
Study of Pentacyclic Triterpenes from Lyophilised Aguaje: Anti-Inflammatory and Antioxidant Properties

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Isolation and Characterisation of Compounds of M. flexuosa
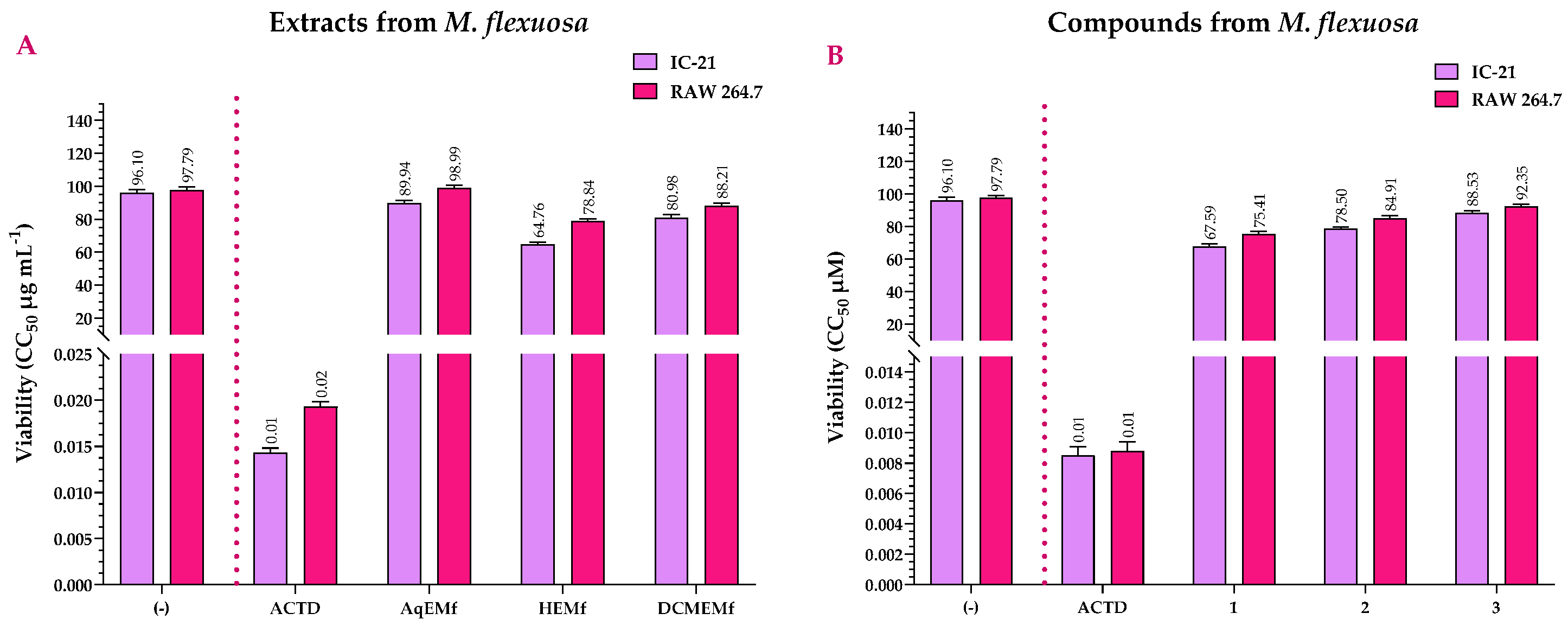
2.2. Viability Assay of the Extracts and Compounds of M. flexuosa
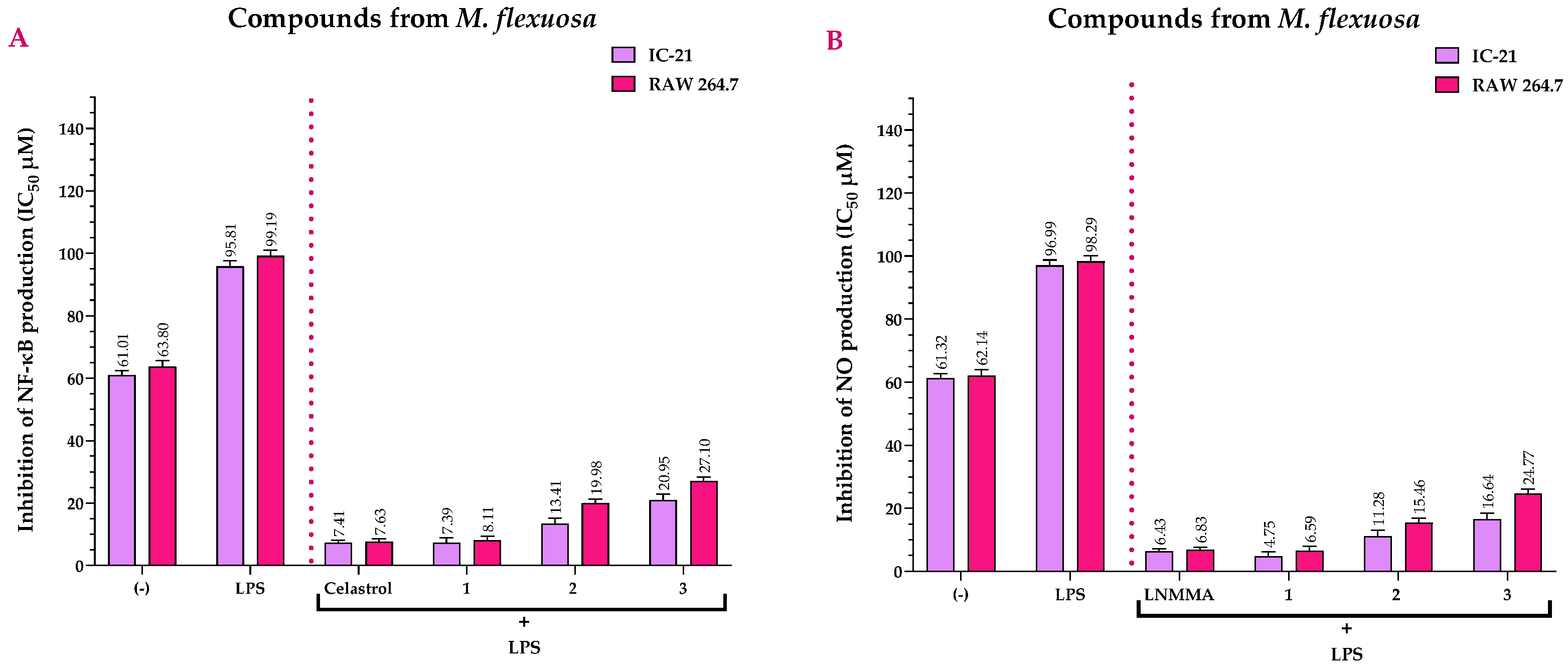
2.3. Anti-Inflammatory Activity of the Extracts and Compounds of M. flexuosa
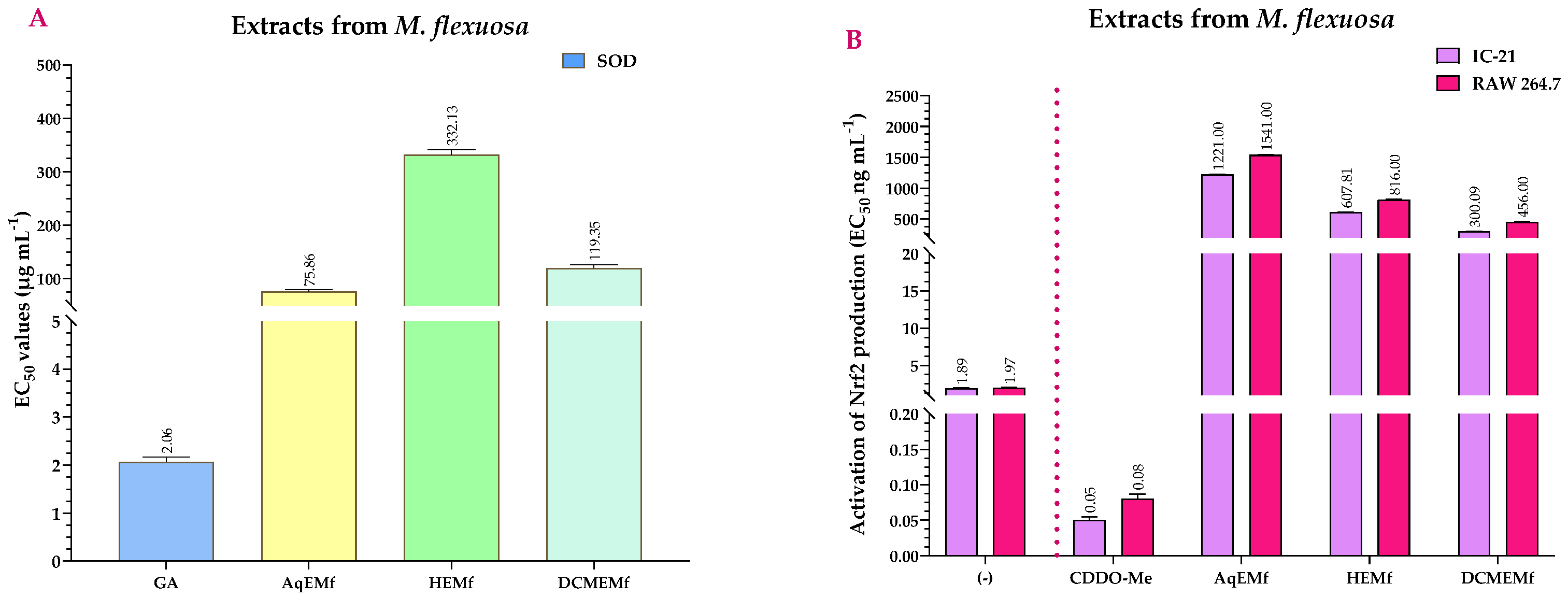
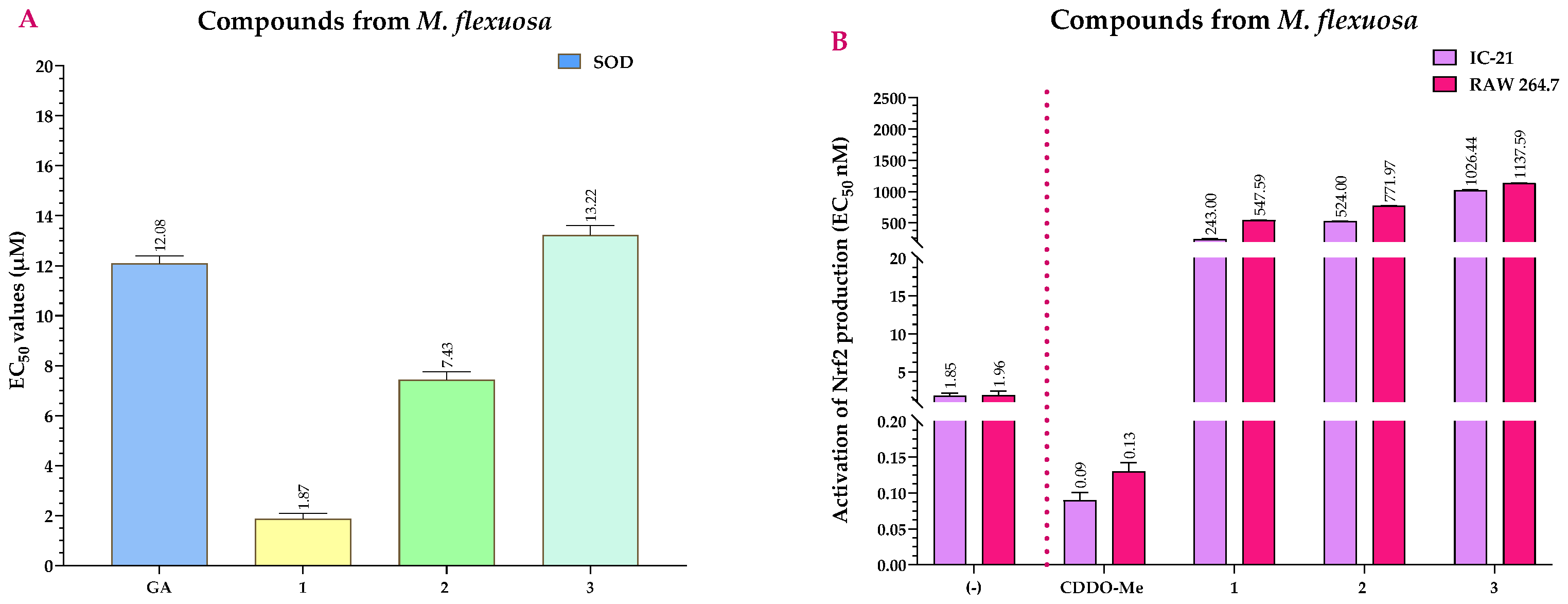
2.4. Antioxidant Activity of the Extracts and Compounds of M. flexuosa
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Extraction and Isolation
4.3. Spectroscopic Data
4.3.1. 3,11-Dioxours-12-en-28-oic Acid (1)
4.3.2. (3β)-3-Acetyloxy-11-oxours-12-en-28-oic Acid (2)
4.3.3. (3β)-3-Hydroxy-11-oxours-12-en-28-oic Acid (3)
4.4. Cell Culture
4.5. Statistical Analysis
4.6. In Vitro Viability Assay
4.7. Anti-Inflammatory Assays
4.7.1. NF-κB Inhibition Assay
4.7.2. NO Inhibition Assay
4.8. Antioxidant Assays
4.8.1. SOD Assay
4.8.2. Nrf2 Activity Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roe, K. An inflammation classification system using cytokine parameters. Scand. J. Immunol. 2021, 93, e12970. [Google Scholar] [CrossRef] [PubMed]
- Barnabei, L.; Laplantine, E.; Mbongo, W.; Rieux-Laucat, F.; Weil, R. NF-κB: At the Borders of Autoimmunity and Inflammation. Front. Immunol. 2021, 12, 716469. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 86. [Google Scholar] [CrossRef]
- Cinelli, M.A.; Do, H.T.; Miley, G.P.; Silverman, R.B. Inducible nitric oxide synthase: Regulation, structure, and inhibition. Med. Res. Rev. 2020, 40, 158–189. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Tran, G.B.; Nguyen, C.T. Anti-oxidative effects of superoxide dismutase 3 on inflammatory diseases. J. Mol. Med. 2020, 98, 59–69. [Google Scholar] [CrossRef]
- Wang, X.; Gong, Y.; Zhou, B.; Yang, J.; Cheng, Y.; Zhao, J.; Qi, M. Ursolic acid ameliorates oxidative stress, inflammation, and fibrosis in diabetic cardiomyopathy rats. Biomed. Pharmacother. 2018, 97, 1461–1467. [Google Scholar] [CrossRef]
- Minhas, R.; Bansal, Y.; Bansal, G. Inducible nitric oxide synthase inhibitors: A comprehensive update. Med. Res. Rev. 2020, 40, 823–855. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Yamawaki, K. Bardoxolone methyl: Drug development for diabetic kidney disease. Clin. Exp. Nephrol. 2020, 24, 857–864. [Google Scholar] [CrossRef]
- Robledinos-Antón, N.; Fernández-Ginés, R.; Manda, G.; Cuadrado, A. Activators and Inhibitors of NRF2: A Review of Their Potential for Clinical Development. Oxid. Med. Cell. Longev. 2019, 2019, 9372182. [Google Scholar] [CrossRef]
