Long Dynamic β1–β2 Loops in M. tb MazF Toxins Affect the Interaction Modes and Strengths of the Toxin–Antitoxin Pairs


Abstract
1. Introduction
2. Results
2.1. Structures of Apo-MazF-mt3
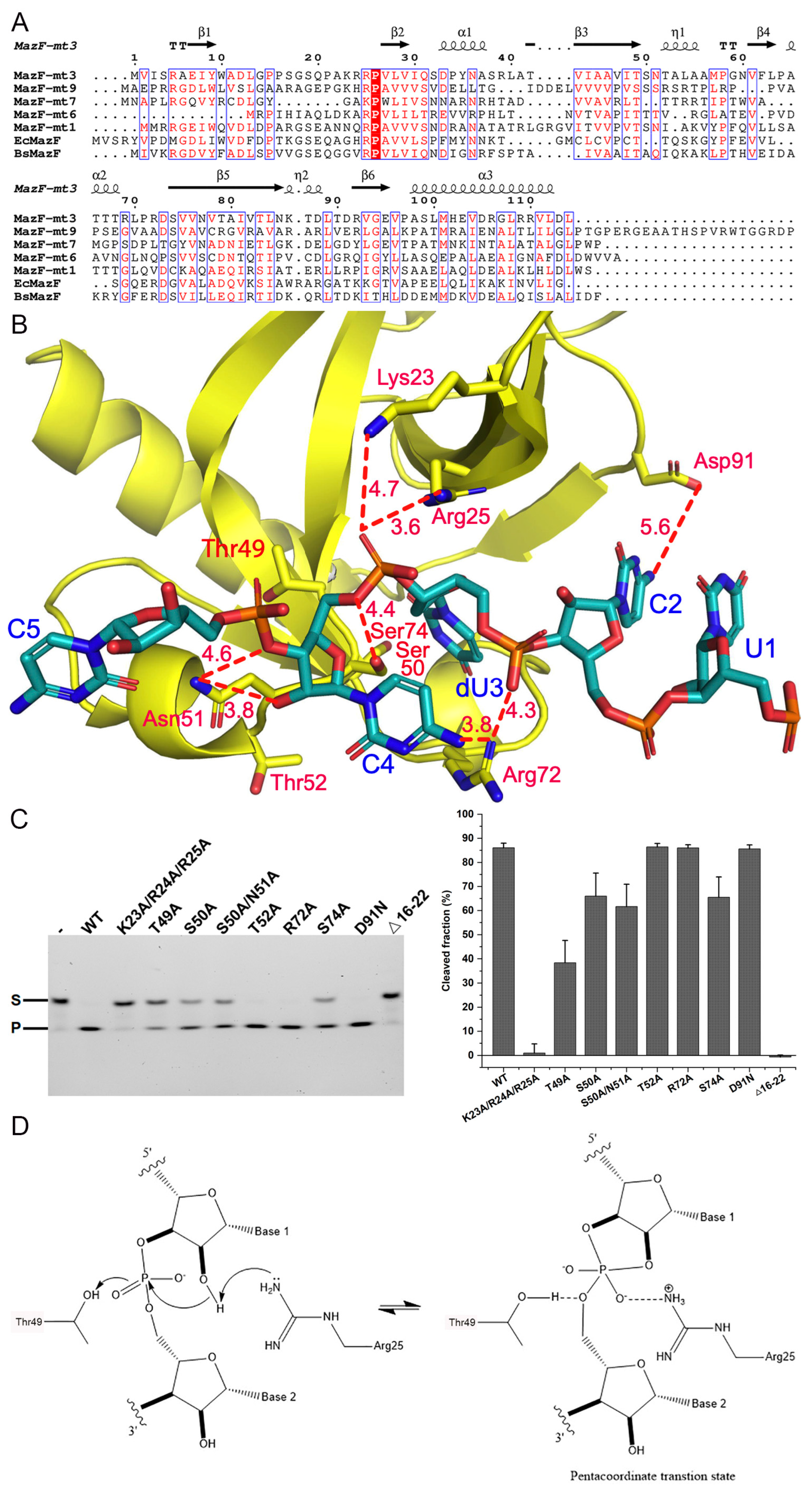
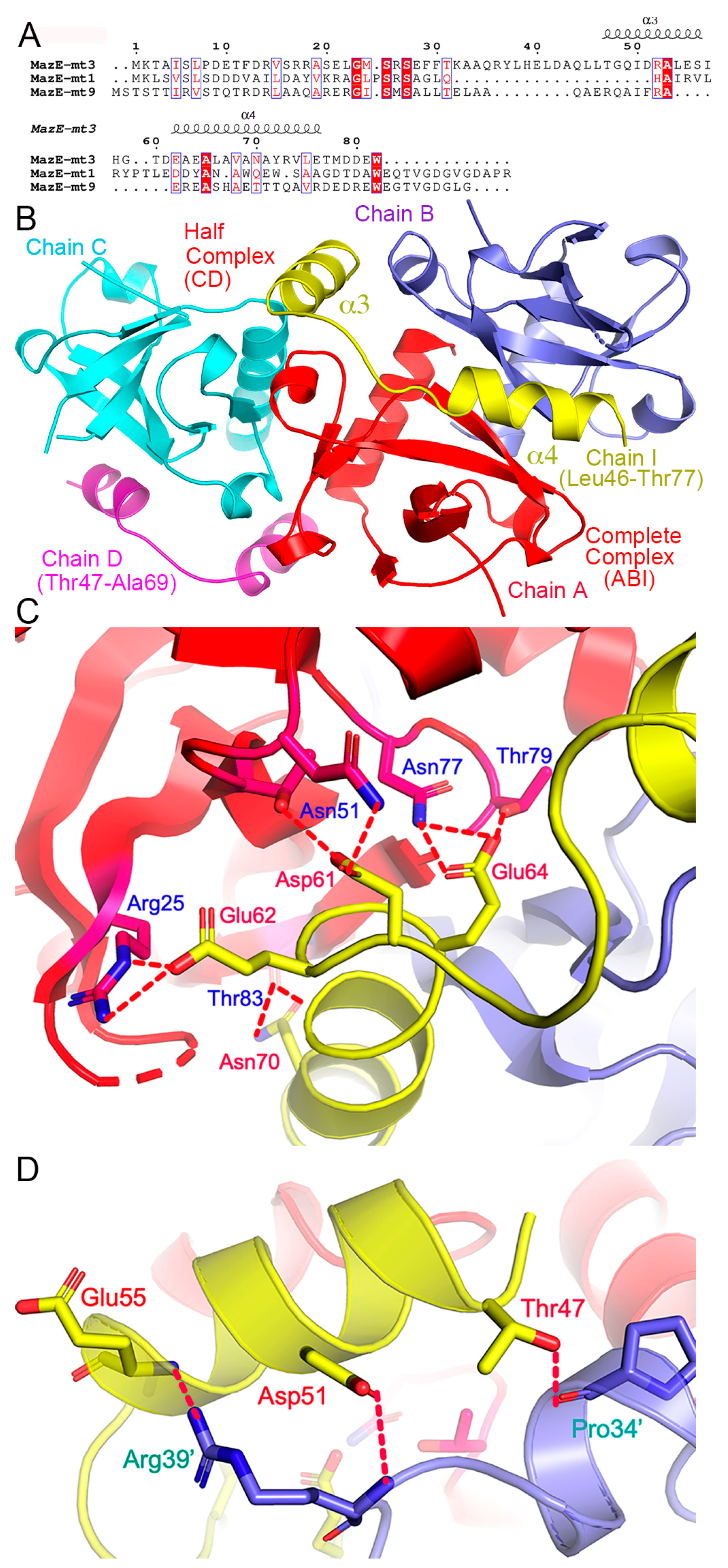
2.2. Sequence and Structure Comparison with of Other Members of the MazEF Family
2.3. RNA Cleavage Activity and Key Residues Involved
2.4. Structures of the MazF-mt3 in Complex of Its Antitoxin Fragment
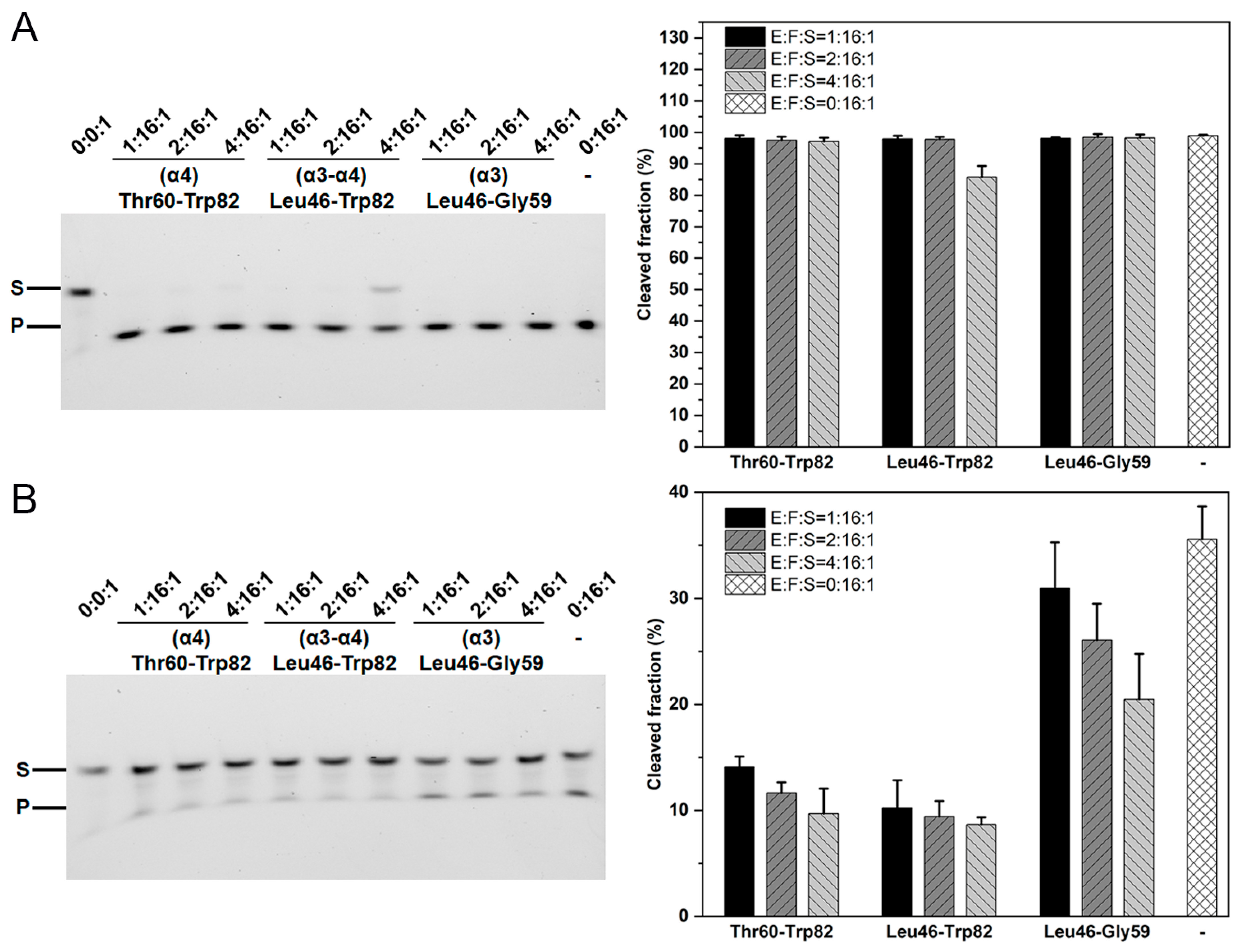
2.5. The Role of the β1–β2 Loop in the Toxin–Antitoxin Interactions
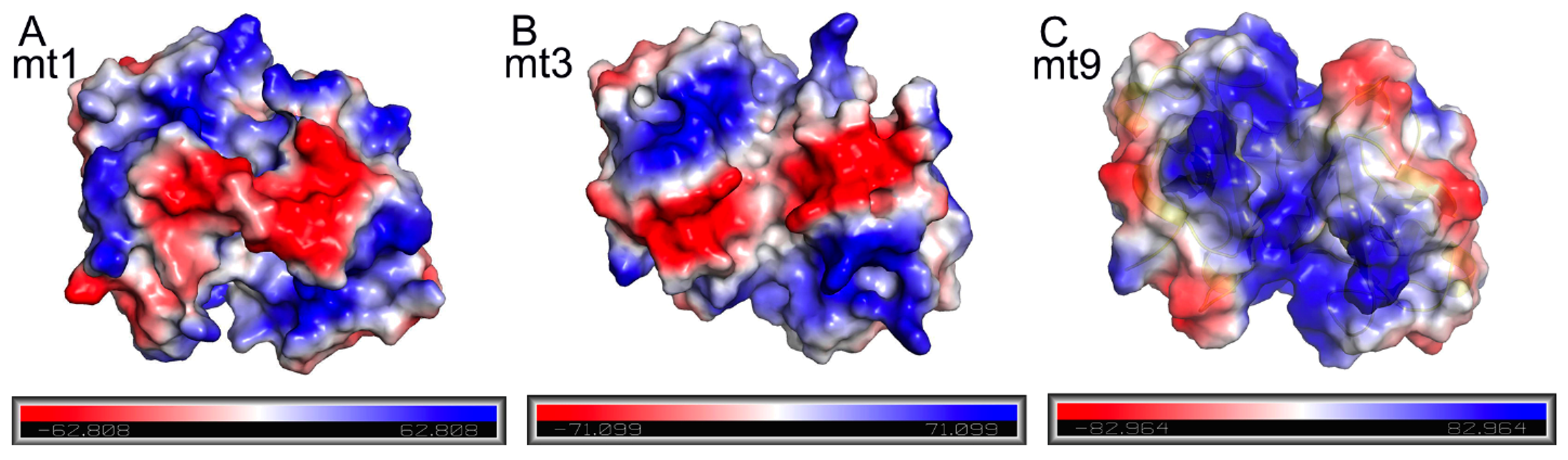
2.6. The Effects of TA Charge Complementation on Binding
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Cloning, Expression, and Purification of Proteins
4.2.2. Crystallization
4.2.3. Data Collection and Structure Determination
4.2.4. In Vitro Cleavage Assays
4.2.5. Thermal Shift Assay (TSA)
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2023; World Health Organization: Geneva, Switzerland, 2023. [Google Scholar]
- Pai, M.; Behr, M.A.; Dowdy, D.; Dheda, K.; Divangahi, M.; Boehme, C.C.; Ginsberg, A.; Swaminathan, S.; Spigelman, M.; Getahun, H.; et al. Tuberculosis. Nat. Rev. Dis. Primers 2016, 2, 16076. [Google Scholar] [CrossRef] [PubMed]
- Koul, A.; Arnoult, E.; Lounis, N.; Guillemont, J.; Andries, K. The challenge of new drug discovery for tuberculosis. Nature 2011, 469, 483–490. [Google Scholar] [CrossRef]
- Zwerling, A.; Behr, M.A.; Verma, A.; Brewer, T.F.; Menzies, D.; Pai, M. The BCG world atlas: A database of global BCG vaccination policies and practices. PLoS Med. 2011, 8, e1001012. [Google Scholar] [CrossRef] [PubMed]
- Huebner, R.E. BCG vaccination in the control of tuberculosis. Tuberculosis 1996, 215, 263–282. [Google Scholar]
