
Slot Blot- and Electrospray Ionization–Mass Spectrometry/Matrix-Assisted Laser Desorption/Ionization–Mass Spectrometry-Based Novel Analysis Methods for the Identification and Quantification of Advanced Glycation End-Products in the Urine

 , , and
, , and
Abstract
:1. Introduction
2. Clinical Urinalysis
3. Lifestyle-Related Diseases
4. Advanced Glycation End-Products
4.1. Origin of AGEs
4.2. Free-Type AGEs
4.2.1. Free-Type AGEs Containing One Amino Acid Residue
4.2.2. Free-Type AGEs Containing Two Amino Acid Residues
4.3. Crude, Diverse, and Multiple AGE Patterns
4.3.1. Crude AGE Pattern
4.3.2. Diverse AGE Patterns
4.3.3. Multiple AGE Pattern
4.3.4. Influence of Crude, Diverse, and Multiple AGE Patterns on Cells
5. AGEs and LSRDs
5.1. Intracellular AGEs and LSRDs
5.2. Extracellular AGEs and LSRDs
5.2.1. AGEs in Body Fluids
5.2.2. AGEs in the Extracellular Matrix
5.2.3. Dietary AGEs
5.2.4. Fluid AGEs as a Biomarker
5.2.5. Pentosidine in the Urine and Skin as a Biomarker for Aging, Osteoporosis, and CVD
6. Previous Methods Used for the Identification and Quantification of AGEs in the Urine
7. Novel Analysis Method Combining Slot Blot and ESI-MS/MALDI-MS for the Identification and Quantification of AGEs in Urine
7.1. Concept and Novelty
7.2. Steps Involved in the Novel Analysis Method for AGEs in Urine
7.3. Overcoming the Challenges Associated with the Collection of AGE-Modified Proteins in Urine
7.4. Slot Blotting and Analysis
7.4.1. Binding of AGE-Modified Proteins in Urine onto PVDF Membrane
7.4.2. Carbamoylation of AGE-Modified Proteins onto the PVDF Membrane

7.4.3. Cutting the PVDF Membrane
7.4.4. Slot Blot Analysis for AGE-Modified Proteins
7.5. ESI-/MALDI-MS Analysis of the AGE-Modified Proteins on the PVDF Membrane
7.5.1. ESI-/MALDI-MS in AGE Research and Challenges Associated with Their Use for Detecting AGE-Modified Proteins
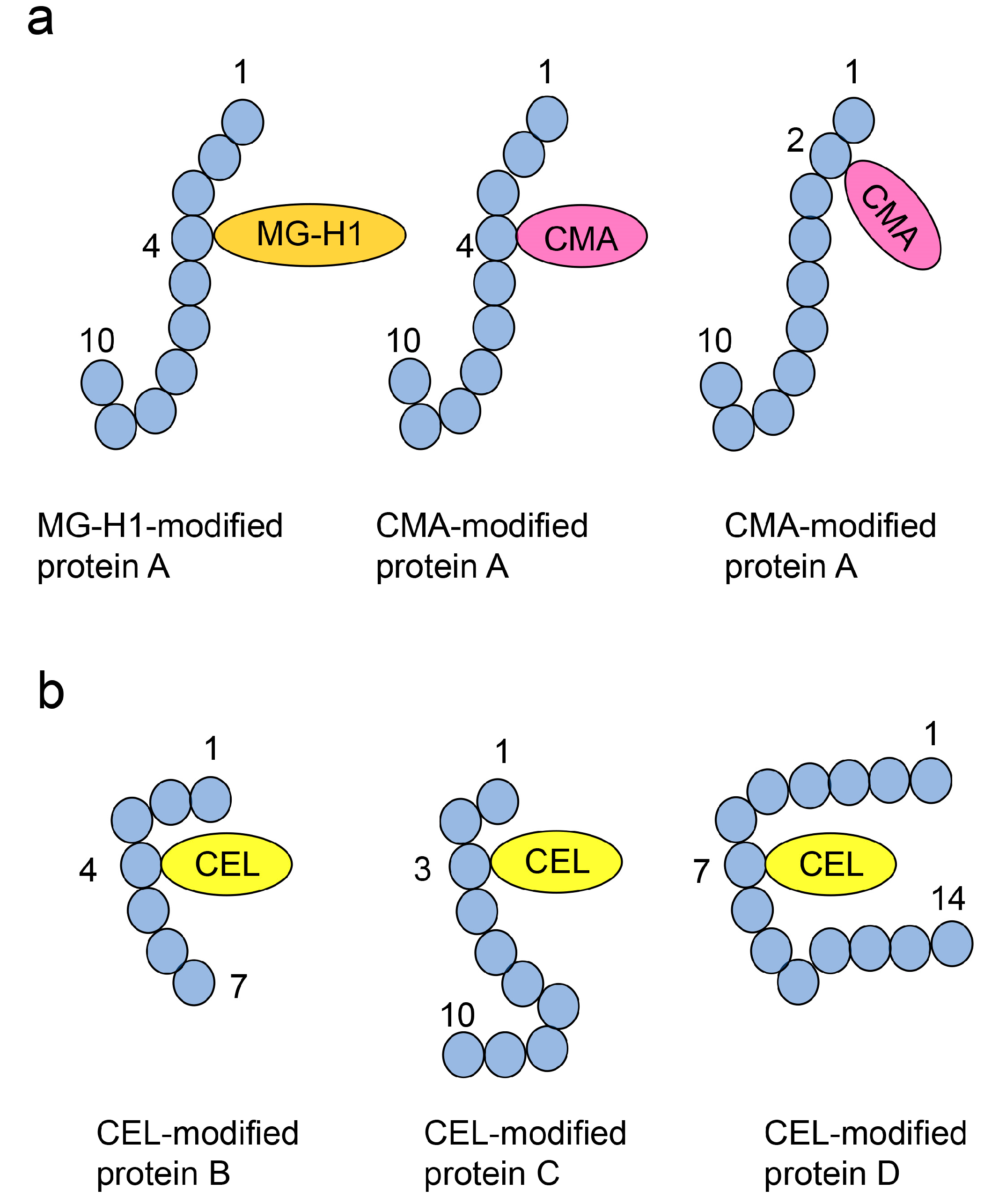
7.5.2. ESI-/MALDI-MS-Based Analysis of Free-Type AGEs after On-Membrane Acid Hydrolysis
7.5.3. ESI-/MALDI-MS Analysis of AGE-Modified Peptides after On-Membrane Enzymatic Digestion
7.6. Comparison of Existing Methods and Our Novel Method for the Identification and Quantification of AGEs in the Urine
7.7. Advantages of Using Three Methods to Analyze AGEs
7.8. Desirable Practical Clinical Applications
7.9. Limitations of the Novel Strategy
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AGEs | Advanced glycation end-products |
| Arg-P | Argpyrimidine |
| CHAPS | 3-[3-(cholamidopropyl)-dimethylammonio]-1-propanesulfonate |
| CEL | Nε-carboxyethyl-lysine |
| CMA | Nω-carboxymethyl-arginine |
| CML | Nε-carboxymethyl-lysine |
| CVD | Cardiovascular disease |
| DM | Diabetes mellitus |
| DODIC | 3-Deoxyglucospane-derived imidazolium cross-link |
| ELISA | Enzyme-linked immunosorbent assay |
| ESI-MS | Electrospray ionization–mass spectrometry |
| GA-AGEs | Glyceraldehyde-derived advanced glycation end-products |
| GC-MS | Gas chromatography–mass spectrometry |
| G-H1 | Glyoxal-derived hydroimidazolone |
| GLAP | Glyceraldehyde-derived pyridinium |
| GLO | Glyoxalase |
| GO-AGEs | Glyoxal-derived advanced glycation end products |
| GODIC | Glyoxal-derived imidazolium cross-link |
| GOLA | Nε-{2-[(5-Amino-5-carboxypentyl)-amino]-2-oxoethyl}-lysine |
| HPLC | High-performance liquid chromatography |
| HPLC-ESI-MS | High-performance liquid chromatography electrospray ionization–mass spectrometry |
| LC | Liquid chromatography |
| LC-ESI-MS | Liquid chromatography electrospray ionization–mass spectrometry |
| LSRD | Lifestyle-related disease |
| MAGE | Melibiose-derived advanced glycation end-products |
| MALDI-MS | Matrix-assisted laser desorption–mass spectrometry |
| MG-H1 | Nδ-(5-hydro-5-methyl-4-imidazolone-2-yl)-ornithine |
| MGO-AGEs | Methylglyoxal-derived advanced glycation end-products |
| MODIC | Methylglyoxal-derived imidazolium cross-link |
| MOLD | Methylglyoxal-lysine dimer |
| NASH | Non-alcoholic steatohepatitis |
| NMR | Nuclear magnetic resonance |
| PPG | Pyrrolopyridinium lysine dimer derived from glyceraldehyde |
| RAGE | Receptor for advanced glycation end-products |
| TAGE | Toxic advanced glycation end-products |
| TLR4 | Tool-like receptor 4 |
| Tris | 3-[3-(Cholamidepropyl)-tris-(hydroxymethyl)-aminomethane |
| T2R | Bitter taste receptor |
References
- Thakkar, R.N.; Kim, D.; Knight, A.M.; Riedel, S.; Vaidya, D.; Wright, S.M. Impact of an educational intervention on the frequency of daily blood test orders for hospitalized patients. Am. J. Clin. Pathol. 2015, 143, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yi, Y.; Luo, X.; Xiong, N.; Liu, Y.; Li, S.; Sun, R.; Wang, Y.; Hu, B.; Chen, W.; et al. Development and clinical application of a rapid IgM-IgG combined antibody test for SARS-CoV-2 infection diagnosis. J. Med. Virol. 2020, 92, 1518–1524. [Google Scholar] [CrossRef] [PubMed]
- Garraway, L.A.; Lander, E.S. Lessons from the cancer genome. Cell 2013, 153, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Twomey, J.D.; Zhang, B. Cancer immunotherapy update: FDA-approved checkpoint inhibitors and companion diagnostics. AAPS J. 2021, 23, 39. [Google Scholar] [CrossRef]
- Geyer, P.E.; Voytik, E.; Treit, P.V.; Doll, S.; Kleinhempel, A.; Niu, L.; Müller, J.B.; Buchholtz, M.L.; Bader, J.M.; Teupser, D.; et al. Plasma Proteome Profiling to detect and avoid sample-related biases in biomarker studies. EMBO Mol. Med. 2019, 11, e10427. [Google Scholar] [CrossRef]
- Froesch, E.R. Specific enzymatic determination of glucose in blood and urine using glucose oxidase. Science 1956, 125, 1082–1083. [Google Scholar] [CrossRef]
- van Rhijn, B.W.G.; van der Poel, H.G.; van der Kwast, T.H. Urine markers for bladder cancer surveillance: A systematic review. Eur. Urol. 2005, 47, 736–748. [Google Scholar] [CrossRef]
- Waikar, S.S. Biomarker blues: Balancing hope and hype in acute kidney injury. Kidney Int. 2024, 105, 679–682. [Google Scholar] [CrossRef]
- Kuhara, T. Gas chromatographic–mass spectrometric urinary metabolome analysis to study mutations of inborn errors of metabolism. Mass Spectrom. Rev. 2005, 24, 814–827. [Google Scholar] [CrossRef]