- Borgstahl, G.E.O.; Oberley-Deegan, R.E. Superoxide Dismutases (SODs) and SOD Mimetics. Antioxidants 2018, 7, 156. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Panahi, Y.; Rahimnia, A.R.; Sharafi, M.; Alishiri, G.; Saburi, A.; Sahebkar, A. Curcuminoid treatment for knee osteoarthritis: A randomized double-blind placebo-controlled trial. Phytother. Res. 2014, 28, 1625–1631. [Google Scholar] [CrossRef]
- Comalada, M.; Camuesco, D.; Sierra, S.; Ballester, I.; Xaus, J.; Gálvez, J.; Zarzuelo, A. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-kappaB pathway. Eur. J. Immunol. 2005, 35, 584–592. [Google Scholar] [CrossRef]
- Shao, Z.H.; Becker, L.B.; Vanden Hoek, T.L.; Schumacker, P.T.; Li, C.Q.; Zhao, D.; Wojcik, K.; Anderson, T.; Qin, Y.; Dey, L.; et al. Grape seed proanthocyanidin extract attenuates oxidant injury in cardiomyocytes. Pharmacol. Res. 2003, 47, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Yokozawa, T. Direct scavenging of nitric oxide and superoxide by green tea. Food Chem. Toxicol. 2002, 40, 1745–1750. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane and Other Nutrigenomic Nrf2 Activators: Can the Clinician’s Expectation Be Matched by the Reality? Oxid. Med. Cell. Longev. 2016, 2016, 7857186. [Google Scholar] [CrossRef]
- Benitez Mora, M.P.; Kosior, M.A.; Damiano, S.; Longobardi, V.; Presicce, G.A.; Di Vuolo, G.; Pacelli, G.; Campanile, G.; Gasparrini, B. Dietary supplementation with green tea extract improves the antioxidant status and oocyte developmental competence in Italian Mediterranean buffaloes. Theriogenology 2024, 215, 50–57. [Google Scholar] [CrossRef]
- Fuentes, E.; Rodríguez-Pérez, W.; Guzmán, L.; Alarcón, M.; Navarrete, S.; Forero-Doria, O.; Palomo, I. Mauritia flexuosa Presents In Vitro and In Vivo Antiplatelet and Antithrombotic Activities. Evid. Based Complement. Alternat. Med. 2013, 2013, 2013653257. [Google Scholar] [CrossRef] [PubMed]
- Sotero, V.; Luján, M.; Freitas, L.; Merino, C.; Dávila, É. Stability and antioxidant activity of freeze-dried flour of three morphotypes of aguaje (Mauritia flexuosa L. f.). Rev. Soc. Quím. Perú. 2013, 79, 136–143. [Google Scholar]
- Garzón Gómez, C.; Macuritofe Ramírez, V. La Noche, las Plantas y sus Sueños: Aproximación al Conocimiento Botánico en una Cultura Amazónica, 1st ed.; Corporación Colombiana para la Amazonia: Bogotá, Colombia, 1990. [Google Scholar]
- Bensaada, H.; Soto-Garcia, M.F.; Carmona-Hernandez, J.C. Antioxidant Activity of Polyphenols, from Mauritia flexuosa (Aguaje), Based on Controlled Dehydration. Molecules 2022, 27, 3065. [Google Scholar] [CrossRef] [PubMed]