- Ogura, T.; Hiraga, S. Mini-F plasmid genes that couple host cell division to plasmid proliferation. Proc. Natl. Acad. Sci. USA 1983, 80, 4784–4788. [Google Scholar] [CrossRef] [PubMed]
- Gerdes, K.; Christensen, S.K.; Løbner-Olesen, A. Prokaryotic toxin–antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
- Pandey, D.P. Toxin-antitoxin loci are highly abundant in free-living but lost from host-associated prokaryotes. Nucleic Acids Res. 2005, 33, 966–976. [Google Scholar] [CrossRef]
- Rosenberg, S.M.; Van Melderen, L.; Saavedra De Bast, M. Bacterial toxin–antitoxin systems: More than selfish entities? PLoS Genet. 2009, 5, e1000437. [Google Scholar] [CrossRef]
- Jurenas, D.; Fraikin, N.; Goormaghtigh, F.; Van Melderen, L. Biology and evolution of bacterial toxin-antitoxin systems. Nat. Rev. Microbiol. 2022, 20, 335–350. [Google Scholar] [CrossRef]
- Sala, A.; Bordes, P.; Genevaux, P. Multiple toxin-antitoxin systems in Mycobacterium tuberculosis. Toxins 2014, 6, 1002–1020. [Google Scholar] [CrossRef]
- Singh, G.; Yadav, M.; Ghosh, C.; Rathore, J.S. Bacterial toxin-antitoxin modules: Classification, functions, and association with persistence. Curr. Res. Microb. Sci. 2021, 2, 100047. [Google Scholar] [CrossRef] [PubMed]
- Sonika, S.; Singh, S.; Mishra, S.; Verma, S. Toxin-antitoxin systems in bacterial pathogenesis. Heliyon 2023, 9, e14220. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Arora, G.; Singh, M.; Kidwai, S.; Narayan, O.P.; Singh, R. MazF ribonucleases promote Mycobacterium tuberculosis drug tolerance and virulence in guinea pigs. Nat. Commun. 2015, 6, 6059. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Pati, S.; Kaushik, H.; Singh, S.; Garg, L.C. Toxin-antitoxin systems and their medical applications: Current status and future perspective. Appl. Microbiol. Biot. 2021, 105, 1803–1821. [Google Scholar] [CrossRef]
