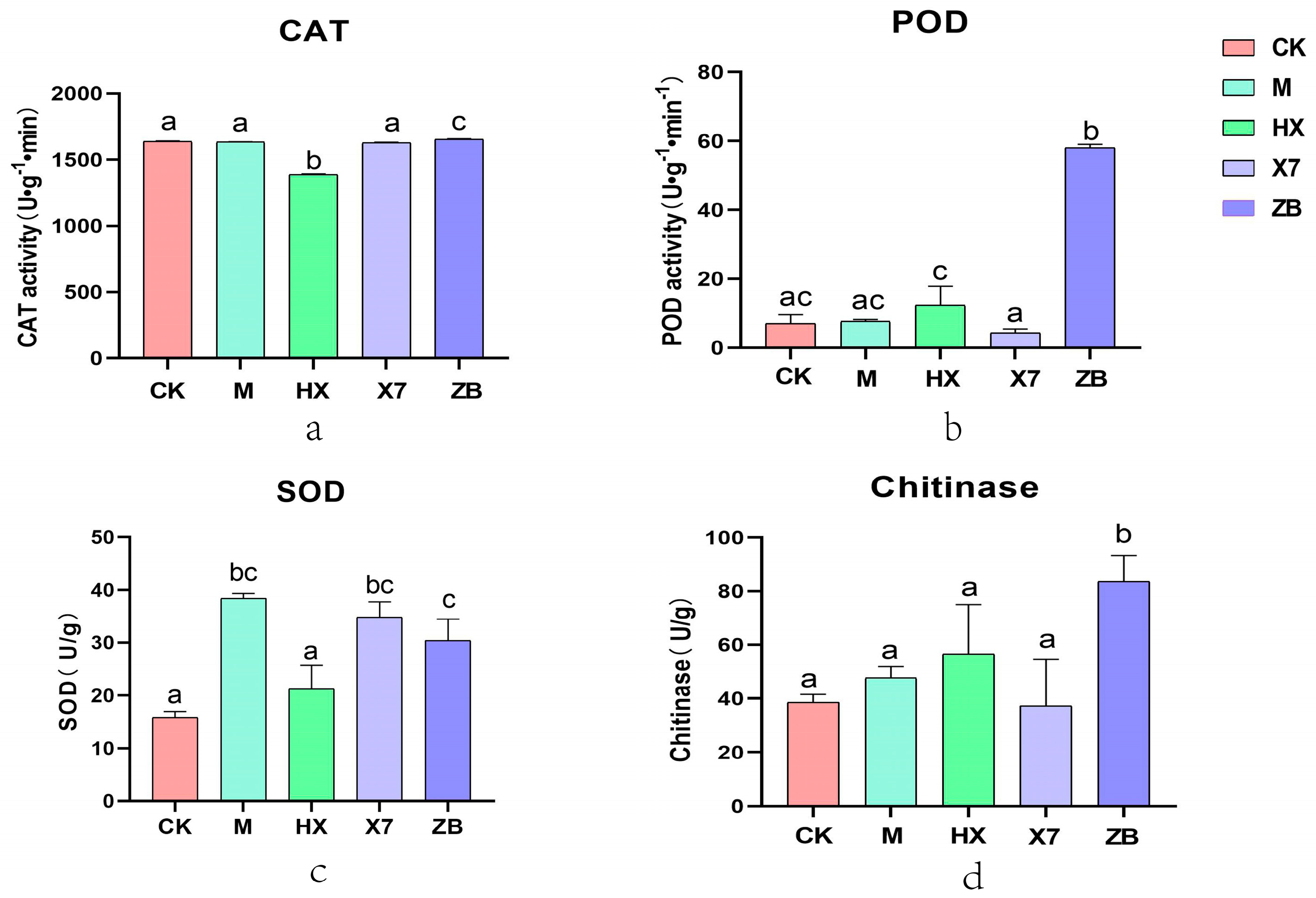
2.5. Analysis of Enzyme Activities in Different Treatment Groups of Treated Sorghum sudangrass hybrids Seedlings
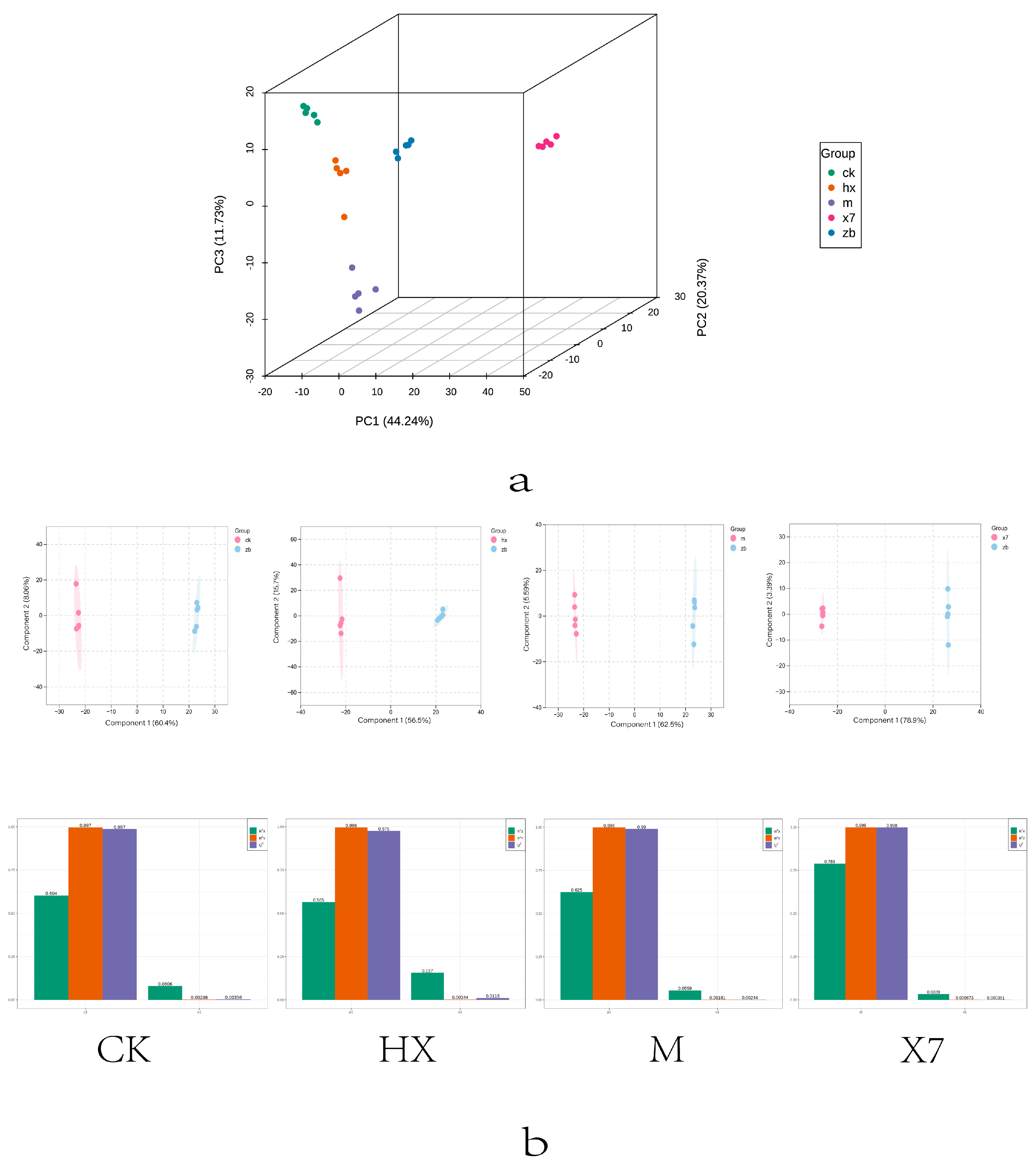
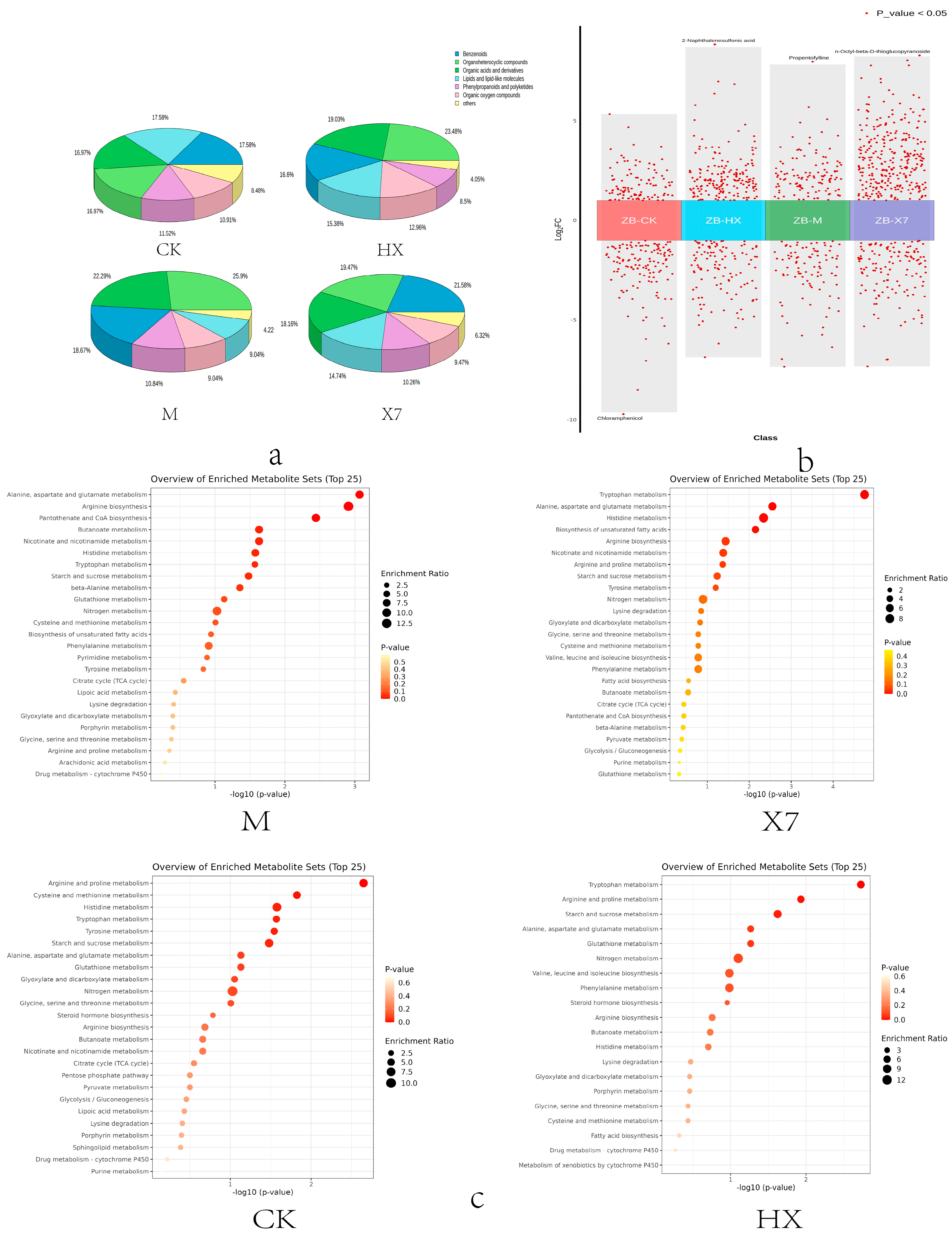
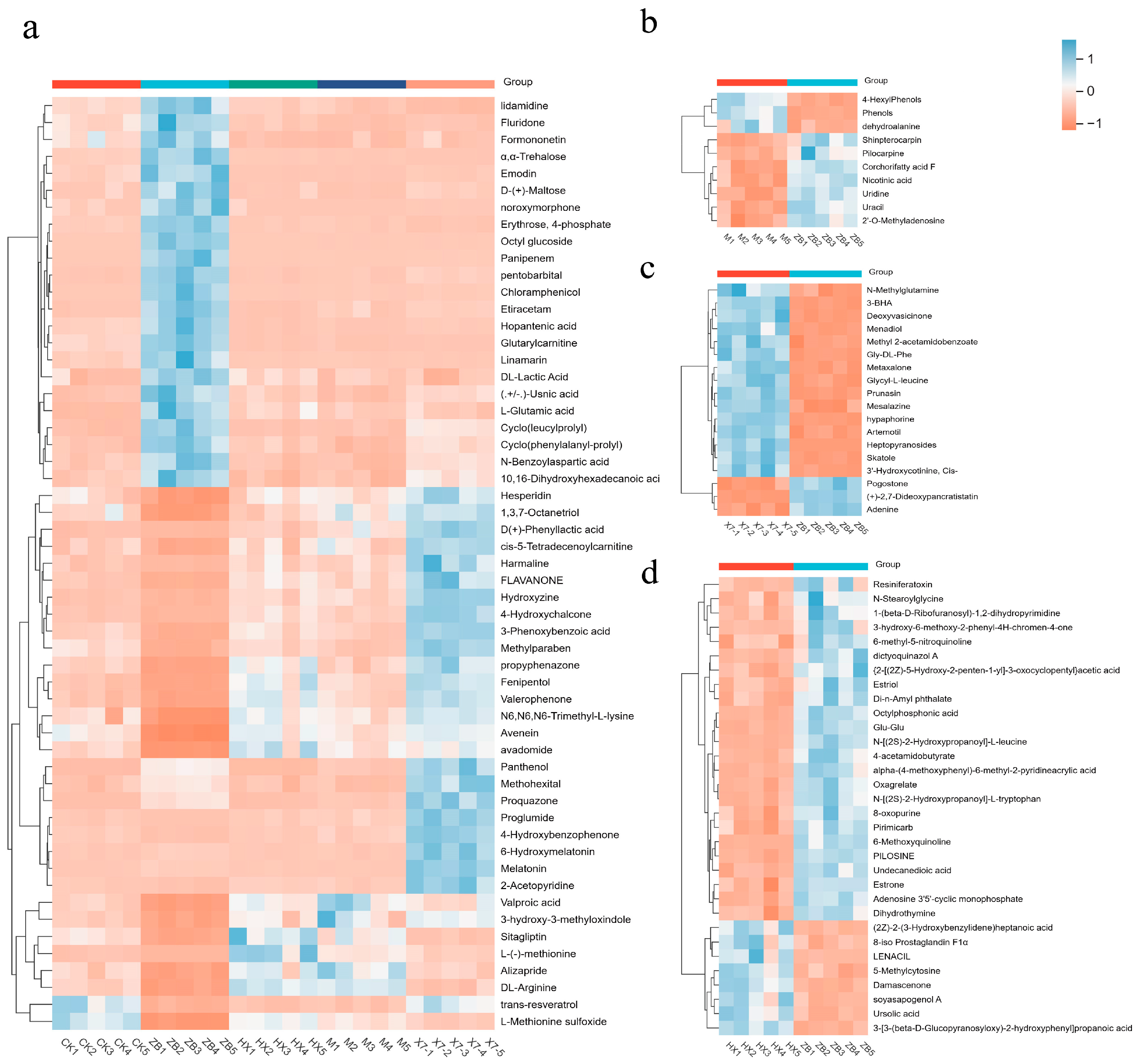
2.6. Metabolomic Analysis
3. Discussion
3.1. Plant Fungal Disease Control Tools Are Diverse
3.2. Antioxidant Enzymes Have an Important Role in Plant Response to Pathogen Stress
3.3. The Influence of Different Treatment Methods on the Important Metabolites of Sorghum sudangrass hybirds
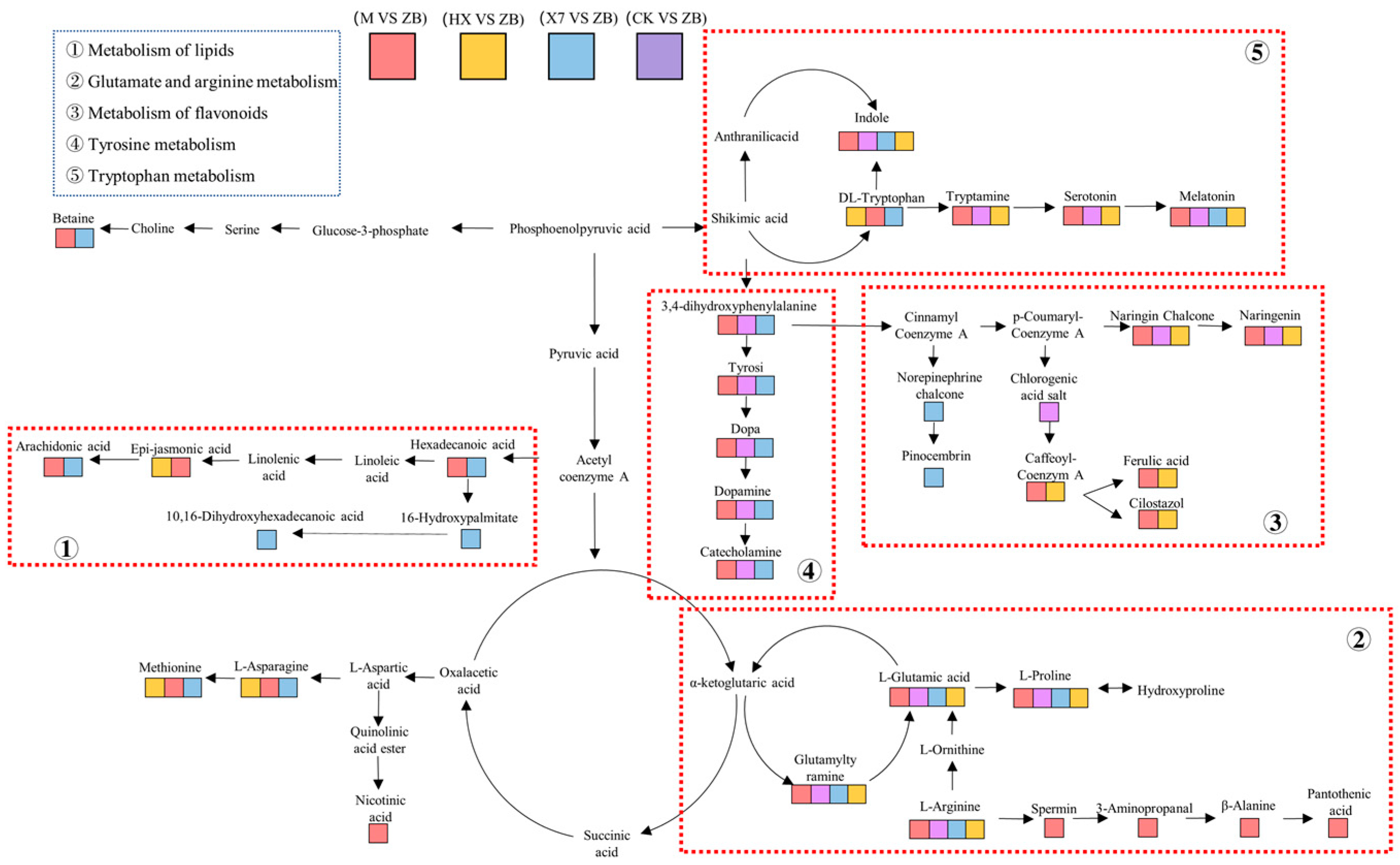
3.4. Amino Acid Metabolism, Secondary Metabolism of Flavonoids and the Jasmonic Acid Pathway May Exert Disease-Resistant Effects
4. Materials and Methods
4.1. Trial Material
4.2. Screening of Chemical Fungicides
4.3. Screening of Biological Fungicides
4.4. Potting Trials
4.5. Measurements of Plant Physiological Parameters
4.6. Antioxidant Enzyme Activity Assay
4.7. Sample Preparation of Non-Targeted Metabolome of Sorghum sudangrass hybrids Leaves
4.8. Data Analysis
5. Conclusions
- The best chemical inhibitor was zalfexam, with a minimum inhibitory concentration of 10 mg/mL. Among the choices of biocides were the highest inhibition rate of Bacillus velezensis, where X7 could reach 51.85%, and the highest inhibition rate of Trichoderma harzianum, which was 80.56%.
- Trichoderma harzianum-treated group M showed an increase in both POD and chitinase enzyme activities, zalfexam-treated group HX showed an increase in SOD, POD, and chitinase enzyme activities to defend against the aggression, and the Bacillus velezensis-treated group X7 only showed a significant increase in POD enzymes. This study concludes that all three groups exhibited antioxidant capacities; however, SOD was identified as the primary free radical scavenger, with a significant enhancement in activity observed in group M. Therefore, it can be concluded that group M had a more pronounced influence on antioxidant enzyme properties.
- The common pathway of the four groups CK vs. ZB, X7 vs. ZB, M vs. ZB, and HX vs. ZB, is glutamate–arginine and tryptophan metabolism, and three of the treatment groups (X7, M, and HX groups) are resistant to stress. The X7 group reduces reactive oxygen species and improves plant disease resistance through tyrosine metabolism, pinocembrin production, and AA up-regulation. The M group responds to ROS and bolster plant resistance via its glutathione–ascorbic acid and tyrosine metabolism, along with the up-regulation of jasmonic acid-salicylic acid (JA-SA) and the production of ferulic acid and cilostazol, which collectively contribute to stress mitigation by eliminating ROS and improving plant disease resistance. In contrast, the HX group primarily increases jasmonic acid content and the production of ferulic acid and cilostazol to withstand stress.