- Koolen, H.H.F.; da Silva, F.M.A.; da Silva, V.S.V.; Paz, W.H.P.; Bataglion, G.A. Buriti fruit-Mauritia flexuosa. In Exotic Fruits; Rodrigues, S., de Oliveira Silva, E., Sousa de Brito, E., Eds.; Academic Express: London, UK, 2018; pp. 61–66. [Google Scholar] [CrossRef]
- Santos, M.F.; Marmesat, S.; Brito, E.S.; Alves, R.E.; Dobarganes, M.C. Major components in oils obtained from Amazonian palm fruits. Grasas Aceites 2013, 64, 328–334. [Google Scholar] [CrossRef]
- do Nascimento Silva, N.R.R.; Cavalcante, R.B.M.; da Silva, F.A. Nutritional properties of Buriti (Mauritia flexuosa) and health benefits. J. Food Compos. Anal. 2023, 117, 105092. [Google Scholar] [CrossRef]
- Bataglion, G.A.; da Silva, F.M.A.; Santos, J.M.; Barcia, M.T.; Godoy, H.T.; Eberlin, M.N.; Koolen, H.H.F. Integrative Approach Using GC-MS and Easy Ambient Sonic-Spray Ionization Mass Spectrometry (EASI-MS) for Comprehensive Lipid Characterization of Buriti (Mauritia flexuosa) Oil. J. Braz. Chem. Soc. 2015, 26, 171–177. [Google Scholar] [CrossRef]
- Yang, B.; Zhu, J.P.; Rong, L.; Jin, J.; Cao, D.; Li, H.; Zhou, X.H.; Zhao, Z.X. Triterpenoids with antiplatelet aggregation activity from Ilex rotunda. Phytochemistry. 2018, 145, 179–186. [Google Scholar] [CrossRef]
- Sutthivaiyakit, S.; Seeka, C.; Kritwinyu, T.; Pisutchareonpong, S.; Chimnoi, N. 19-nor-11-Oxooleanan-12-ene and 18,19-seco-19-oxours-11,13(18)-diene triterpenes from Diospyros decandra bark. Tetrahedron Lett. 2012, 53, 1713–1716. [Google Scholar] [CrossRef]
- Lin, Z.; Tong, Y.; Li, N.; Zhu, Z.; Li, J. Network pharmacology-based study of the mechanisms of action of anti-diabetic triterpenoids from Cyclocarya paliurus. RSC Adv. 2020, 10, 37168–37181. [Google Scholar] [CrossRef] [PubMed]
- Nistor, M.; Rugina, D.; Diaconeasa, Z.; Socaciu, C.; Socaciu, M.A. Pentacyclic Triterpenoid Phytochemicals with Anticancer Activity: Updated Studies on Mechanisms and Targeted Delivery. Int. J. Mol. Sci. 2023, 24, 12923. [Google Scholar] [CrossRef]
- Holden, L.; Burke, C.S.; Cullinane, D.; Keyes, E.T. Strategies to promote permeation and vectorization, and reduce cytotoxicity of metal complex luminophores for bioimaging and intracellular sensing. RSC Chem. Biol. 2021, 2, 1021–1049. [Google Scholar] [CrossRef]
- ADMAlab 2.0. Available online: https://admetmesh.scbdd.com (accessed on 2 August 2024).
- Komleva, N.V.; Lapshina, M.A.; Kostyuk, G.V.; Ivanov, A.V.; Parkhomenko, I.I.; Papina, R.I.; Sen, V.D.; Terentiev, A.A. Comparative analysis of cytotoxic effects and intracellular accumulation of platinum(IV) nitroxyl complexes. Russ. Chem. Bull. 2015, 64, 1178–1182. [Google Scholar] [CrossRef]