- Qiu, J.; Zhai, Y.; Wei, M.; Zheng, C.; Jiao, X. Toxin-antitoxin systems: Classification, biological roles, and applications. Microbiol. Res. 2022, 264, 127159. [Google Scholar] [CrossRef]
- Tran Van Nhieu, G.; Norton, J.P.; Mulvey, M.A. Toxin-Antitoxin systems are important for niche-specific colonization and stress resistance of uropathogenic Escherichia coli. PLoS Pathog. 2012, 8, e1002954. [Google Scholar] [CrossRef]
- Ren, D.; Walker, A.N.; Daines, D.A. Toxin-antitoxin loci vapBC-1 and vapXD contribute to survival and virulence in nontypeable Haemophilus influenzae. BMC Microbiol. 2012, 12, 263. [Google Scholar] [CrossRef]
- Zhu, L.; Inoue, K.; Yoshizumi, S.; Kobayashi, H.; Zhang, Y.; Ouyang, M.; Kato, F.; Sugai, M.; Inouye, M. Staphylococcus aureus MazF specifically cleaves a pentad sequence, UACAU, which is unusually abundant in the mRNA for pathogenic adhesive factor SraP. J. Bacteriol. 2009, 191, 3248–3255. [Google Scholar] [CrossRef] [PubMed]
- Vadivelu, J.; Wang, Y.; Wang, H.; Hay, A.J.; Zhong, Z.; Zhu, J.; Kan, B. Functional RelBE-family toxin-antitoxin pairs affect biofilm maturation and intestine colonization in Vibrio cholerae. PLoS ONE 2015, 10, e0135696. [Google Scholar] [CrossRef]
- Yu, X.; Gao, X.; Zhu, K.; Yin, H.; Mao, X.; Wojdyla, J.A.; Qin, B.; Huang, H.; Wang, M.; Sun, Y.-C.; et al. Characterization of a toxin-antitoxin system in Mycobacterium tuberculosis suggests neutralization by phosphorylation as the antitoxicity mechanism. Commun. Biol. 2020, 3, 216. [Google Scholar] [CrossRef]
- Shao, Y.; Harrison, E.M.; Bi, D.; Tai, C.; He, X.; Ou, H.-Y.; Rajakumar, K.; Deng, Z. TADB: A web-based resource for type 2 toxin–antitoxin loci in bacteria and archaea. Nucleic Acids Res. 2011, 39, D606–D611. [Google Scholar] [CrossRef] [PubMed]