- This study demonstrates that the combination of bacterial inhibition observed during plate culture, plant growth indices, alterations in antioxidant enzyme activities, and key metabolic pathways indicates that group M, treated with Trichoderma harzianum, is more effective for disease control and enhances plant resistance to stress.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Traetments | 3 d Diameter | Anti-Fungal Rate | 7 d Diameter | Anti-Fungal Rate | 15d Diameter | Anti-Fungal Rate |
|---|---|---|---|---|---|---|---|
| zalfexam | 0.1 mg/mL | 1.200 ± 0.1000 a | 74–76% | 2.767 ± 0.3215 a | 66.11–72.55% | - | - |
| 5 mg/mL | 0.000 b | 100% | 1.767 ± 0.2082 b | 77.35–82.65% | - | - | |
| 10 mg/mL | 0.000 b | 100% | 0.000 c | 100% | - | - | |
| pyraclostrobin | 0.1 mg/mL | 1.337 ± 0.09609 a | 68.90–71.71% | 3.457 ± 0.09292 a | 54.53–57.55% | - | - |
| 5 mg/mL | 0.7000 ± 0.1000 b | 86.22–82.74% | 2.2000 ± 0.1000 b | 71.35–72.73% | - | - | |
| 10 mg/mL | 0.6167 ± 0.04509 b | 84.76–87.7% | 1.800 ± 0.2000 c | 75.15–79.15% | - | - | |
| 20 mg/mL | 0.000 c | 100% | 0.000 d | 100% | - | - | |
| thiophanate- methyi | 0.1 mg/mL | 0.7000 ± 0.000 a | 78.79% | 1.333 ± 0.1528 a | 79.36–83.60% | - | - |
| 5 mg/mL | 0.7333 ± 0.05774 a | 76.03–79.53% | 1.133 ± 0.05774 ab | 83.46–85.06% | - | - | |
| 10 mg/mL | 0.6667 ± 0.05774 a | 78.05–81.54% | 1.033 ± 0.05774 b | 84.85–86.45% | - | - | |
| 20 mg/mL | 0.000 b | 100% | 0.000 c | 100% | - | - | |
| mancozeb | 0.1 mg/mL | 1.833 ± 0.2082 a | 55.62–64.68% | 5.233 ± 0.6807 a | 24.19–41.66% | - | - |
| 5 mg/mL | 0.000 b | 100% | 3.067 ± 0.2082 b | 58.01–63.35% | - | - | |
| 10 mg/mL | 0.000 b | 100% | 0.000 c | 100% | - | - | |
| X7 | - | 2.300 ± 0.2000 a | 46.80–55.32% | 4.800 ± 0.7550 a | 33.87–51.85% | 6.367 ± 0.2082 a | 26.944–31.576% |
| X15 | - | 3.033 ± 0.1528 ab | 32.21–38.71% | 5.400 ± 0.9644 a | 24.24–47.2% | 7.700 ± 0.3606 b | 7.307–11.939% |
| X12 | - | 2.633 ± 0.5132 ab | 33.05–54.89% | 6.000 ± 1.044 a | 16.14–41% | 8.167 ± 0.1528 c | 4.169–5.45% |
| X21 | - | 3.167 ± 0.2517 b | 27.26–37.98% | 6.000 ± 0.3000 a | 25–32.14% | 8.000 ± 0.1000 d | 4.44–6.66% |
| Z7 | - | 3.683 ± 0.2843 ab | −10.1–5.5% | 8.500 ± 0.5000 a | 0–1.1% | - | - |
| Z8 | - | 2.300 ± 0.6245 a | 13.72–39.98% | 3.500 ± 2.166 b | 37.05–85.17% | - | - |
| Z9 | - | 3.967 ± 0.9238 a | −35.85–15.47% | 7.603 ± 0.7071 a | 7.66–23.38% | - | - |
| Z19 | - | 4.20 0± 0.9341 b | −42.62–9.28% | 7.150 ± 0.7858 a | 11.86–29.26% | - | - |
| Trichoderma harzianum | - | 2.167 ± 0.2754 a | 32.17–47.45% | 1.867 ± 0.1258 b | 77.86–80.65% | - | - |

References
- Ding, Q.; Xie, G.; Zhang, H.; Ban, Q.; Lu, X.; Wang, X.; Huang, L. Comparative experiment on the production performance of four Sorghum bicolor × S. sudanense cultivars in the Chengdu Plain. Pratacultural Sci. 2021, 38, 2421–2428. [Google Scholar]
- Zhao, Y. QTL Mapping for Important Traits Such as Yield in Sorghum-Sudangrass. Master’s Thesis, Inner Mongolia University, Hohhot, China, 2024. [Google Scholar]
- Li, J.; Xu, J.; Wang, H.; Wu, C.; Zheng, J.; Zhang, C.; Han, Y. First Report of Fungal Pathogens Causing Leaf Spot on Sorghum–Sudangrass Hybrids and Their Interactions with Plants. Plants 2023, 12, 3091. [Google Scholar] [CrossRef]