- John, G.W.; Shrivastava, R.; Chevalier, A.; Pognat, J.F.; Massingham, R. An in vitro investigation of the relationships between potency, lipophilicity, cytotoxicity and chemical class of representative calcium antagonist drugs. Pharmacol. Res. 1993, 27, 253–262. [Google Scholar] [CrossRef]
- Rodrigues, V.; Do Nascimento, D.; Do Nascimento, J.; Silva, C.; Nunes, L.; Cardoso, A.; Ribeiro, F.; Castro, F.; Santos, M.; Alves, J.; et al. Anti-inflammatory mechanisms of fruits and by-products from Mauritia flexuosa, an exotic plant with functional benefits. J. Environ. Toxicol. 2021, 84, 441–457. [Google Scholar] [CrossRef]
- Chun, J.; Lee, C.; Hwang, S.W.; Im, J.P.; Kim, J.S. Ursolic acid inhibits nuclear factor-κB signaling in intestinal epithelial cells and macrophages, and attenuates experimental colitis in mice. Life Sci. 2014, 110, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Li, Z.; Hua, Q.; Song, P.; Gao, L.; Zhou, L.; Cai, Q. Ursolic Acid Alleviates Neuroinflammation after Intracerebral Hemorrhage by Mediating Microglial Pyroptosis via the NF-κB/NLRP3/GSDMD Pathway. Int. J. Mol. Sci. 2023, 24, 14771. [Google Scholar] [CrossRef]
- Li, J.; Li, N.; Yan, S.; Liu, M.; Sun, B.; Lu, Y.; Shao, Y. Ursolic acid alleviates inflammation and against diabetes-induced nephropathy through TLR4-mediated inflammatory pathway. Mol. Med. Rep. 2018, 18, 4675–4681. [Google Scholar] [CrossRef]
- Wang, B.; Parobchak, N.; Martin, A.; Rosen, M.; Yu, L.J.; Nguyen, M.; Gololobova, K.; Rosen, T. Screening a small molecule library to identify inhibitors of NF-κB inducing kinase and pro-labor genes in human placenta. Sci. Rep. 2018, 8, 1657. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rodríguez, A.; González-Ortiz, M.; Martínez-Abundis, E.; Acuña, N. Effect of Ursolic Acid on Metabolic Syndrome, Insulin Sensitivity, and Inflammation. J. Med. Food. 2017, 20, 882–886. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed]
- Roberti, A.; Chaffey, L.E.; Greaves, D.R. NF-κB Signaling and Inflammation-Drug Repurposing to Treat Inflammatory Disorders? Biology 2022, 11, 372. [Google Scholar] [CrossRef]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef]
- Chen, H.; Gao, Y.; Wang, A.; Zhou, X.; Zheng, Y.; Zhou, J. Evolution in medicinal chemistry of ursolic acid derivatives as anticancer agents. Eur. J. Med. Chem. 2015, 92, 648–655. [Google Scholar] [CrossRef]