- Kumari, K.; Sarma, S.P. Structural and mutational analysis of MazE6-operator DNA complex provide insights into autoregulation of toxin-antitoxin systems. Commun. Biol. 2022, 5, 963. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhang, Y.; Teh, J.S.; Zhang, J.; Connell, N.; Rubin, H.; Inouye, M. Characterization of mRNA interferases from Mycobacterium tuberculosis. J. Biol. Chem. 2006, 281, 18638–18643. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Minami, S.; Rubin, E.; Lewis, K.; Davies, J.E. Characterization and transcriptome analysis of Mycobacterium tuberculosis persisters. mBio 2011, 2, e00100–e00111. [Google Scholar] [CrossRef]
- Kazemian, H.; Heidari, H.; Kardan-Yamchi, J.; Ghafourian, S.; Pakzad, I.; Kouhsari, E.; Valadbeigi, H.; Sadeghifard, N. Comparison of toxin-antitoxin expression among drug-susceptible and drug-resistant clinical isolates of Mycobacterium tuberculosis. Adv. Respir. Med. 2021, 89, 110–114. [Google Scholar] [CrossRef]
- Wayne, L.G. Dormancy of Mycobacterium tuberculosis and latency of disease. Eur. J. Clin. Microbiol. 1994, 13, 908–914. [Google Scholar] [CrossRef]
- Ramirez, M.V.; Dawson, C.C.; Crew, R.; England, K.; Slayden, R.A. MazF6 toxin of Mycobacterium tuberculosis demonstrates antitoxin specificity and is coupled to regulation of cell growth by a Soj-like protein. BMC Microbiol. 2013, 13, 240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Hara, H.; Kato, I.; Inouye, M. Insights into the mRNA cleavage mechanism by MazF, an mRNA Interferase. J. Biol. Chem. 2005, 280, 3143–3150. [Google Scholar] [CrossRef]
- Zhu, L.; Phadtare, S.; Nariya, H.; Ouyang, M.; Husson, R.N.; Inouye, M. The mRNA interferases, MazF-mt3 and MazF-mt7 from Mycobacterium tuberculosis target unique pentad sequences in single-stranded RNA. Mol. Microbiol. 2008, 69, 559–569. [Google Scholar] [CrossRef]