- Yasui, T.; Yanagida, T.; Ito, S.; Konakade, Y.; Takeshita, D.; Naganawa, T.; Nagashima, K.; Shimada, T.; Kaji, N.; Nakamura, Y.; et al. Unveiling massive numbers of cancer-related urinary-microRNA candidates via nanowires. Sci. Adv. 2017, 3, e1701133. [Google Scholar] [CrossRef]
- Jaberi, S.A.; Cohen, A.; D’Souza, C.; Abdulrazzaq, Y.M.; Ojha, S.; Bastaki, S.; Adeghate, E.A. Lipocalin-2: Structure, function, distribution and role in metabolic disorders. Biomed. Pharmacother. 2021, 142, 112002. [Google Scholar] [CrossRef]
- Mori, Y.; Ajay, A.K.; Chang, J.H.; Mou, S.; Zhao, H.; Kishi, S.; Li, J.; Brooks, C.R.; Xiao, S.; Woo, H.M.; et al. KIM-1 mediates fatty acid uptake by renal tubular cells to promote progressive diabetic kidney disease. Cell Metab. 2021, 33, 1042–1061.e7. [Google Scholar] [CrossRef] [PubMed]
- Kamijo, A.; Sugaya, T.; Hikawa, A.; Yamanouchi, M.; Hirata, Y.; Ishimitsu, T.; Numabe, A.; Takagi, M.; Hayakawa, H.; Tabei, F.; et al. Urinary liver-type fatty acid binding protein as a useful biomarker in chronic kidney disease. Mol. Cell. Biochem. 2006, 284, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; He, G.; Goh, S.; Low, A.W.X.; Tay, K.J.; Lim, T.K.H.; Yeong, J.; Khor, L.Y.; Lim, T.S. Biomarkers for precision urothelial carcinoma diagnosis: Current approaches and the application of single-cell technologies. Cancers 2021, 13, 260. [Google Scholar] [CrossRef] [PubMed]
- Rose, K.M.; Huelster, H.L.; Meeks, J.J.; Faltas, B.M.; Sonpavde, G.P.; Lerner, S.P.; Ross, J.S.; Spiess, P.E.; Grass, G.D.; Jain, R.K.; et al. Circulating and urinary tumour DNA in urothelial carcinoma-upper tract, lower tract and metastatic disease. Nat. Rev. Urol. 2023, 20, 406–419. [Google Scholar] [CrossRef]
- Gug, I.T.; Tertis, M.; Hosu, O.; Cristea, C. Salivary biomarkers detection: Analytical and immunological methods overview. TrAC Trends Anal. Chem. 2019, 113, 301–316. [Google Scholar] [CrossRef]
- Dawes, C.; Wong, D.T.W. Role of saliva and salivary diagnostics in the advancement of Oral Health. J. Dent. Res. 2019, 98, 133–141. [Google Scholar] [CrossRef]
- Butler-Laporte, G.; Lawandi, A.; Schiller, I.; Yao, M.; Dendukuri, N.; McDonald, E.G.; Lee, T.C. Comparison of saliva and nasopharyngeal swab nucleic acid amplification testing for detection of SARS-CoV-2: A systematic review and meta-analysis. JAMA Intern. Med. 2021, 181, 353–360. [Google Scholar] [CrossRef]
- Nonaka, T.; Wong, D.T.W. Saliva diagnostics: Salivaomics, saliva exosomics, and saliva liquid biopsy. J. Am. Dent. Assoc. 2023, 154, 696–704. [Google Scholar] [CrossRef]
- Laxton, C.S.; Peno, C.; Hahn, A.M.; Allicock, O.M.; Perniciaro, S.; Wyllie, A.L. The potential of saliva as an accessible and sensitive sample type for the detection of respiratory pathogens and host immunity. Lancet Microbe 2023, 4, e837–e850. [Google Scholar] [CrossRef]
- Moravcová, K.; Sovová, M.; Ožana, J.; Karbanová, M.; Klásek, J.; Kolasińska, A.B.; Sovová, E. Comparing the efficacy of digital and in-person weight loss interventions for patients with obesity and glycemic disorders: Evidence from a randomized non-inferiority trial. Nutrients 2024, 16, 1510. [Google Scholar] [CrossRef] [PubMed]
- Shu, M.; Xi, Y.; Wu, J.; Zhuo, L.B.; Yan, Y.; Yang, Y.D.; Feng, Y.Y.; Tan, H.Q.; Yang, H.F.; Chen, Y.M. Relationship between circulating 25-hydroxyvitamin D and metabolic syndrome in Chinese adults: A large nationwide longitudinal study. Nutrients 2024, 16, 1480. [Google Scholar] [CrossRef] [PubMed]
- Nugnes, M.; Baldassarre, M.; Ribichini, D.; Tedesco, D.; Capelli, I.; Vetrano, D.; Marchignoli, F.; Brodosi, L.; Pompili, E.; Petroni, M.L.; et al. Association between albumin alterations and renal function in patients with type 2 diabetes mellitus. Int. J. Mol. Sci. 2024, 25, 3168. [Google Scholar] [CrossRef] [PubMed]
- Jun, L.; Ding, X.W.; Robinson, M.; Jafari, H.; Knight, E.; Geetha, T.; Greene, M.W.; Babu, J.R. Targeting molecular mechanisms of obesity- and type 2 diabetes mellitus-induced skeletal muscle atrophy with nerve growth factor. Int. J. Mol. Sci. 2024, 25, 4307. [Google Scholar] [CrossRef]
- Reed, J.; Higginbotham, V.; Bain, S.; Kanamarlapudi, V. Comparative analysis of orthosteric and allosteric GLP-1R agonists’ effects on insulin secretion from healthy, diabetic, and recovered INS-1E pancreatic beta cells. Int. J. Mol. Sci. 2024, 25, 6331. [Google Scholar] [CrossRef]
- Barchetta, I.; Cimini, F.A.; Sentinelli, F.; Chiappetta, C.; Di Cristofano, C.D.; Silecchia, G.; Leonetti, F.; Baroni, M.G.; Cavallo, M.G. Reduced lipopolysaccharide-binding protein (LBP) levels are associated with non-alcoholic fatty liver disease (NAFLD) and adipose inflammation in human obesity. Int. J. Mol. Sci. 2023, 24, 17174. [Google Scholar] [CrossRef]
- Mouskeftara, T.; Kalopitas, G.; Liapikos, T.; Arvanitakis, K.; Germanidis, G.; Gika, H. Predicting non-alcoholic steatohepatitis: A lipidomics-driven machine learning approach. Int. J. Mol. Sci. 2024, 25, 5965. [Google Scholar] [CrossRef]
- Mphekgwana, P.M.; Monyeki, K.D.; Mothiba, T.M.; Makgahlela, M.; Kgatla, N.; Malema, R.N.; Sodi, T. Screening and interventions for cardiovascular disease prevention in the Limpopo Province, South Africa: Use of the community action model. Metabolites 2022, 12, 1067. [Google Scholar] [CrossRef]
- Stančáková Yaluri, A.S.; Tkáč, I.; Tokarčíková, K.; Kozelová, Z.; Rašiová, M.; Javorský, M.; Kozárová, M. Decreased 25-hydroxy vitamin D Level is associated with all-cause mortality in patients with type 2 diabetes at high cardiovascular risk. Metabolites 2023, 13, 887. [Google Scholar] [CrossRef]
- Ou-Yang, Y.N.; Yuan, M.D.; Yang, Z.M.; Min, Z.; Jin, Y.X.; Tian, Z.M. Revealing the pathogenesis of salt-sensitive hypertension in dahl salt-sensitive rats through integrated multi-omics analysis. Metabolites 2022, 12, 1076. [Google Scholar] [CrossRef]
- Genovesi, S.; Montelisciani, L.; Giussani, M.; Lieti, G.; Patti, I.; Orlando, A.; Antolini, L.; Parati, G. Role of insulin resistance as a mediator of the relationship between body weight, waist circumference, and systolic blood pressure in a pediatric population. Metabolites 2023, 13, 327. [Google Scholar] [CrossRef]
