
Proinflammatory Microenvironment in Adenocarcinoma Tissue of Colorectal Carcinoma

 , , , , , , , , , , , , , and
, , , , , , , , , , , , , and
Abstract
:1. Introduction
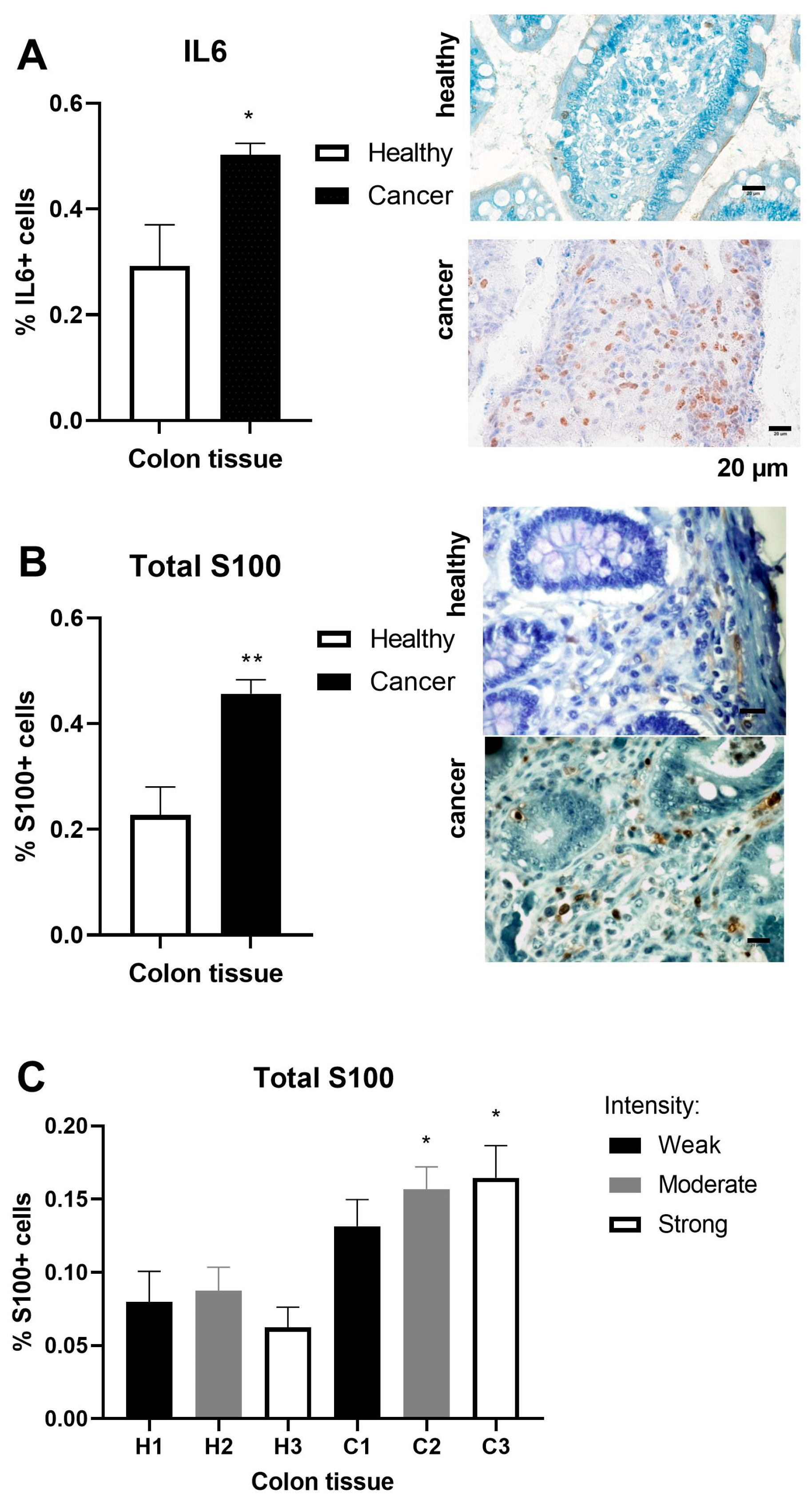
2. Results
2.1. Markers of Intestinal Differentiation in Colorectal Adenocarcinoma
2.2. DNA Susceptibility and Activity in Colorectal Adenocarcinoma
2.3. Inflammation Markers in Colorectal Adenocarcinoma
2.4. Inflammation and Cancer-Related Gene Expression Profile in Colorectal Adenocarcinoma
2.5. Correlation of Clinical and Laboratory Parameters with the Markers of Intestinal Differentiation, Proliferation, and Inflammation in Patients with Colorectal Adenocarcinoma
3. Discussion
4. Materials and Methods
4.1. Clinical and Laboratory Data Collection
4.2. RT-qPCR
4.3. Immunohistochemical Procedure
4.4. Immunoblotting Procedure
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stewart, S.L.; Wike, J.M.; Kato, I.; Lewis, D.R.; Michaud, F. A population-based study of colorectal cancer histology in the United States, 1998–2001. Cancer 2006, 107 (Suppl. S5), 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Przegląd Gastroenterol. 2019, 14, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Dalmartello, M.; La Vecchia, C.; Bertuccio, P.; Boffetta, P.; Levi, F.; Negri, E.; Malvezzi, M. European cancer mortality predictions for the year 2022 with focus on ovarian cancer. Ann. Oncol. 2022, 33, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Testa, U.; Pelosi, E.; Castelli, G. Colorectal Cancer: Genetic Abnormalities, Tumor Progression, Tumor Heterogeneity, Clonal Evolution and Tumor-Initiating Cells. Med. Sci. 2018, 6, 31. [Google Scholar] [CrossRef]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-catenin signalling: Function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef]
- Ghiselli, G.; Agrawal, A. The human D-glucuronyl C5-epimerase gene is transcriptionally activated through the β-catenin–TCF4 pathway. Biochem. J. 2005, 390, 493–499. [Google Scholar] [CrossRef]
- Huang, J.L.; Oshi, M.; Endo, I.; Takabe, K. Clinical relevance of stem cell surface markers CD133, CD24, and CD44 in colorectal cancer. Am. J. Cancer Res. 2021, 11, 5141–5154. [Google Scholar]