- Schifano, J.M.; Vvedenskaya, I.O.; Knoblauch, J.G.; Ouyang, M.; Nickels, B.E.; Woychik, N.A. An RNA-seq method for defining endoribonuclease cleavage specificity identifies dual rRNA substrates for toxin MazF-mt3. Nat. Commun. 2014, 5, 3538. [Google Scholar] [CrossRef]
- Simanshu, D.K.; Yamaguchi, Y.; Park, J.-H.; Inouye, M.; Patel, D.J. Structural basis of mRNA recognition and cleavage by toxin MazF and its regulation by antitoxin MazE in Bacillus subtilis. Mol. Cell 2013, 52, 447–458. [Google Scholar] [CrossRef]
- Zorzini, V.; Mernik, A.; Lah, J.; Sterckx, Y.G.J.; De Jonge, N.; Garcia-Pino, A.; De Greve, H.; Versées, W.; Loris, R. Substrate recognition and activity regulation of the Escherichia coli mRNA endonuclease MazF. J. Biol. Chem. 2016, 291, 10950–10960. [Google Scholar] [CrossRef]
- Zorzini, V.; Haesaerts, S.; Cheung, A.; Loris, R.; van Nuland, N.A.J. 1H, 13C, and 15N backbone and side-chain chemical shift assignment of the staphylococcal MazF mRNA interferase. Biomol. NMR Assign. 2011, 5, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Zorzini, V.; Buts, L.; Sleutel, M.; Garcia-Pino, A.; Talavera, A.; Haesaerts, S.; Greve, H.D.; Cheung, A.; van Nuland, N.A.J.; Loris, R. Structural and biophysical characterization of Staphylococcus aureus SaMazF shows conservation of functional dynamics. Nucleic Acids Res. 2014, 42, 6709–6725. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhou, J.; Sun, R.; Du, C.; Xie, W. Conserved conformational changes in the regulation of Mycobacterium tuberculosis MazEF-mt1. ACS Infect. Dis. 2020, 6, 1783–1795. [Google Scholar] [CrossRef]
- Chen, R.; Zhou, J.; Xie, W. Mechanistic insight into the peptide binding modes to two M. tb MazF toxins. Toxins 2021, 13, 319. [Google Scholar] [CrossRef]