- Yu, W.C.; Huang, R.Y.; Chou, T.C. Oligo-fucoidan improves diabetes-induced renal fibrosis via activation of Sirt-1, GLP-1R, and Nrf2/HO-1: An in vitro and in vivo study. Nutrients 2020, 12, 3068. [Google Scholar] [CrossRef]
- Hammad, F.T.; Al-Salam, S.; Ahmad, R.; Yasin, J.; Hammad, A.F.; Rasheed, J.A.; Lubbad, L. The Effect of nerolidol Renal Dysfunction following ischemia-reperfusion Injury in the Rat. Nutrients 2023, 15, 455. [Google Scholar] [CrossRef] [PubMed]
- Shiraki, M.; Kuroda, T.; Tanaka, S.; Saito, M.; Fukunaga, M.; Nakamura, T. Nonenzymatic collagen cross-links induced by glycoxidation (pentosidine) predicts vertebral fractures. J. Bone Miner. Metab. 2008, 26, 93–100. [Google Scholar] [CrossRef]
- Kida, Y.; Saito, M.; Shinohara, A.; Soshi, S.; Marumo, K. Non-invasive skin autofluorescence, blood and urine assays of the advanced glycation end product (AGE) pentosidine as an indirect indicator of AGE content in human bone. B.M.C. Musculoskelet. Disord. 2019, 20, 627. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.A.; DeGroot, J.; Huisman, A.M.; Oostveen, J.C.; Marijnissen, A.C.; Bijlsma, J.W.; van El, B.; Zuurmond, A.M.; Lafeber, F.P. Skin and urine pentosidine weakly correlate with joint damage in a cohort of patients with early signs of osteoarthritis (CHECK). Osteoarthr. Cartil. 2010, 18, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Vos, P.A.; Mastbergen, S.C.; Huisman, A.M.; de Boer, T.N.; Degroot, J.; Polak, A.A.; Lafeber, F.P.J.G. In end stage osteoarthritis, cartilage tissue pentosidine levels are inversely related to parameters of cartilage damage. Osteoarthr. Cartil. 2012, 20, 233–240. [Google Scholar] [CrossRef]
- Twarda-Clapa, A.; Olczak, A.; BiałKowska, A.M.; KoziołKiewicz, M. Advanced glycation end-products (AGEs): Formation, chemistry, classification, receptors, and diseases related to AGEs. Cells 2022, 11, 1312. [Google Scholar] [CrossRef]
- Takata, T.; Motoo, Y. Novel in vitro assay of the effects of Kampo medicines against intra/extracellular advanced glycation end-products in oral, esophageal, and gastric epithelial cells. Metabolites 2023, 13, 878. [Google Scholar] [CrossRef]
- Baskal, S.; Büttner, P.; Werner, S.; Besler, C.; Lurz, P.; Thiele, H.; Tsikas, D. Profile of urinary amino acids and their post-translational modifications (PTM) including advanced glycation end-products (AGEs) of lysine, arginine and cysteine in lean and obese ZSF1 rats. Amino Acids 2022, 54, 643–652. [Google Scholar] [CrossRef]
- Indyk, D.; Bronowicka-Szydełko, A.; Gamian, A.; Kuzan, A. Advanced glycation end products and their receptors in serum of patients with type 2 diabetes. Sci. Rep. 2021, 11, 13264. [Google Scholar] [CrossRef]
- Mastrocola, R.; Collino, M.; Nigro, D.; Chiazza, F.; D’Antona, G.; Aragno, M.; Minetto, M.A. Accumulation of advanced glycation end-products and activation of the SCAP/SREBP lipogenetic pathway occur in diet-induced obese mouse skeletal muscle. PLoS ONE 2015, 10, e0119587. [Google Scholar] [CrossRef] [PubMed]
- Mastrocola, R.; Nigro, D.; Chiazza, F.; Medana, C.; Dal Bello, F.; Boccuzzi, G.; Collino, M.; Aragno, M. Fructose-derived advanced glycation end-products drive lipogenesis and skeletal muscle reprogramming via SREBP-1c dysregulation in mice. Free Radic. Biol. Med. 2016, 91, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Litwinowicz, K.; Waszczuk, E.; Kuzan, A.; Bronowicka-Szydełko, A.; Gostomska-Pampuch, K.; Naporowski, P.; Gamian, A. Alcoholic liver disease is associated with elevated plasma levels of novel advanced glycation end-products: A preliminary study. Nutrients 2022, 14, 5266. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, M.; Holewinski, R.J.; Previs, S.B.; Martin, T.G.; Stachowski, M.J.; Li, A.; Blair, C.A.; Moravec, C.S.; Van Eyk, J.E.; Campbell, K.S.; et al. Diabetes with heart failure increases methylglyoxal modifications in the sarcomere, which inhibit function. JCI Insight 2018, 3, e121264. [Google Scholar] [CrossRef]
- Papadaki, M.; Kampaengsri, T.; Barrick, S.K.; Campbell, S.G.; von Lewinski, D.; Rainer, P.P.; Harris, S.P.; Greenberg, M.J.; Kirk, J.A. Myofilament glycation in diabetes reduces contractility by inhibiting tropomyosin movement, is rescued by cMyBPC domains. J. Mol. Cell. Cardiol. 2022, 162, 1–9. [Google Scholar] [CrossRef]
- Ruiz-Meana, M.; Minguet, M.; Bou-Teen, D.; Miro-Casas, E.; Castans, C.; Castellano, J.; Bonzon-Kulichenko, E.; Igual, A.; Rodriguez-Lecoq, R.; Vázquez, J.; et al. Ryanodine receptor glycation favors mitochondrial damage in the senescent heart. Circulation 2019, 139, 949–964. [Google Scholar] [CrossRef]
- Kashiwabara, S.; Hosoe, H.; Ohno, R.I.; Nagai, R.; Shiraki, M. Development and evaluation of novel ELISA for determination of urinary pentosidine. J. Nutr. Sci. Vitaminol. 2019, 65, 526–533. [Google Scholar] [CrossRef]
- Takata, T.; Inoue, S.; Masauji, T.; Miyazawa, K.; Motoo, Y. Generation and accumulation of various advanced glycation end-products in cardiomyocytes may induce cardiovascular disease. Int. J. Mol. Sci. 2024, 25, 7319. [Google Scholar] [CrossRef]
- Takata, T. Is the novel slot blot a useful method for quantification of intracellular advanced glycation end-products? Metabolites 2023, 13, 564. [Google Scholar] [CrossRef]
- Takata, T.; Masauji, T.; Motoo, Y. Analysis of crude, diverse, and multiple advanced glycation end-product patterns may be important and beneficial. Metabolites 2024, 14, 3. [Google Scholar] [CrossRef]
- Jung, W.K.; Park, S.B.; Kim, H.R.; Ryu, H.Y.; Kim, Y.H.; Kim, J. Advanced glycation end products increase salivary gland hypofunction in d-galactose-induced aging rats and its prevention by physical exercise. Curr. Issues Mol. Biol. 2021, 43, 2059–2067. [Google Scholar] [CrossRef]
- Damasiewicz-Bodzek, A.; Łabuz-Roszak, B.; Kumaszka, B.; Tadeusiak, B.; Tyrpień-Golder, K. The assessment of serum concentrations of AGEs and their soluble receptor (sRAGE) in multiple sclerosis patients. Brain Sci. 2021, 11, 1021. [Google Scholar] [CrossRef] [PubMed]
- Baskal, S.; Kaiser, A.; Mels, C.; Kruger, R.; Tsikas, D. Specific and sensitive GC-MS analysis of hypusine, Nε-(4-amino-2-hydroxybutyl)lysine, a biomarker of hypusinated eukaryotic initiation factor eIF5A, and its application to the bi-ethnic ASOS study. Amino Acids 2022, 54, 1083–1099. [Google Scholar] [CrossRef] [PubMed]