- Patil, K.R.; Mohapatra, P.; Patel, H.M.; Goyal, S.N.; Ojha, S.; Kundu, C.N.; Patil, C.R. Pentacyclic Triterpenoids Inhibit IKKβ Mediated Activation of NF-κB Pathway: In Silico and In Vitro Evidences. PLoS ONE 2015, 10, e0125709. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.R.; Prasad, S.; Sung, B.; Kannappan, R.; Aggarwal, B.B. Targeting inflammatory pathways by triterpenoids for prevention and treatment of cancer. Toxins 2010, 2, 2428–2466. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.Y.; Dai, S.J.; Zhao, F.; Liu, J.F.; Fang, W.S.; Liu, K. New ursane-type triterpene with NO production suppressing activity from Nauclea officinalis. J. Asian Nat. Prod. Res. 2012, 14, 97–104. [Google Scholar] [CrossRef]
- Gatbonton-Schwager, T.N.; Letterio, J.J.; Tochtrop, G.P. Bryonolic acid transcriptional control of anti-inflammatory and antioxidant genes in macrophages in vitro and in vivo. J. Nat. Prod. 2012, 75, 591–598. [Google Scholar] [CrossRef]
- Yang, Z.G.; Li, H.R.; Wang, L.Y.; Li, Y.H.; Lu, S.G.; Wen, X.F.; Wang, J.; Daikonya, A.; Kitanaka, S. Triterpenoids from Hippophae rhamnoides L. and their nitric oxide production-inhibitory and DPPH radical-scavenging activities. Chem. Pharm. Bull. 2007, 55, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Suh, N.; Honda, T.; Finlay, H.J.; Barchowsky, A.; Williams, C.; Benoit, N.E.; Xie, Q.W.; Nathan, C.; Gribble, G.W.; Sporn, M.B. Novel triterpenoids suppress inducible nitric oxide synthase (iNOS) and inducible cyclooxygenase (COX-2) in mouse macrophages. Cancer Res. 1998, 58, 717–723. [Google Scholar] [PubMed]
- Özenver, N.; Efferth, T. Small molecule inhibitors and stimulators of inducible nitric oxide synthase in cancer cells from natural origin (phytochemicals, marine compounds, antibiotics). Biochem. Pharmacol. 2020, 176, 113792. [Google Scholar] [CrossRef]
- Katsuyama, K.; Shichiri, M.; Marumo, F.; Hirata, Y. NO inhibits cytokine-induced iNOS expression and NF-kappaB activation by interfering with phosphorylation and degradation of IkappaB-alpha. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1796–1802. [Google Scholar] [CrossRef]
- Jia, J.; Liu, Y.; Zhang, X.; Liu, X.; Qi, J. Regulation of iNOS expression by NF-κB in human lens epithelial cells treated with high levels of glucose. Invest. Ophthalmol. Vis. Sci. 2013, 54, 5070–5077. [Google Scholar] [CrossRef]
- Rivero-Chan, B.E.; Marrero, J.G.; Hernández-Ortega, S.; Mena-Rejón, G.J.; Miranda, L.D. Rapid access to ketones related to oleanolic and ursolic acids. Nat. Prod. Res. 2012, 26, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Harun, N.H.; Septama, A.W.; Ahmad, W.A.N.W.; Suppian, R. Immunomodulatory effects and structure-activity relationship of botanical pentacyclic triterpenes: A review. Chin. Herb. Med. 2020, 12, 118–124. [Google Scholar] [CrossRef]