- Damm, U.; Cannon, P.F.; Woudenberg, J.H.C.; Johnston, P.R.; Weir, B.S.; Tan, Y.P.; Shivas, R.G.; Crous, P.W. The Colletotrichum boninense Species Complex. Stud. Mycol. 2012, 73, 1–36. [Google Scholar] [CrossRef]
- Ding, H.-X.; Dong, W.-P.; Mo, W.-D.; Peng, L.-J.; Liu, Z.-Y. First Report of Colletotrichum boninense Causing Anthracnose on Rosa chinensis in China. Plant Dis. 2021, 105, 2717. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.L.; Maciel-Zambolim, E.; Zambolim, L.; Lelis, D.T.; Caixeta, E.T.; Lopes, U.P.; Pereira, O.L. Colletotrichum boninense Causing Anthracnose on Coffee Trees in Brazil. Plant Dis. 2013, 97, 1255. [Google Scholar] [CrossRef]
- Ding, H.-X.; Cheng, H.-H.; Li, J.-Y.; Liu, Z.-Y.; Peng, L.-J. First Report of Colletotrichum boninense Causing Anthracnose on Goldthread (Coptis chinensis) in China. Plant Dis. 2020, 104, 1538. [Google Scholar] [CrossRef]
- Li, T.; Tang, H. An analysis of the control of pests and diseases and processing and utilisation of Sorghum sudangrass hybrids. Chin. Livest. Poult. Breed. 2018, 14, 36–37. [Google Scholar]
- Xu, F. Occurrence and control of plant pests and diseases. China Agric. Inf. 2015, 72. [Google Scholar]
- Ahamad, L.; Zaidi, R.K. Seed Priming Through Fungicides, Biocontrol Agents and Botanicals as Effective Method for Controlling Spot Blotch Pathogen, Bipolaris sorokiniana in Barley. Gesunde Pflanz. 2022, 74, 501–510. [Google Scholar] [CrossRef]
- Ren, S.; Ma, F.; Li, M.; Fan, Y.; Zhu, H.; Zhu, K.; Guo, Z. Field efficacy test of 20% pyraclostrobin-pentaconazole suspension for the control of powdery mildew on chrysanthemums. J. Seed Ind. Guide 2023, 46–48. [Google Scholar]
- McLaughlin, M.S.; Roy, M.; Abbasi, P.A.; Carisse, O.; Yurgel, S.N.; Ali, S. Why Do We Need Alternative Methods for Fungal Disease Management in Plants? Plants 2023, 12, 3822. [Google Scholar] [CrossRef]
- LIin, M. Research on biological control techniques in plant disease control. J. Agric. Catastrophology 2023, 13, 13–15. [Google Scholar]
- Cenobio-Galindo, A.d.J.; Hernández-Fuentes, A.D.; González-Lemus, U.; Zaldívar-Ortega, A.K.; González-Montiel, L.; Madariaga-Navarrete, A.; Hernández-Soto, I. Biofungicides Based on Plant Extracts: On the Road to Organic Farming. Int. J. Mol. Sci. 2024, 25, 6879. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, B.; Verma, S.; Pal, J.; Mukesh; Akanksha; Chauhan, P. Unveiling the Biocontrol Potential of Trichoderma. Eur. J. Plant Pathol. 2023, 167, 569–591. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, T.; Li, R.; Li, M.; Lian, H. Effect of Trichoderma pseudokoningii on the growth, antioxidant system and wilt control of cucumber seedlings. Agric. Res. Arid. Areas 2022, 40, 72–81+107. [Google Scholar]
- Guo, Z.; Zhang, J.; Liu, Z.; Li, Y.; Li, M.; Meng, Q.; Yang, Z.; Luo, Y.; Zhang, Q.; Yan, M. Trichoderma harzianum Prevents Red Kidney Bean Root Rot by Increasing Plant Antioxidant Enzyme Activity and Regulating the Rhizosphere Microbial Community. Front. Microbiol. 2024, 15, 1348680. [Google Scholar] [CrossRef]