- Ahn, D.H.; Lee, K.Y.; Lee, S.J.; Park, S.J.; Yoon, H.J.; Kim, S.J.; Lee, B.J. Structural analyses of the MazEF4 toxin-antitoxin pair in Mycobacterium tuberculosis provide evidence for a unique extracellular death factor. J. Biol. Chem. 2017, 292, 18832–18847. [Google Scholar] [CrossRef] [PubMed]
- Santos-Sierra, S.; Pardo-Abarrio, C.; Giraldo, R.; Díaz-Orejas, R. Genetic identification of two functional regions in the antitoxin of the parD killer system of plasmid R1. FEMS Microbiol. Lett. 2002, 206, 115–119. [Google Scholar] [CrossRef]
- Smith, J.A.; Magnuson, R.D. Modular organization of the Phd repressor/antitoxin protein. J. Bacteriol. 2004, 186, 2692–2698. [Google Scholar] [CrossRef]
- Bernard, P.; Couturier, M. The 41 carboxy-terminal residues of the miniF plasmid CcdA protein are sufficient to antagonize the killer activity of the CcdB protein. Mol. Gen. Genet. 1991, 226, 297–304. [Google Scholar] [CrossRef]
- Chen, R.; Tu, J.; Liu, Z.; Meng, F.; Ma, P.; Ding, Z.; Yang, C.; Chen, L.; Deng, X.; Xie, W. Structure of the MazF-mt9 toxin, a tRNA-specific endonuclease from Mycobacterium tuberculosis. Biochem. Bioph. Res. Co. 2017, 486, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Tu, J.; Tan, Y.; Cai, X.; Yang, C.; Deng, X.; Su, B.; Ma, S.; Liu, X.; Ma, P.; et al. Structural and biochemical characterization of the cognate and heterologous interactions of the MazEF-mt9 TA system. ACS Infect. Dis. 2019, 5, 1306–1316. [Google Scholar] [CrossRef]
- Hoffer, E.D.; Miles, S.J.; Dunham, C.M. The structure and function of Mycobacterium tuberculosis MazF-mt6 toxin provide insights into conserved features of MazF endonucleases. J. Biol. Chem. 2017, 292, 7718–7726. [Google Scholar] [CrossRef]