- Veerappan, K.; Natarajan, S.; Ethiraj, P.; Vetrivel, U.; Samuel, S. Inhibition of IKKβ by Celastrol and its analogues -an in silico and in vitro approach. Pharm. Biol. 2017, 55, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Lauria, A.; Ippolito, M.; Fazzari, M.; Tutone, M.; Di Blasi, F.; Mingoia, F.; Almerico, A.M. IKK- inhibitors: An analysis of drug–receptor interaction by using Molecular Docking and Pharmacophore 3D-QSAR approaches. J. Mol. Graph. Modell. 2010, 29, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Ngo, V.; Duennwald, M.L. Nrf2 and Oxidative Stress: A General Overview of Mechanisms and Implications in Human Disease. Antioxidants 2022, 11, 2345. [Google Scholar] [CrossRef]
- Barboza, N.L.; Cruz, J.M.D.A.; Corrêa, R.F.; Lamarão, C.V.; Lima, A.R.; Inada, N.M.; Sanches, E.A.; Bezerra, J.A.; Campelo, P.H. Buriti (Mauritia flexuosa L. f.): An Amazonian fruit with potential health benefits. Food Res. Int. 2022, 159, 111654. [Google Scholar] [CrossRef]
- Nobre, C.B.; Sousa, E.O.; Camilo, C.J.; Machado, J.F.; Silva, J.M.F.L.; Filho, J.R.; Coutinho, H.D.M.; Costa, J.G.M. Antioxidative effect and phytochemical profile of natural products from the fruits of “babaçu” (Orbignia speciose) and “buriti” (Mauritia flexuosa). Food Chem. Toxicol. 2018, 121, 423–429. [Google Scholar] [CrossRef]
- Wang, Z.; Song, S.; Lu, H.; Chen, G.; Xu, S.; Sagara, Y.; Kitaoka, N.; Manabe, M.; Kodama, H. Effect of three triterpenoid compounds isolated from root bark of Aralia elata on stimulus-induced superoxide generation and tyrosyl phosphorylation and translocation of p47(phox) and p67(phox) to cell membrane in human neutrophil. Clin. Chim. Acta 2003, 336, 65–72. [Google Scholar] [CrossRef]
- Ling, T.; Boyd, L.; Rivas, F. Triterpenoids as Reactive Oxygen Species Modulators of Cell Fate. Chem. Res. Toxicol. 2022, 35, 569–584. [Google Scholar] [CrossRef]
- Staurengo-Ferrari, L.; Badaro-Garcia, S.; Hohmann, M.S.N.; Manchope, M.F.; Zaninelli, T.H.; Casagrande, R.; Verri, W.A., Jr. Contribution of Nrf2 Modulation to the Mechanism of Action of Analgesic and Anti-inflammatory Drugs in Pre-clinical and Clinical Stages. Front. Pharmacol. 2019, 9, 1536. [Google Scholar] [CrossRef]
- Osburn, W.O.; Kensler, T.W. Nrf2 signaling. An adaptive response pathway for protection against environmental toxic insults. Mutat. Res. 2008, 659, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Kamble, S.M.; Patel, H.M.; Goyal, S.N.; Noolvi, M.N.; Mahajan, U.B.; Ojha, S.; Patil, C.R. In silico Evidence for Binding of Pentacyclic Triterpenoids to Keap1-Nrf2 Protein-Protein Binding Site. Comb. Chem. High Throughput Screen. 