- Luo, S.; Tian, C.; Zhang, H.; Yao, Z.; Guan, Z.; Li, Y.; Zhang, J.; Song, Y. Isolation and Identification of Biocontrol Bacteria against Atractylodes chinensis Root Rot and Their Effects. Microorganisms 2023, 11, 2384. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Wang, S.; Li, X.; Li, X.; Cai, J.; Zhao, L.; Wang, J.; Zeng, J.; Qin, Y.; Xiong, X.; et al. Biological Control of Potato Common Scab and Growth Promotion of Potato by Bacillus velezensis Y6. Front. Microbiol. 2023, 14, 1295107. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Fu, M.; Xu, M.; Chen, X.; Qiu, J.; Wang, F.; Yan, R.; Wang, J.; Zhao, S.; Xin, X.; et al. Application of Antagonist Bacillus amyloliquefaciens NCPSJ7 against Botrytis cinerea in Postharvest Red Globe Grapes. Food Sci. Nutr. 2020, 8, 1499–1508. [Google Scholar] [CrossRef]
- Vaghela, B.; Vashi, R.; Rajput, K.; Joshi, R. Plant Chitinases and Their Role in Plant Defense: A Comprehensive Review. Enzym. Microb. Technol. 2022, 159, 110055. [Google Scholar] [CrossRef]
- Pang, Z.; Mao, X.; Zhou, S.; Yu, S.; Liu, G.; Lu, C.; Wan, J.; Hu, L.; Xu, P. Microbiota-Mediated Nitrogen Fixation and Microhabitat Homeostasis in Aerial Root-Mucilage. Microbiome 2023, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Ingrisano, R.; Tosato, E.; Trost, P.; Gurrieri, L.; Sparla, F. Proline, Cysteine and Branched-Chain Amino Acids in Abiotic Stress Response of Land Plants and Microalgae. Plants 2023, 12, 3410. [Google Scholar] [CrossRef]
- Al Aboody, M.S.; Mickymaray, S. Anti-Fungal Efficacy and Mechanisms of Flavonoids. Antibiotics 2020, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.; Choudhary, A.K.; Lekharam; Kumawat, H. In Vitro and In Vivo Evaluation of Fungicides, against Colletotrichum lindemuthianum Causing Anthracnose of Mungbean. Int. J. Plant Soil. Sci. 2022, 34, 311–316. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, T. Strong Opponent of Walnut Anthracnose—Bacillus velezensis and Its Transcriptome Analysis. Microorganisms 2023, 11, 1885. [Google Scholar] [CrossRef]
- Yeo, Y.J.; Park, A.R.; Vuong, B.S.; Kim, J.-C. Biocontrol of Fusarium Head Blight in Rice Using Bacillus velezensis JCK-7158. Front. Microbiol. 2024, 15, 1358689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kong, N.; Liu, B.; Yang, Y.; Li, C.; Qi, J.; Ma, Y.; Ji, S.; Liu, Z. Biocontrol Potential of Trichoderma harzianum CGMCC20739 (Tha739) against Postharvest Bitter Rot of Apples. Microbiol. Res. 2022, 265, 127182. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.-Q.; Wang, M.; Sun, J.; Gao, J.-X.; Chen, J. Effect of Trichoderma harzianum on Maize Rhizosphere Microbiome and Biocontrol of Fusarium Stalk Rot. Sci. Rep. 2017, 7, 1771. [Google Scholar] [CrossRef]
- Elad, Y.; Kapat, A. The Role of Trichoderma harzianum Protease in the Biocontrol of Botrytis cinerea. Eur. J. Plant Pathol. 1999, 105, 177–189. [Google Scholar] [CrossRef]