- Nagel, J.M.; Lahm, H.; Ofner, A.; Göke, B.; Kolligs, F.T. γ-Catenin acts as a tumor suppressor through context-dependent mechanisms in colorectal cancer. Int. J. Color. Dis. 2017, 32, 1243–1251. [Google Scholar] [CrossRef]
- Yang, K.; Popova, N.V.; Yang, W.C.; Lozonschi, I.; Tadesse, S.; Kent, S.; Bancroft, L.; Matise, I.; Cormier, R.T.; Scherer, S.J.; et al. Interaction of Muc2 and Apc on Wnt Signaling and in Intestinal Tumorigenesis: Potential Role of Chronic Inflammation. Cancer Res. 2008, 68, 7313–7322. [Google Scholar] [CrossRef]
- Velcich, A.; Yang, W.; Heyer, J.; Fragale, A.; Nicholas, C.; Viani, S.; Kucherlapati, R.; Lipkin, M.; Yang, K.; Augenlicht, L. Colorectal Cancer in Mice Genetically Deficient in the Mucin Muc2. Science 2002, 295, 1726–1729. [Google Scholar] [CrossRef]
- Multhoff, G.; Molls, M.; Radons, J. Chronic Inflammation in Cancer Development. Front. Immunol. 2012, 2, 98. [Google Scholar] [CrossRef] [PubMed]
- Il’yasova, D.; Colbert, L.H.; Harris, T.B.; Newman, A.B.; Bauer, D.C.; Satterfield, S.; Kritchevsky, S.B. Circulating Levels of Inflammatory Markers and Cancer Risk in the Health Aging and Body Composition Cohort. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2413–2418. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Lee, J.K.; Choi, J.W.; Lee, C.S.; Sim, J.H.; Cho, C.H.; Lee, K.H.; Cho, I.H.; Chung, M.H.; Kim, H.R.; et al. Interleukin-6 induces S100A9 expression in colonic epithelial cells through STAT3 activation in experimental ulcerative colitis. PLoS ONE 2012, 7, e38801. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Walia, B.; Evans, J.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. IL-6 induces NF-κB activation in the intestinal epithelia. J. Immunol. 2003, 171, 3194–3201. [Google Scholar] [CrossRef]
- Kwon, C.H.; Moon, H.J.; Park, H.J.; Choi, J.H. S100A8 and S100A9 promote invasion and migration through p38 mitogen-activated protein kinase-dependent NF-κB activation in gastric cancer cells. Mol. Cells 2013, 35, 226–234. [Google Scholar] [CrossRef]
- Turovskaya, O.; Foell, D.; Sinha, V.; Vogl, T.; Newlin, R.; Nayak, J.; Nguyen, M.; Olsson, A.; Nawroth, P.; Bierhaus, A.; et al. RAGE, carboxylated glycans, and S100A8/A9 play essential roles in colitis-associated carcinogenesis. Carcinogenesis 2008, 29, 2035–2043. [Google Scholar] [CrossRef]