- Steyaert, J. A decade of protein engineering on ribonuclease T1 atomic dissection of the enzyme—Substrate interactions. Eur. J. Biochem. 2004, 247, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Schifano, J.M.; Edifor, R.; Sharp, J.D.; Ouyang, M.; Konkimalla, A.; Husson, R.N.; Woychik, N.A. Mycobacterial toxin MazF-mt6 inhibits translation through cleavage of 23S rRNA at the ribosomal A site. Proc. Natl. Acad. Sci. USA 2013, 110, 8501–8506. [Google Scholar] [CrossRef] [PubMed]
- Minor, W.; Cymborowski, M.; Otwinowski, Z.; Chruszcz, M. HKL-3000: The integration of data reduction and structure solution—From diffraction images to an initial model in minutes. Acta Crystallogr. Sect. D Biol. Crystallogr. 2006, 62, 859–866. [Google Scholar] [CrossRef]
- Minor, W.; Cymborowski, M.; Borek, D.; Cooper, D.R.; Chruszcz, M.; Otwinowski, Z. Optimal structure determination from sub-optimal diffraction data. Protein Sci. 2021, 31, 259–268. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2009, 66, 12–21. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MazF-mt3-Apo (9IKD) | MazF-mt3-MazE (8ZWS) | |
|---|---|---|
| Data Collection | ||
| Space group | P43212 | P3121 |
| Cell dimensions | ||
| a, b, c (Å) | 110.8, 110.8, 271.2 | 84.7, 84.7, 101.8 |
| α, β, γ (°) | 90, 90, 90 | 90, 90, 120 |
| Resolution (Å) | 75.27–3.32 (3.50–3.32) * | 73.39–3.27 (3.45–3.27) * |
| Rmerge | 0.16 (1.31) | 0.16 (0.99) |
| I/σI | 18.4 (2.8) | 14.8 (2.8) |
| Completeness (%) | 92.7 (78.8) | 94.2 (81.0) |
| Redundancy | 24.2 (21.3) | 14.1 (12.4) |
| Refinement | ||
| Resolution (Å) | 75.27–3.32 (3.47–3.32)* | 73.39–3.27 (4.12–3.27) * |
| No. reflections | 23,795 | 6445 |
| Rwork/Rfree (%) | 24.2/27.9 | 23.9/26.1 |
| No. atoms | ||
| Protein | 9020 | 2690 |
| B-factors (Å2) | ||
| Protein | 109.8 | 83.3 |
| R.m.s. deviations | ||
| Bond lengths (Å) | 0.003 | 0.002 |
| Bond angles (°) | 0.61 | 0.48 |
| Ramachandran | ||