- Bisht, A.; Garg, N. AMF Species Improve Yielding Potential of Cd Stressed Pigeonpea Plants by Modulating Sucrose-Starch Metabolism, Nutrients Acquisition and Soil Microbial Enzymatic Activities. Plant Growth Regul. 2022, 96, 409–430. [Google Scholar] [CrossRef]
- Fang, S.; Tao, Y.; Zhang, Y.; Kong, F.; Wang, Y. Effects of Metalaxyl Enantiomers Stress on Root Activity and Leaf Antioxidant Enzyme Activities in Tobacco Seedlings. Chirality 2018, 30, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Advances in Metal-Induced Oxidative Stress and Human Disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Cadmium Stress Alters the Redox Reaction and Hormone Balance in Oilseed Rape (Brassica napus L.) Leaves|Environmental Science and Pollution Research. Available online: https://link.springer.com/article/10.1007/s11356-015-5640-y (accessed on 21 July 2024).
- Tian, S.; Chen, Y.; Cai, Z.; Wang, Q.; Zhang, J.; Liu, Z.; Li, Y.; Zhao, X. Effects of Harpin and Carbendazim on Antioxidant Accumulation in Young Jujube Leaves. Open Chem. 2023, 21, 20220284. [Google Scholar] [CrossRef]
- Zehra, A.; Meena, M.; Dubey, M.K.; Aamir, M.; Upadhyay, R.S. Synergistic Effects of Plant Defense Elicitors and Trichoderma harzianum on Enhanced Induction of Antioxidant Defense System in Tomato against Fusarium Wilt Disease. Bot. Stud. 2017, 58, 44. [Google Scholar] [CrossRef]
- Zhang, F.; Ruan, X.; Wang, X.; Liu, Z.; Hu, L.; Li, C. Overexpression of a Chitinase Gene from Trichoderma asperellum Increases Disease Resistance in Transgenic Soybean. Appl. Biochem. Biotechnol. 2016, 180, 1542–1558. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Z.; Lu, B.; Quan, X.; Zhao, G.; Zhang, Z.; Liu, W.; Tian, Y. Antagonistic Potential of Trichoderma as a Biocontrol Agent against Sclerotinia asari. Front. Microbiol. 2022, 13, 997050. [Google Scholar] [CrossRef]
- Tančić-Živanov, S.; Medić-Pap, S.; Danojević, D.; Prvulović, D. Effect of Trichoderma Spp. on Growth Promotion and Antioxidative Activity of Pepper Seedlings. Braz. Arch. Biol. Technol. 2020, 63, e20180659. [Google Scholar] [CrossRef]
- Arias Padró, M.D.; Caboni, E.; Salazar Morin, K.A.; Meraz Mercado, M.A.; Olalde-Portugal, V. Effect of Bacillus subtilis on Antioxidant Enzyme Activities in Tomato Grafting. PeerJ 2021, 9, e10984. [Google Scholar] [CrossRef]
- Rais, A.; Jabeen, Z.; Shair, F.; Hafeez, F.Y.; Hassan, M.N. Bacillus Spp., a Bio-Control Agent Enhances the Activity of Antioxidant Defense Enzymes in Rice against Pyricularia oryzae. PLoS ONE 2017, 12, e0187412. [Google Scholar] [CrossRef]
- Ma, X.; Wang, J. Formononetin: A Pathway to Protect Neurons. Front. Integr. Neurosci. 2022, 16, 908378. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Gao, J.; Pang, X.; Chen, A.; Wang, Y. Molecular Mechanisms of Action of Emodin: As an Anti-Cardiovascular Disease Drug. Front. Pharmacol. 2020, 11, 559607. [Google Scholar] [CrossRef] [PubMed]