- Baskal, S.; Bollenbach, A.; Mels, C.; Kruger, R.; Tsikas, D. Development, validation of a GC-MS method for the simultaneous measurement of amino acids, their PTM metabolites and AGEs in human urine, and application to the bi-ethnic ASOS study with special emphasis to lysine. Amino Acids 2022, 54, 615–641. [Google Scholar] [CrossRef]
- Wada, K.; Nakashima, Y.; Yamakawa, M.; Hori, A.; Seishima, M.; Tanabashi, S.; Matsushita, S.; Tokimitsu, N.; Nagata, C. Dietary advanced glycation end products and cancer risk in Japan: From the Takayama study. Cancer Sci. 2022, 113, 2839–2848. [Google Scholar] [CrossRef] [PubMed]
- Phuong-Nguyen, K.; McNeill, B.A.; Aston-Mourney, K.; Rivera, L.R. Advanced glycation end-products and their effects on gut health. Nutrients 2023, 15, 405. [Google Scholar] [CrossRef]
- Lin, K.; Deng, T.; Qu, H.; Ou, H.; Huang, Q.; Gao, B.; Li, X.; Wei, N. Gastric protective effect of Alpinia officinarum flavonoids: Mediating TLR4/NF-κB and TRPV1 signalling pathways and gastric mucosal healing. Pharm. Biol. 2023, 61, 50–60. [Google Scholar] [CrossRef]
- González-Guerrero, D.E.; Lazo-de-la-Vega-Monroy, M.L.; Gómez-Ojeda, A.; Luévano-Contreras, C.; Rojas-Rubio, A.; Garay-Sevilla, M.E. Polymorphisms −374 T/A and −429 T/C of the receptor for advanced glycation end-products (RAGE) and serum levels of RAGE (sRAGE) are not associated with metabolic syndrome. Metabolites 2023, 13, 521. [Google Scholar] [CrossRef]
- Hayashi, K.; Sato, K.; Ochi, S.; Kawano, S.; Munesue, S.; Harashima, A.; Oshima, Y.; Kimura, K.; Kyoi, T.; Yamamoto, Y. Inhibitory effects of Saururus chinensis extract on receptor for advanced glycation end-products-dependent inflammation and diabetes-induced dysregulation of vasodilation. Int. J. Mol. Sci. 2022, 23, 5757. [Google Scholar] [CrossRef]
- Lee, H.W.; Gu, M.J.; Lee, J.Y.; Lee, S.; Kim, Y.; Ha, S.K. Methylglyoxal-lysine dimer, an advanced glycation end product, Induces inflammation via interaction with RAGE in mesangial cells. Mol. Nutr. Food Res. 2021, 65, e2000799. [Google Scholar] [CrossRef] [PubMed]
- Palaseweenun, P.; Hagen-Plantinga, E.A.; Schonewille, J.T.; Koop, G.; Butre, C.; Jonathan, M.; Wierenga, P.A.; Hendriks, W.H. Urinary excretion of advanced glycation end products in dogs and cats. J. Anim. Physiol. Anim. Nutr. 2021, 105, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, K.; Nakamura, K.; Kanai, M.; Nagayama, Y.; Takahashi, S.; Saito, N.; Nagata, M. Determination of urinary and serum pentosidine and its application to elder patients. Biol. Pharm. Bull. 1998, 21, 1005–1008. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.V.; Garnero, P.; Hillier, T.A.; Sellmeyer, D.E.; Strotmeyer, E.S.; Feingold, K.R.; Resnick, H.E.; Tylavsky, F.A.; Black, D.M.; Cummings, S.R.; et al. Pentosidine and increased fracture risk in older adults with type 2 diabetes. J. Clin. Endocrinol. Metab. 2009, 94, 2380–2386. [Google Scholar] [CrossRef]
- Kato, S.; Itoh, K.; Ochiai, M.; Iwai, A.; Park, Y.; Hata, S.; Takeuchi, K.; Ito, M.; Imaki, J.; Miura, S.; et al. Increased pentosidine, an advanced glycation end-product, in urine and tissue reflects disease activity in inflammatory bowel diseases. J. Gastroenterol. Hepatol. 2008, 23 (Suppl. S2), S140–S145. [Google Scholar] [CrossRef]
- Kumar, R.; Tyagi, N.; Nagpal, A.; Kaushik, J.K.; Mohanty, A.K.; Kumar, S. Peptidome profiling of Bubalus bubalis urine and assessment of its antimicrobial activity against mastitis-causing pathogens. Antibiotics 2024, 13, 299. [Google Scholar] [CrossRef]
- Yang, T.; Liu, T.; Lei, T.; Li, T.; Liu, N.; Zhang, M. Unveiling the potential of SLURP1 protein as a biomarker for prostate cancer screening. Front. Oncol. 2024, 14, 1365615. [Google Scholar] [CrossRef]
- Bellei, E.; Caramaschi, S.; Giannico, G.A.; Monari, E.; Martorana, E.; Reggiani Bonetti, L.R.; Bergamini, S. Research of prostate cancer urinary diagnostic biomarkers by proteomics: The noteworthy influence of inflammation. Diagnostics 2023, 13, 1318. [Google Scholar] [CrossRef]
- Zhang, D.; Li, D.; Wang, X.; Sui, Y.; Ma, F.; Dai, Y.; Wang, M.; Qin, W. Urine proteomic signatures of mild hypothermia treatment in cerebral ischemia-reperfusion injury in rats. Cell. Mol. Neurobiol. 2024, 44, 49. [Google Scholar] [CrossRef]
- Ameen, M.; Zafar, M.; Ahmad, M.; Munir, M.; Abid, I.; Mustafa, A.E.M.A.; Athar, M.; Makhkamov, T.; Mamarakhimov, O.; Yuldashev, A.; et al. Cleaner biofuel production via process parametric optimization of nonedible feedstock in a membrane reactor using a titania-based heterogeneous nanocatalyst: An aid to sustainable energy development. Membranes 2023, 13, 889. [Google Scholar] [CrossRef]
- Steegmüller, T.; Kratky, T.; Gollwitzer, L.; Schwaminger, S.P.; Berensmeier, S. Development of a new affinity gold polymer membrane with immobilized protein A. Membranes 2024, 14, 31. [Google Scholar] [CrossRef] [PubMed]
- Dibdiakova, J.; Matic, J.; Wubshet, S.G.; Uhl, W.; Manamperuma, L.D.; Rusten, B.; Vik, E.A. Membrane separation of chicken byproduct hydrolysate for up-concentration of bioactive peptides. Membranes 2024, 14, 28. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Ueda, T.; Sakasai-Sakai, A.; Takeuchi, M. Generation of glyceraldehyde-derived advanced glycation end-products in pancreatic cancer cells and the potential of tumor promotion. World J. Gastroenterol. 2017, 23, 4910–4919. [Google Scholar] [CrossRef]
- Kato, S.; Sugawa, H.; Tabe, K.; Ito, K.; Nakashima, H.; Nagai, R. Rapid pretreatment for multi-sample analysis of advanced glycation end products and their role in nephropathy. J. Clin. Biochem. Nutr. 2022, 70, 256–261. [Google Scholar] [CrossRef]
- Katsuta, N.; Takahashi, H.; Nagai, M.; Sugawa, H.; Nagai, R. Changes in S-(2-succinyl)cysteine and advanced glycation end-products levels in mouse tissues associated with aging. Amino Acids 2022, 54, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Ohno, R.I.; Ichimaru, K.; Tanaka, S.; Sugawa, H.; Katsuta, N.; Sakake, S.; Tominaga, Y.K.; Ban, I.; Shirakawa, J.I.; Yamaguchi, Y.; et al. Glucoselysine is derived from fructose and accumulates in the eye lens of diabetic rats. J. Biol. Chem. 2019, 294, 17326–17338. [Google Scholar] [CrossRef]