2017, 20, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Baer-Dubowska, W.; Narożna, M.; Krajka-Kuźniak, V. Anti-Cancer Potential of Synthetic Oleanolic Acid Derivatives and Their Conjugates with NSAIDs. Molecules 2021, 26, 4957. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D.; Lo, S.C.; Sun, Z.; Habib, G.M.; Lieberman, M.W.; Hannink, M. Ubiquitination of Keap1, a BTB-Kelch Substrate Adaptor Protein for Cul3, Targets Keap1 for Degradation by a Proteasome-independent Pathway. J. Biol. Chem. 2005, 280, 30091–30099. [Google Scholar] [CrossRef] [PubMed]
- Egbujor, M.C.; Tucci, P.; Onyeije, U.C.; Emeruwa, C.N.; Saso, L. NRF2 Activation by Nitrogen Heterocycles: A Review. Molecules 2023, 28, 2751. [Google Scholar] [CrossRef]
- El Kebbaj, R.; Bouchab, H.; Tahri-Joutey, M.; Rabbaa, S.; Limami, Y.; Nasser, B.; Egbujor, M.C.; Tucci, P.; Andreoletti, P.; Saso, L.; et al. The Potential Role of Major Argan Oil Compounds as Nrf2 Regulators and Their Antioxidant Effects. Antioxidants 2024, 13, 344. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Goyal, S.N.; Suchal, K.; Sharma, C.; Patil, C.R.; Ojha, S.K. Pharmacological Properties, Molecular Mechanisms, and Pharmaceutical Development of Asiatic Acid: A Pentacyclic Triterpenoid of Therapeutic Promise. Front. Pharmacol. 2018, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- Na, H.K.; Surh, Y.J. Modulation of Nrf2-mediated antioxidant and detoxifying enzyme induction by the green tea polyphenol EGCG. Food Chem. Toxicol. 2008, 46, 1271–1278. [Google Scholar] [CrossRef]
- Apaza Ticona, L.; Rumbero Sánchez, Á.; Sánchez Sánchez-Corral, J.; Iglesias Moreno, P.; Ortega Domenech, M. Anti-inflammatory, pro-proliferative and antimicrobial potential of the compounds isolated from Daemonorops draco (Willd.) Blume. J. Ethnopharmacol. 2021, 268, 113668. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Chang, J.Y.; Kuo, C.C.; Chang, C.Y.; Kuo, Y.H. Cytotoxic triterpenes from the aerial roots of Ficus microcarpa. Phytochemistry 2005, 66, 495–501. [Google Scholar] [CrossRef]
- Leal, A.S.; Wang, R.; Salvador, J.A.; Jing, Y. Synthesis of novel ursolic acid heterocyclic derivatives with improved abilities of antiproliferation and induction of p53, p21waf1 and NOXA in pancreatic cancer cells. Bioorg. Med. Chem. 2012, 20, 5774–5786. [Google Scholar] [CrossRef]
- Seebacher, W.; Simic, N.; Weis, R.; Saf, R.; Kunert, O. Complete assignments of 1H and 13C NMR resonances of oleanolic acid, 18α-oleanolic acid, ursolic acid and their 11-oxo derivatives. Magn. Reson. Chem. 2003, 41, 636–638. [Google Scholar] [CrossRef]