- Nozaka, A.; Nishiwaki, A.; Nagashima, Y.; Endo, S.; Kuroki, M.; Nakajima, M.; Narukawa, M.; Kamisuki, S.; Arazoe, T.; Taguchi, H.; et al. Chloramphenicol Inhibits Eukaryotic Ser/Thr Phosphatase and Infection-Specific Cell Differentiation in the Rice Blast Fungus. Sci. Rep. 2019, 9, 9283. [Google Scholar] [CrossRef]
- Paguirigan, J.A.; Liu, R.; Im, S.M.; Hur, J.-S.; Kim, W. Evaluation of Antimicrobial Properties of Lichen Substances against Plant Pathogens. Plant Pathol. J. 2022, 38, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Bing, H.; Qi, C.; Gu, J.; Zhao, T.; Yu, X.; Cai, Y.; Zhang, Y.; Li, A.; Wang, X.; Zhao, J.; et al. Isolation and Identification of NEAU-CP5: A Seed-Endophytic Strain of B. velezensis That Controls Tomato Bacterial Wilt. Microb. Pathog. 2024, 192, 106707. [Google Scholar] [CrossRef]
- Kinnersley, A.M.; Scott, T.C.; Yopp, J.H.; Whitten, G.H. Promotion of Plant Growth by Polymers of Lactic Acid. Plant Growth Regul. 1990, 9, 137–146. [Google Scholar] [CrossRef]
- Semwal, P.; Misra, S.; Misra, A.; Kar, S.; Majhi, B.; Mishra, S.K.; Srivastava, S.; Chauhan, P.S. Endophytic Bacillus Strains Enhance Biomass and Bioactive Metabolites of Gloriosa superba. Ind. Crops Prod. 2023, 204, 117296. [Google Scholar] [CrossRef]
- Xie, Z.; Chu, Y.; Zhang, W.; Lang, D.; Zhang, X. Bacillus pumilus Alleviates Drought Stress and Increases Metabolite Accumulation in Glycyrrhiza uralensis Fisch. Environ. Exp. Bot. 2019, 158, 99–106. [Google Scholar] [CrossRef]
- Choi, S.-S.; Lee, S.-H.; Lee, K.-A. A Comparative Study of Hesperetin, Hesperidin and Hesperidin Glucoside: Antioxidant, Anti-Inflammatory, and Antibacterial Activities In Vitro. Antioxidants 2022, 11, 1618. [Google Scholar] [CrossRef]
- Hagel, J.M.; Chen, X.; Facchini, P.J. Production of Methylparaben in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2019, 46, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Robakowski, P.; Bielinis, E.; Stachowiak, J.; Mejza, I.; Bułaj, B. Seasonal Changes Affect Root Prunasin Concentration in Prunus serotina and Override Species Interactions between P. serotina and Quercus petraea. J. Chem. Ecol. 2016, 42, 202–214. [Google Scholar] [CrossRef]
- Jardim-Messeder, D.; Margis-Pinheiro, M.; Sachetto-Martins, G. Salicylic Acid and Adenine Nucleotides Regulate the Electron Transport System and ROS Production in Plant Mitochondria. Biochim. Et. Biophys. Acta (BBA) Bioenerg. 2022, 1863, 148559. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.V.; Pereira, N.R.; Pereira, R.C.G.; Duarte, L.P.; Takahashi, J.A.; Silva, R.R. Synthesis and Antimicrobial Activity of Ursolic Acid Ester Derivatives. Chem. Biodivers. 2022, 19, e202100566. [Google Scholar] [CrossRef]
- Guang, C.; Chen, J.; Sang, S.; Cheng, S. Biological Functionality of Soyasaponins and Soyasapogenols. J. Agric. Food Chem. 2014, 62, 8247–8255. [Google Scholar] [CrossRef]
- Zenkov, N.K.; Chechushkov, A.V.; Kozhin, P.M.; Kandalintseva, N.V.; Martinovich, G.G.; Menshchikova, E.B. Plant Phenols and Autophagy. Biochemistry 2016, 81, 297–314. [Google Scholar] [CrossRef]