- Senavirathna, L.; Ma, C.; Chen, R.; Pan, S. Proteomic investigation of glyceraldehyde-derived intracellular AGEs and their potential influence on pancreatic ductal cells. Cells 2021, 10, 1005. [Google Scholar] [CrossRef]
- Luque-Garcia, J.L.; Neubert, T.A. On-membrane tryptic digestion of proteins for mass spectrometry analysis. Methods Mol. Biol. 2009, 536, 331–341. [Google Scholar]
- Kurien, B.T.; Scofield, R.H. Purification of tryptic digests on polyvinylidene difluoride membrane. Methods Mol. Biol. 2015, 1314, 273–277. [Google Scholar]
- Bickner, A.N.; Champion, M.M.; Hummon, A.B.; Bruening, M.L. Electroblotting through a tryptic membrane for LC-MS/MS analysis of proteins separated in electrophoretic gels. Analyst 2020, 145, 7724–7735. [Google Scholar] [CrossRef]
- Takeuchi, M.; Sakasai-Sakai, A.; Takata, T.; Takino, J.I.; Koriyama, Y. Effects of toxic AGEs (TAGE) on human health. Cells 2022, 11, 2178. [Google Scholar] [CrossRef] [PubMed]
- Hassel, B.; Sørnes, K.; Elsais, A.; Cordero, P.R.; Frøland, A.S.; Rise, F. Glyceraldehyde metabolism in mouse brain and the entry of blood-borne glyceraldehyde into the brain. J. Neurochem. 2024. Online print ahead. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, B.; Hedin, U.; Caidahl, K. Glycolaldehyde and maleyl conjugated human serum albumin as potential macrophage-targeting carriers for molecular imaging purposes. Contrast Media Mol. Imaging 2015, 10, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Zilmer, K.; Leping, V.; Zilmer, M. Serum methylglyoxal level and its association with oxidative stress and disease severity in patients with psoriasis. Arch. Dermatol. Res. 2013, 305, 489–494. [Google Scholar] [CrossRef]
- Dhananjayan, K.; Irrgang, F.; Raju, R.; Harman, D.G.; Moran, C.; Srikanth, V.; Münch, G. Determination of glyoxal and methylglyoxal in serum by UHPLC coupled with fluorescence detection. Anal. Biochem. 2019, 573, 51–66. [Google Scholar] [CrossRef]
- Sarmah, S.; Goswami, A.; Kumar Belwal, V.; Singha Roy, A. Mitigation of ribose and glyoxal induced glycation, AGEs formation and aggregation of human serum albumin by citrus fruit phytochemicals naringin and naringenin: An insight into their mechanism of action. Food Res. Int. 2022, 157, 111358. [Google Scholar] [CrossRef]
- Mirza, M.A.; Kandhro, A.J.; Memon, S.Q.; Khuhawar, M.Y.; Arain, R. Determination of glyoxal and methylglyoxal in the serum of diabetic patients by MEKC using stilbenediamine as derivatizing reagent. Electrophoresis 2007, 28, 3940–3947. [Google Scholar] [CrossRef]
- Shallan, A.I.; Abdel-Hakim, A.; Hammad, M.A.; Abou El-Alamin, M.M. Highly sensitive spectrofluorimetric method for the determination of the genotoxic methylglyoxal in glycerol-containing pharmaceuticals and dietary supplements. Luminescence 2023, 38, 39–46. [Google Scholar] [CrossRef]
- Melvin, M.S.; Avery, K.C.; Ballentine, R.M.; Flora, J.W.; Gardner, W.; Karles, G.D.; Pithawalla, Y.B.; Smith, D.C.; Ehman, K.D.; Wagner, K.A. Formation of diacetyl and other α-dicarbonyl compounds during the generation of E-vapor product aerosols. ACS Omega 2020, 5, 17565–17575. [Google Scholar] [CrossRef]
- Sugiura, K.; Koike, S.; Suzuki, T.; Ogasawara, Y. Oxidative formation of methylglyoxal in glycerol preparations during storage. Biol. Pharm. Bull. 2020, 43, 879–883. [Google Scholar] [CrossRef]
- Schoppel, K.; Trachtmann, N.; Korzin, E.J.; Tzanavari, A.; Sprenger, G.A.; Weuster-Botz, D. Metabolic control analysis enables rational improvement of E. coli L-tryptophan producers but methylglyoxal formation limits glycerol-based production. Microb. Cell Fact. 2022, 21, 201. [Google Scholar] [CrossRef]
- Lapolla, A.; Fedele, D.; Seraglia, R.; Traldi, P. The role of mass spectrometry in the study of non-enzymatic protein glycation in diabetes: An update. Mass Spectrom. Rev. 2006, 25, 775–797. [Google Scholar] [CrossRef]
- Niwa, T. Mass spectrometry for the study of protein glycation in disease. Mass Spectrom. Rev. 2006, 25, 713–723. [Google Scholar] [CrossRef]
- Rhein, S.; Inderhees, J.; Herrmann, O.; Othman, A.; Begemann, K.; Fleming, T.; Nawroth, P.P.; Klika, K.D.; Isa, R.; König, I.R.; et al. Glyoxal in hyperglycaemic ischemic stroke—A cohort study. Cardiovasc. Diabetol. 2023, 22, 173. [Google Scholar] [CrossRef]
- Chen, G.C.; Chai, J.C.; Xing, J.; Moon, J.Y.; Shan, Z.; Yu, B.; Mossavar-Rahman, Y.; Sotres-Alvarez, D.; Li, J.; Mattei, J.; et al. Healthful eating patterns, serum metabolite profile and risk of diabetes in a population-based prospective study of US Hispanics/Latinos. Diabetologia 2022, 65, 1133–1144. [Google Scholar] [CrossRef]
- Hu, C.; Wang, J.; Qi, F.; Liu, Y.; Zhao, F.; Wang, J.; Sun, B. Untargeted metabolite profiling of serum in rats exposed to pyrraline. Food Sci. Biotechnol. 2023, 32, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Usui, T.; Hayase, F. Isolation and identification of the 3-hydroxy-5-hydroxymethyl-pyridinium compound as a novel advanced glycation end product on glyceraldehyde-related Maillard reaction. Biosci. Biotechnol. Biochem. 2003, 67, 930–932. [Google Scholar] [CrossRef] [PubMed]
- Perkins, B.A.; Rabbani, N.; Weston, A.; Ficociello, L.H.; Adaikalakoteswari, A.; Niewczas, M.; Warram, J.; Krolewski, A.S.; Thornalley, P. Serum levels of advanced glycation endproducts and other markers of protein damage in early diabetic nephropathy in type 1 diabetes. PLoS ONE 2012, 7, e35655. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, S.; Mera, K.; Ichikawa, H.; Shimasaki, S.; Nagai, M.; Taga, Y.; Iijima, K.; Hattori, S.; Fujiwara, Y.; Shirakawa, J.I.; et al. Nω-(carboxymethyl)arginine is one of the dominant advanced glycation end products in glycated collagens and mouse tissues. Oxid. Med. Cell. Longev. 2019, 2019, 9073451. [Google Scholar] [CrossRef]
- Ma, Y.; Zhou, Q.; Zhao, P.; Lv, X.; Gong, C.; Gao, J.; Liu, J. Effect of transferrin glycation induced by high glucose on HK-2 cells in vitro. Front. Endocrinol. 2022, 13, 1009507. [Google Scholar] [CrossRef]
- Van der Lugt, T.; Venema, K.; van Leeuwen, S.; Vrolijk, M.F.; Opperhuizen, A.; Bast, A. Gastrointestinal digestion of dietary advanced glycation endproducts using an in vitro model of the gastrointestinal tract (TIM-1). Food Funct. 2020, 11, 6297–6307. [Google Scholar] [CrossRef] [PubMed]