- Riastri, A.; Putri, D.D.; Sa’adah, M.; Gani, A.P.; Murwanti, R. RAW 264.7 Macrophage Cell Line: In Vitro Model for the Evaluation of the Immunomodulatory Activity of Zingiberaceae. Trop. J. Nat. Prod. Res. TJNPR 2023, 7, 2316–2324. [Google Scholar]
- White, D.D.; Fox, P.K.; Livingston, P. An in vitro mouse macrophage model for use in assessing the effects of anti-inflammatory drugs. In Inflammation: Mechanisms and Treatment, 1st ed.; Willoughby, D.A., Giroud, J.P., Eds.; Wiley-Blackwell: London, UK, 2006; pp. 355–364. [Google Scholar]
- Gomez Perez, M.; Fourcade, L.; Mateescu, M.A.; Paquin, J. Neutral Red versus MTT assay of cell viability in the presence of copper compounds. Anal. Biochem. 2017, 535, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Apaza Ticona, L.; Sánchez Sánchez-Corral, J.; Díaz-Guerra Martín, C.; Calderón Jiménez, S.; López González, A.; Thiebaut Estrada, C. Rubus urticifolius Compounds with Antioxidant Activity, and Inhibition Potential against Tyrosinase, Melanin, Hyaluronidase, Elastase, and Collagenase. Pharmaceuticals 2024, 17, 937. [Google Scholar] [CrossRef]
- Guo, B.; Zhao, C.; Zhang, C.; Xiao, Y.; Yan, G.; Liu, L.; Pan, H. Elucidation of the anti-inflammatory mechanism of Er Miao San by integrative approach of network pharmacology and experimental verification. Pharmacol. Res. 2022, 175, 106000. [Google Scholar] [CrossRef]
- Vincent, A.; Thauvin, M.; Quévrain, E.; Mathieu, E.; Layani, S.; Seksik, P.; Batinic-Haberle, I.; Vriz, S.; Policar, C.; Delsuc, N. Evaluation of the compounds commonly known as superoxide dismutase and catalase mimics in cellular models. J. Inorg. Biochem. 2021, 219, 111431. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apaza Ticona, L.; Sánchez Sánchez-Corral, J.; Montoto Lozano, N.; Prieto Ramos, P.; Sánchez, Á.R. Study of Pentacyclic Triterpenes from Lyophilised Aguaje: Anti-Inflammatory and Antioxidant Properties. Int. J. Mol. Sci. 2024, 25, 9615. https://doi.org/10.3390/ijms25179615
Apaza Ticona L, Sánchez Sánchez-Corral J, Montoto Lozano N, Prieto Ramos P, Sánchez ÁR. Study of Pentacyclic Triterpenes from Lyophilised Aguaje: Anti-Inflammatory and Antioxidant Properties. International Journal of Molecular Sciences. 2024; 25(17):9615. https://doi.org/10.3390/ijms25179615
Chicago/Turabian StyleApaza Ticona, Luis, Javier Sánchez Sánchez-Corral, Natalia Montoto Lozano, Pablo Prieto Ramos, and Ángel Rumbero Sánchez. 2024. "Study of Pentacyclic Triterpenes from Lyophilised Aguaje: Anti-Inflammatory and Antioxidant Properties" International Journal of Molecular Sciences 25, no. 17: 9615. https://doi.org/10.3390/ijms25179615