- Šormová, Z.; Šebesta, K.; Bauerová, J.; Melichar, O.; Šorm, F. Stimulation of Plant Development by Some Uracil Analogues. Experientia 1960, 16, 189–190. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nunes Nesi, A.; Araújo, W.L.; Braun, H.-P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef]
- Bohner, A.; Kojima, S.; Hajirezaei, M.; Melzer, M.; von Wirén, N. Urea Retranslocation from Senescing Arabidopsis Leaves Is Promoted by DUR3-Mediated Urea Retrieval from Leaf Apoplast. Plant J. 2015, 81, 377–387. [Google Scholar] [CrossRef]
- Takahashi, T.; Kakehi, J.-I. Polyamines: Ubiquitous Polycations with Unique Roles in Growth and Stress Responses. Ann. Bot. 2010, 105, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Glutathione Synthesis. Biochim. Et. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Mielecki, J.; Gawroński, P.; Karpiński, S. Retrograde Signaling: Understanding the Communication between Organelles. Int. J. Mol. Sci. 2020, 21, 6173. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-C.; Ren, J.-J.; Zhao, H.-J.; Wang, X.-L.; Wang, T.-H.; Jin, S.-D.; Wang, Z.-H.; Li, C.; Liu, A.-R.; Lin, X.-M.; et al. Trichoderma harzianum Improves Defense against Fusarium oxysporum by Regulating ROS and RNS Metabolism, Redox Balance, and Energy Flow in Cucumber Roots. Phytopathology 2019, 109, 972–982. [Google Scholar] [CrossRef]
- Magnus, V.; Simaga, S.; Iskrić, S.; Kveder, S. Metabolism of Tryptophan, Indole-3-Acetic Acid, and Related Compounds in Parasitic Plants from the Genus Orobanche. Plant Physiol. 1982, 69, 853–858. [Google Scholar] [CrossRef]
- Sun, P.; Huang, Y.; Yang, X.; Liao, A.; Wu, J. The Role of Indole Derivative in the Growth of Plants: A Review. Front. Plant Sci. 2023, 13. [Google Scholar] [CrossRef]
- Pandey, D.; Rajendran, S.R.C.K.; Gaur, M.; Sajeesh, P.K.; Kumar, A. Plant Defense Signaling and Responses against Necrotrophic Fungal Pathogens. J. Plant Growth Regul. 2016, 35, 1159–1174. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant Flavonoids: Classification, Distribution, Biosynthesis, and Antioxidant Activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Hu, X.; Huang, Y. Guest Editorial IEEE International Conference on Bioinformatics and Biomedicine (BIBM) 2015 Special Issue. IEEE Trans. NanoBioscience 2016, 15, 306–308. [Google Scholar] [CrossRef]
- Lan, X.; Wang, W.; Li, Q.; Wang, J. The Natural Flavonoid Pinocembrin: Molecular Targets and Potential Therapeutic Applications. Mol. Neurobiol. 2016, 53, 1794–1801. [Google Scholar] [CrossRef]
- Uçar, K.; Göktaş, Z. Biological Activities of Naringenin: A Narrative Review Based on In Vitro and In Vivo Studies. Nutr. Res. 2023, 119, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ou, Y.; Hu, G.; Wen, C.; Yue, S.; Chen, C.; Xu, L.; Xie, J.; Dai, H.; Xiao, H.; et al. Naringenin Attenuates Non-Alcoholic Fatty Liver Disease by down-Regulating the NLRP3/NF-κB Pathway in Mice. Br. J. Pharmacol. 2020, 177, 1806–1821. [Google Scholar] [CrossRef]