- Nomi, Y.; Kudo, H.; Miyamoto, K.; Okura, T.; Yamamoto, K.; Shimohiro, H.; Kitao, S.; Ito, Y.; Egawa, S.; Kawahara, K.; et al. Free advanced glycation end product distribution in blood components and the effect of genetic polymorphisms. Biochimie 2020, 179, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Adaikalakoteswari, A.; Larkin, J.R.; Panagiotopoulos, S.; MacIsaac, R.J.; Yue, D.K.; Fulcher, G.R.; Roberts, M.A.; Thomas, M.; Ekinci, E.; et al. Analysis of serum advanced glycation endproducts reveals methylglyoxal-derived advanced glycation MG-H1 free adduct is a risk marker in non-diabetic and diabetic chronic kidney disease. Int. J. Mol. Sci. 2022, 24, 152. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, Y.; Sugawa, H.; Hirabayashi, K.; Ikeda, T.; Hoshi, Y.; Nagai, R. Drosera tokaiensis extract containing multiple phenolic compounds inhibits the formation of advanced glycation end-products. Arch. Biochem. Biophys. 2020, 693, 108586. [Google Scholar] [CrossRef]
- Baskal, S.; Tsikas, D. Free L-lysine and its methyl ester react with glyoxal and methylglyoxal in phosphate buffer (100 mM, pH 7.4) to form Nε-carboxymethyl-lysine, Nε-Carboxyethyl-Lysine and Nε-Hydroxymethyl-Lysine. Int. J. Mol. Sci. 2022, 23, 3446. [Google Scholar] [CrossRef]
- Ban, I.; Sugawa, H.; Nagai, R. Protein modification with ribose generates Nδ-(5-hydro-5-methyl-4-imidazolone-2-yl)-ornithine. Int. J. Mol. Sci. 2022, 23, 1224. [Google Scholar] [CrossRef]
- Monnier, V.M.; Mustata, G.T.; Biemel, K.L.; Reihl, O.; Lederer, M.O.; Zhenyu, D.; Sell, D.R. Cross-linking of the extracellular matrix by the Maillard reaction in aging and diabetes: An update on ‘a puzzle nearing resolution’. Ann. N. Y. Acad. Sci. 2005, 1043, 533–544. [Google Scholar] [CrossRef]
- Biemel, K.M.; Reihl, O.; Conrad, J.; Lederer, M.O. Formation pathways for lysine-arginine cross-links derived from hexoses and pentoses by Maillard processes: Unraveling the structure of a pentosidine precursor. J. Biol. Chem. 2001, 276, 23405–23412. [Google Scholar] [CrossRef]
- Sell, D.R.; Biemel, K.M.; Reihl, O.; Lederer, M.O.; Strauch, C.M.; Monnier, V.M. Glucosepane is a major protein cross-link of the senescent human extracellular matrix. Relationship with diabetes. J. Biol. Chem. 2005, 280, 12310–12315. [Google Scholar] [CrossRef]
- Glomb, M.A.; Pfahler, C. Amides are novel protein modifications formed by physiological sugars. J. Biol. Chem. 2001, 276, 41638–41647. [Google Scholar] [CrossRef]
- Biemel, K.M.; Friedl, D.A.; Lederer, M.O. Identification and quantification of major maillard cross-links in human serum albumin and lens protein. Evidence for glucosepane as the dominant compound. J. Biol. Chem. 2002, 277, 24907–24915. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, T.; Sasamoto, K.; Yamamoto, T. Glyceraldehyde-derived advanced glycation end-products having pyrrolopyridinium-based crosslinks. Biochem. Biophys. Rep. 2021, 26, 100963. [Google Scholar] [CrossRef] [PubMed]
- Tessier, F.J.; Monnier, V.M.; Sayre, L.M.; Kornfield, J.A. Triosidines: Novel Maillard reaction products and cross-links from the reaction of triose sugars with lysine and arginine residues. Biochem. J. 2003, 369, 705–719. [Google Scholar] [CrossRef]
- Takeuchi, M.; Suzuki, H.; Takeda, K.; Sakai-Sakasai, A. Toxic advanced glycation end-products (TAGE) are major structures of cytotoxic AGEs Derived from glyceraldehyde. Med. Hypotheses 2024, 183, 11248. [Google Scholar] [CrossRef]
- Oya-Ito, T.; Naito, Y.; Takagi, T.; Handa, O.; Matsui, H.; Yamada, M.; Shima, K.; Yoshikawa, T. Heat-shock protein 27 (Hsp27) as a target of methylglyoxal in gastrointestinal cancer. Biochim. Biophys. Acta 2011, 1812, 769–781. [Google Scholar] [CrossRef]
- Senavirathna, L.; Pan, S.; Chen, R. Protein advanced glycation end products and their implications in pancreatic cancer. Cancer Prev. Res. 2023, 16, 601–610. [Google Scholar] [CrossRef]
- Takata, T.; Masauji, T.; Motoo, Y. Potential of the novel slot blot method with a PVDF membrane for protein identification and quantification in Kampo medicines. Membranes 2023, 13, 896. [Google Scholar] [CrossRef]
- Quansah, E.; Shaik, T.A.; Çevik, E.; Wang, X.; Höppener, C.; Meyer-Zedler, T.; Deckert, V.; Schmitt, M.; Popp, J.; Krafft, C. Investigating biochemical and structural changes of glycated collagen using multimodal multiphoton imaging, Raman spectroscopy, and atomic force microscopy. Anal. Bioanal. Chem. 2023, 415, 6257–6267. [Google Scholar] [CrossRef]
- Matsui, T.; Joo, H.D.; Lee, J.M.; Ju, S.M.; Tao, W.H.; Higashimoto, Y.; Fukami, K.; Yamagishi, S. Development of a monoclonal antibody-based ELISA system for glyceraldehyde-derived advanced glycation end products. Immunol. Lett. 2015, 167, 141–146. [Google Scholar] [CrossRef]
- Kehm, R.; Rückriemen, J.; Weber, D.; Deubel, S.; Grune, T.; Höhn, A. Endogenous advanced glycation end products in pancreatic islets after short-term carbohydrate intervention in obese, diabetes-prone mice. Nutr. Diabetes 2019, 9, 9. [Google Scholar] [CrossRef]
- Kuang, L.; Jing, Z.; Wang, J.; Ma, L.; Liu, X.; Yang, J. Quantitative determination of ɛ-N-carboxymethyl-L-lysine in human plasma by liquid chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2014, 90, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.S.; Ferreira, G.S.; Silvestre, G.C.R.; Santana, M.F.M.; Nunes, V.S.; Ledesma, L.; Pinto, P.R.; de Assis, S.I.S.; Machado, U.F.; da Silva, E.S.; et al. Plasma advanced glycation end products and soluble receptor for advanced glycation end products as indicators of sterol content in human carotid atherosclerotic plaques. Diab. Vasc. Dis. Res. 2022, 19, 14791641221085269. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Du, H.; Ma, Y.; Wang, T.; Zhu, H.; Zhu, L.; Pan, S.; Min, N.; Wang, X.; Liu, Z. Matrine inhibits advanced glycation end products-induced macrophage M1 polarization by reducing DNMT3a/b-mediated DNA methylation of GPX1 promoter. Eur. J. Pharmacol. 2022, 926, 175039. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Gu, M.J.; Kim, Y.; Lee, J.Y.; Lee, S.; Choi, I.W.; Ha, S.K. Glyoxal-lysine dimer, an advanced glycation end product, Induces oxidative damage and inflammatory response by interacting with RAGE. Antioxidants 2021, 10, 1486. [Google Scholar] [CrossRef]