| Favored (%) | 97.51 | 96.80 |
| Outliers (%) | 0.33 | 0 |
| Antitoxin (Chain I) | Toxin (Chain A) | Toxin (Chain B) |
|---|---|---|
| Thr47 (OG1) | Pro34′ (O) | |
| Asp51 (OD1) | Arg39′ (N) | |
| Glu55 (N) | Arg39′ (NH2) | |
| Asp61 (OD1) | Asn51 (ND2) | |
| Asp61 (OD2) | Thr49 (OG1) | |
| Glu62 (OE2) | Arg25 (NH2) | |
| Glu62 (OE2) | Arg25 (NE) | |
| Glu64 (OE1) | Asn77 (ND2) | |
| Glu64 (OE2) | Asn77 (ND2) | |
| Glu64 (OE2) | Thr79 (OG1) | |
| Asn70 (OD1) | Thr83 (N) | |
| Asn70 (ND2) | Thr83 (O) |
| TA Systems | MazE | Full-Length MazF | |||
|---|---|---|---|---|---|
| α3 | α4 | α3–α4 | Full-Length | ||
| mt1 | 3.43 | 3.70 | 3.47 | 4.37 | 9.10 |
| mt3 | 5.20 | 3.83 | 4.00 | 4.63 | 8.40 |
| mt9 | 6.80 | 3.70 | 4.44 | 5.10 | 10.40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Z.; Jiang, P.; Xie, W. Long Dynamic β1–β2 Loops in M. tb MazF Toxins Affect the Interaction Modes and Strengths of the Toxin–Antitoxin Pairs. Int. J. Mol. Sci. 2024, 25, 9630. https://doi.org/10.3390/ijms25179630
Tang Z, Jiang P, Xie W. Long Dynamic β1–β2 Loops in M. tb MazF Toxins Affect the Interaction Modes and Strengths of the Toxin–Antitoxin Pairs. International Journal of Molecular Sciences. 2024; 25(17):9630. https://doi.org/10.3390/ijms25179630
Chicago/Turabian StyleTang, Ziyun, Pengcheng Jiang, and Wei Xie. 2024. "Long Dynamic β1–β2 Loops in M. tb MazF Toxins Affect the Interaction Modes and Strengths of the Toxin–Antitoxin Pairs" International Journal of Molecular Sciences 25, no. 17: 9630. https://doi.org/10.3390/ijms25179630
APA StyleTang, Z., Jiang, P., & Xie, W. (2024). Long Dynamic β1–β2 Loops in M. tb MazF Toxins Affect the Interaction Modes and Strengths of the Toxin–Antitoxin Pairs. International Journal of Molecular Sciences, 25(17), 9630. https://doi.org/10.3390/ijms25179630