- Guo, M.; Li, C.; Huang, R.; Qu, L.; Liu, J.; Zhang, C.; Ge, Y. Ferulic Acid Enhanced Resistance against Blue Mold of Malus domestica by Regulating Reactive Oxygen Species and Phenylpropanoid Metabolism. Postharvest Biol. Technol. 2023, 202, 112378. [Google Scholar] [CrossRef]
- Tenenboim, H.; Burgos, A.; Willmitzer, L.; Brotman, Y. Using Lipidomics for Expanding the Knowledge on Lipid Metabolism in Plants. Biochimie 2016, 130, 91–96. [Google Scholar] [CrossRef]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of Arachidonic Acid Metabolism: A Review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Goossens, A.; Lacchini, E. Jasmonate: A Hormone of Primary Importance for Plant Metabolism. Curr. Opin. Plant Biol. 2022, 67, 102197. [Google Scholar] [CrossRef]
- Guerreiro, A.; Figureueiredo, J.; Sousa Silva, M.; Figureueiredo, A. Linking Jasmonic Acid to Grapevine Resistance against the Biotrophic Oomycete Plasmopara viticola. Front. Plant Sci. 2016, 7, 565. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Tian, L.; Zhu, Z.; Sang, Z.; Ma, L.; Jia, Z. Growth and Physiological Responses of Magnoliaceae to NaCl Stress. Plants 2024, 13, 170. [Google Scholar] [CrossRef]
- Zhang, C.; Ou, X.; Wang, J.; Wang, Z.; Du, W.; Zhao, J.; Han, Y. Antifungal Peptide P852 Controls Fusarium Wilt in Faba Bean (Viciafaba L.) by Promoting Antioxidant Defense and Isoquinoline Alkaloid, Betaine, and Arginine Biosyntheses. Antioxidants 2022, 11, 1767. [Google Scholar] [CrossRef]
| Groups | Plant Height (cm) | Number of Roots (Pieces) | Root Length (cm) | Root Volume (mL) | Leaf Length (cm) |
|---|---|---|---|---|---|
| CK | 49.50 ± 2.179 a | 6.667 ± 1.528 a | 7.333 ± 1.528 a | 0.04333 ± 0.005774 a | 34.33 ± 3.215 a |
| ZB | 34.33 ± 9.018 b | 5.000 ± 1.000 a | 6.067 ± 1.290 a | 0.01667 ± 0.005774 a | 19.00 ± 5.568 b |
| M | 51.83 ± 4.537 a | 5.333 ± 1.155 a | 7.067 ± 1.888 a | 0.06000 ± 0.01732 a | 37.00 ± 2.646 a |
| X7 | 49.50 ± 2.179 a | 7.667 ± 1.155 a | 8.333 ± 2.255 a | 0.05667 ± 0.02887 a | 36.83 ± 3.547 a |
| HX | 50.00 ± 2.179 a | 6.000 ± 1.732 a | 13.93 ± 6.361 a | 0.06000 ± 0.02646 a | 35.07 ± 3.614 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Li, J.; Wang, H.; Liu, X.; Gao, Z.; Chen, J.; Han, Y. Metabolic and Antioxidant Responses of Different Control Methods to the Interaction of Sorghum sudangrass hybrids-Colletotrichum boninense. Int. J. Mol. Sci. 2024, 25, 9505. https://doi.org/10.3390/ijms25179505
Xu J, Li J, Wang H, Liu X, Gao Z, Chen J, Han Y. Metabolic and Antioxidant Responses of Different Control Methods to the Interaction of Sorghum sudangrass hybrids-Colletotrichum boninense. International Journal of Molecular Sciences. 2024; 25(17):9505. https://doi.org/10.3390/ijms25179505
Chicago/Turabian StyleXu, Jingxuan, Junying Li, Hongji Wang, Xinhao Liu, Zhen Gao, Jie Chen, and Yuzhu Han. 2024. "Metabolic and Antioxidant Responses of Different Control Methods to the Interaction of Sorghum sudangrass hybrids-Colletotrichum boninense" International Journal of Molecular Sciences 25, no. 17: 9505. https://doi.org/10.3390/ijms25179505