- Oliveira, A.L.; de Oliveira, M.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal and advanced glycation end products (AGEs): Targets for the prevention and treatment of diabetes-associated bladder dysfunction? Biomedicines 2024, 12, 939. [Google Scholar] [CrossRef]
- Lin, Y.Y.; Huang, S.F.; Liao, K.W.; Ho, C.T.; Hung, W.L. Quantitation of α-dicarbonyls, lysine- and arginine-derived advanced glycation end products, in commercial canned meat and seafood products. J. Agric. Food Chem. 2023, 71, 6727–6737. [Google Scholar] [CrossRef]
- Chen, J.; Radjabzadeh, D.; Medina-Gomez, C.; Voortman, T.; van Meurs, J.B.J.; Ikram, M.A.; Uitterlinden, A.G.; Kraaij, R.; Zillikens, M.C. Advanced glycation end products (AGEs) in diet and skin in relation to stool microbiota: The Rotterdam study. Nutrients 2023, 15, 2567. [Google Scholar] [CrossRef]
- Jaggupilli, A.; Howard, R.; Aluko, R.E.; Chelikani, P. Advanced glycation end-products can activate or block bitter taste receptors. Nutrients 2019, 11, 1317. [Google Scholar] [CrossRef]
- Nimmo, J.T.; Verma, A.; Dodart, J.C.; Wang, C.Y.; Savistchenko, J.; Melki, R.; Carare, R.O.; Nicoll, J.A.R. Novel antibodies detect additional α-synuclein pathology in synucleinopathies: Potential development for immunotherapy. Alzheimers. Res. Ther. 2020, 12, 159. [Google Scholar] [CrossRef]
- Kumar, S.T.; Jagannath, S.; Francois, C.; Vanderstichele, H.; Stoops, E.; Lashuel, H.A. How specific are the conformation-specific α-synuclein antibodies? Characterization and validation of 16 α-synuclein conformation-specific antibodies using well-characterized preparations of α-synuclein monomers, fibrils and oligomers with distinct structures and morphology. Neurobiol. Dis. 2020, 146, 105086. [Google Scholar]
- Barandalla, M.; Haucke, E.; Fischer, B.; Navarrete Santos, A.; Colleoni, S.; Galli, C.; Navarrete Santos, A.; Lazzari, G. Comparative analysis of AGE and RAGE levels in human somatic and embryonic stem cells under H2O2-induced noncytotoxic oxidative stress conditions. Oxid. Med. Cell. Longev. 2017, 2017, 4240136. [Google Scholar] [CrossRef]
- Koriyama, Y.; Furukawa, A.; Muramatsu, M.; Takino, J.; Takeuchi, M. Glyceraldehyde caused Alzheimer’s disease-like alterations in diagnostic marker levels in SH-SY5Y human neuroblastoma cells. Sci. Rep. 2015, 5, 13313. [Google Scholar] [CrossRef]
- Bronowicka-Szydełko, A.; Krzystek-Korpacka, M.; Kuzan, A.; Gostomska-Pampuch, K.; Gacka, M.; Jakobsche-Policht, U.; Adamiec, R.; Gamian, A. Non-standard AGE4 epitopes that predict polyneuropathy independently of obesity can be detected by slot dot-blot immunoassay. Adv. Clin. Exp. Med. 2020, 29, 91–100. [Google Scholar] [CrossRef]
- Takata, T.; Murayama, H.; Masauji, T. Slot blot analysis of intracellular glyceraldehyde-derived advanced glycation end products using a novel lysis buffer and polyvinylidene difluoride membrane. Bio. Protoc. 2024, 14, e5038. [Google Scholar] [CrossRef]
- Silva, M.M.; Campos, T.A.; Cavalcanti, I.M.F.; Oliveira, I.S.; Pérez, C.D.; Silva, R.A.D.; Wanderley, M.S.O.; Santos, N.P.S. Proteomic characterization and biological activities of the mucus produced by the zoanthid Palythoa caribaeorum (Duchassaing & Michelotti, 1860). An. Acad. Bras. Cienc. 2023, 95, e20200325. [Google Scholar]
- Shaba, E.; Landi, C.; Marzocchi, C.; Vantaggiato, L.; Bini, L.; Ricci, C.; Cantara, S. Proteomics reveals how the tardigrade damage suppressor protein teaches transfected human cells to survive UV-C stress. Int. J. Mol. Sci. 2023, 24, 11463. [Google Scholar] [CrossRef] [PubMed]
- Gil-Agusti, M.T.; Campostrini, N.; Zolla, L.; Ciambella, C.; Invernizzi, C.; Righetti, P.G. Two-dimensional mapping as a tool for classification of green coffee bean species. Proteomics 2005, 5, 710–718. [Google Scholar] [CrossRef]
- Sakolvaree, Y.; Maneewatch, S.; Jiemsup, S.; Klaysing, B.; Tongtawe, P.; Srimanote, P.; Saengjaruk, P.; Banyen, S.; Tapchaisri, P.; Chonsa-Nguan, M.; et al. Proteome and immunome of pathogenic Leptospira spp. revealed by 2DE and 2DE-immunoblotting with immune serum. Asian Pac. J. Allergy Immunol. 2007, 25, 53–73. [Google Scholar] [PubMed]
- Takata, T.; Sakasai-Sakai, A.; Ueda, T.; Takeuchi, M. Intracellular toxic advanced glycation end-products in cardiomyocytes may cause cardiovascular disease. Sci. Rep. 2019, 9, 2121. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Sakasai-Sakai, A.; Takeuchi, M. Impact of intracellular toxic advanced glycation end-products (TAGE) on murine myoblast cell death. Diabetol. Metab. Syndr. 2020, 12, 54. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Sakasai-Sakai, A.; Takeuchi, M. Intracellular toxic advanced glycation end-products in 1.4E7 cell line induce death with reduction of microtubule-associated protein 1 light chain 3 and p62. Nutrients 2022, 14, 332. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Sakasai-Sakai, A.; Takeuchi, M. Intracellular toxic advanced glycation end-products may induce cell death and suppress cardiac fibroblasts. Metabolites 2022, 12, 615. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, C.; Sakasai-Sakai, A.; Okimura, R.; Tanaka, H.; Takata, T.; Takeuchi, M.; Matsunaga, T. Accumulation of toxic advanced glycation end-products induces cytotoxicity and inflammation in hepatocyte-like cells differentiated from human induced pluripotent stem cells. Biol. Pharm. Bull. 2021, 44, 1399–1402. [Google Scholar] [CrossRef] [PubMed]
- Takata, T.; Sakasai-Sakai, A.; Takino, J.I.; Takeuchi, M. Evidence for toxic advanced glycation end-products generated in the normal Rat liver. Nutrients 2019, 11, 1612. [Google Scholar] [CrossRef]
- Inoue, S.; Takata, T.; Nakazawa, Y.; Nakamura, Y.; Guo, X.; Yamada, S.; Ishigaki, Y.; Takeuchi, M.; Miyazawa, K. Potential of an interorgan network mediated by toxic advanced glycation end-products in a Rat model. Nutrients 2021, 13, 80. [Google Scholar] [CrossRef]
- Sakasai-Sakai, A.; Takata, T.; Takino, J.I.; Takeuchi, M. Impact of intracellular glyceraldehyde-derived advanced glycation end-products on human hepatocyte cell death. Sci. Rep. 2017, 7, 14282. [Google Scholar] [CrossRef]
- Sakasai-Sakai, A.; Takata, T.; Takeuchi, M. Intracellular toxic advanced glycation end-products promote the production of reactive oxygen species in HepG2 cells. Int. J. Mol. Sci. 2020, 21, 4861. [Google Scholar] [CrossRef]
- Sakasai-Sakai, A.; Takata, T.; Takeuchi, M. The association between accumulation of toxic advanced glycation end-products and cytotoxic effect in MC3T3-E1 cells. Nutrients 2022, 14, 990. [Google Scholar] [CrossRef]
- Sakasai-Sakai, A.; Takata, T.; Suzuki, H.; Maruyama, I.; Motomiya, Y.; Takeuchi, M. Immunological evidence for in vivo production of novel advanced glycation end-products from 1,5-anhydro-D-fructose, a glycogen metabolite. Sci. Rep. 2019, 9, 10194. [Google Scholar] [CrossRef]
- McCarthy, J.; Hopwood, F.; Oxley, D.; Laver, M.; Castagna, A.; Righetti, P.G.; Williams, K.; Herbert, B. Carbamylation of proteins in 2-D electrophoresis—Myth or reality? J. Proteome Res. 2003, 2, 239–242. [Google Scholar] [CrossRef]
- Herbert, B. Advances in protein solubilisation for two-dimensional electrophoresis. Electrophoresis 1999, 20, 660–663. [Google Scholar] [CrossRef]
- Han, M.; Han, Q.; Wu, S.; Xiao, H.; Zhang, L.; Lin, Y.; Meng, F.; Zhao, S. Unveiling the impacts of sodium hypochlorite on the characteristics and fouling behaviors of different commercial polyvinylidene fluoride hollow fiber membranes. Membranes 2022, 12, 965. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Makita, Z.; Bucala, R.; Suzuki, T.; Koike, T.; Kameda, Y. Immunological evidence that non-carboxymethyllysine advanced glycation end-products are produced from short chain sugars and dicarbonyl compounds in vivo. Mol. Med. 2000, 6, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Puente, P.; Mateos, J.; Blanco, F.J.; Ruiz-Romero, C. LC-MALDI-TOF/TOF for shotgun proteomics. Methods Mol. Biol. 2014, 1156, 27–38. [Google Scholar]
- Maus, A.; Mignon, R.; Basile, F. Enhanced protein identification using graphite-modified MALDI plates for offline LC-MALDI-MS/MS bottom-up proteomics. Anal. Biochem. 2018, 545, 31–37. [Google Scholar] [CrossRef]
- Fujimoto, S.; Murakami, Y.; Miyake, H.; Hayase, F.; Watanabe, H. Identification of a novel advanced glycation end product derived from lactaldehyde. Biosci. Biotechnol. Biochem. 2019, 83, 1136–1145. [Google Scholar] [CrossRef]
- Nomi, Y.; Sato, T.; Mori, Y.; Matsumoto, H. Evaluation of fructo-, inulin-, and galacto-oligosaccharides on the Maillard reaction products in model systems with whey protein. J. Agric. Food Chem. 2022, 70, 9154–9165. [Google Scholar] [CrossRef]
- Suzuki, R.; Fujiwara, Y.; Saito, M.; Arakawa, S.; Shirakawa, J.I.; Yamanaka, M.; Komohara, Y.; Marumo, K.; Nagai, R. Intracellular accumulation of advanced glycation end products induces osteoblast apoptosis via endoplasmic reticulum stress. J. Bone Miner. Res. 2020, 35, 1992–2003. [Google Scholar] [CrossRef]
- Requena, J.R.; Ahmed, M.U.; Fountain, C.W.; Degenhardt, T.P.; Reddy, S.; Perez, C.; Lyons, T.J.; Jenkins, A.J.; Baynes, J.W.; Thorpe, S.R. Carboxymethylethanolamine, a biomarker of phospholipid modification during the Maillard reaction in vivo. J. Biol. Chem. 1997, 272, 17473–17479. [Google Scholar] [CrossRef]
- Cai, J.; Hurst, H.E. Identification and quantitation of N-(carboxymethyl)valine adduct in hemoglobin by gas chromatography/mass spectrometry. J. Mass Spectrom. 1999, 34, 537–543. [Google Scholar] [CrossRef]
- Woodland, B.; Necakov, A.; Coorssen, J.R. Optimized proteome reduction for integrative Top-Down proteomics. Proteomes 2023, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Toprak, M.; Yuksel, N.; Akpinar, G.; Kasap, M.; Pirhan, D.; Yilmaz Tugan, B.Y. Comparative proteomic analysis of the aqueous humor from patients with pseudoexfoliation syndrome. J. Curr. Glaucoma Pract. 2023, 17, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; López-Martinez, M.J.; Ortin-Bustillo, A.; Cerón, J.J.; Martinez-Subiela, S.; Muñoz-Prieto, A.; Lamy, E. Changes in the saliva proteome of pigs with diarrhoea caused by Escherichia coli. Proteomes 2023, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Govindaiah, P.M.; Maheswarappa, N.B.; Banerjee, R.; Mishra, B.P.; Manohar, B.B.; Dasoju, S. Traditional halal meat production without stunning versus commercial slaughter with electrical stunning of slow-growing broiler chicken: Impact on meat quality and proteome changes. Poult. Sci. 2023, 102, 103033. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solution A (Pre-Takata’s Lysis Buffer) | Solution B | Solution C (Takata’s Lysis Buffer) | Solution D |
|---|---|---|---|
| 30 mM Tris base | 1 protease inhibitor cocktail tablet/2 mL | 27 mM Tris base | 30 mM Tris base |
| 7 M urea | 6.3 M urea | 7 M urea | |
| 2 M thiourea | 1.8 M thiourea | 2 M thiourea | |
| 4 w/v (%) CHAPS | 3.6 w/v (%) CHAPS | 4 w/v (%) CHAPS | |
| 10 (v/v) % Solution B | 4 v/v (%) Solution B | ||
| (pH 8.5) | (pH 8.5) | (pH 8.5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takata, T.; Inoue, S.; Kunii, K.; Masauji, T.; Miyazawa, K. Slot Blot- and Electrospray Ionization–Mass Spectrometry/Matrix-Assisted Laser Desorption/Ionization–Mass Spectrometry-Based Novel Analysis Methods for the Identification and Quantification of Advanced Glycation End-Products in the Urine. Int. J. Mol. Sci. 2024, 25, 9632. https://doi.org/10.3390/ijms25179632
Takata T, Inoue S, Kunii K, Masauji T, Miyazawa K. Slot Blot- and Electrospray Ionization–Mass Spectrometry/Matrix-Assisted Laser Desorption/Ionization–Mass Spectrometry-Based Novel Analysis Methods for the Identification and Quantification of Advanced Glycation End-Products in the Urine. International Journal of Molecular Sciences. 2024; 25(17):9632. https://doi.org/10.3390/ijms25179632
Chicago/Turabian StyleTakata, Takanobu, Shinya Inoue, Kenshiro Kunii, Togen Masauji, and Katsuhito Miyazawa. 2024. "Slot Blot- and Electrospray Ionization–Mass Spectrometry/Matrix-Assisted Laser Desorption/Ionization–Mass Spectrometry-Based Novel Analysis Methods for the Identification and Quantification of Advanced Glycation End-Products in the Urine" International Journal of Molecular Sciences 25, no. 17: 9632. https://doi.org/10.3390/ijms25179632