- Zhong, X.; Xie, F.; Chen, L.; Liu, Z.; Wang, Q. S100A8 and S100A9 promote endothelial cell activation through the RAGE-mediated mammalian target of rapamycin complex 2 pathway. Mol. Med. Rep. 2020, 22, 5293–5303. [Google Scholar] [CrossRef]
- Yu, W.; Tu, Y.; Long, Z.; Liu, J.; Kong, D.; Peng, J.; Wu, H.; Zheng, G.; Zhao, J.; Chen, Y.; et al. Reactive Oxygen Species Bridge the Gap between Chronic Inflammation and Tumor Development. Oxidative Med. Cell. Longev. 2022, 2022, 2606928. [Google Scholar] [CrossRef]
- Moldogazieva, N.T.; Lutsenko, S.V.; Terentiev, A.A. Reactive Oxygen and Nitrogen Species–Induced Protein Modifications: Implication in Carcinogenesis and Anticancer Therapy. Cancer Res. 2018, 78, 6040–6047. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef]
- Johansson, M.E.; Larsson, J.M.H.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host–microbial interactions. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4659–4665. [Google Scholar] [CrossRef] [PubMed]
- Soliman, N.A.; Yussif, S.M. Ki-67 as a prognostic marker according to breast cancer molecular subtype. Cancer Biol. Med. 2016, 13, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Feyerabend, T.B.; Li, J.-P.; Lindahl, U.; Rodewald, H.-R. Heparan sulfate C5-epimerase is essential for heparin biosynthesis in mast cells. Nat. Chem. Biol. 2006, 2, 195–196. [Google Scholar] [CrossRef] [PubMed]
- Prudnikova, T.Y.; Mostovich, L.A.; Domanitskaya, N.V.; Pavlova, T.V.; Kashuba, V.I.; Zabarovsky, E.R.; Grigorieva, E.V. Antiproliferative effect of D-glucuronyl C5-epimerase in human breast cancer cells. Cancer Cell Int. 2010, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. mTOR signaling pathway and mTOR inhibitors in cancer: Progress and challenges. Cell Biosci. 2020, 10, 31. [Google Scholar] [CrossRef]
- Fleming, M.; Ravula, S.; Tatishchev, S.F.; Wang, H.L. Colorectal carcinoma: Pathologic aspects. J. Gastrointest. Oncol. 2012, 3, 153–173. [Google Scholar] [CrossRef]
- Schmitt, M.; Greten, F.R. The inflammatory pathogenesis of colorectal cancer. Nat. Rev. Immunol. 2021, 21, 653–667. [Google Scholar] [CrossRef]
- Dulai, P.S.; Sandborn, W.J.; Gupta, S. Colorectal Cancer and Dysplasia in Inflammatory Bowel Disease: A Review of Disease Epidemiology, Pathophysiology, and Management. Cancer Prev. Res. 2016, 9, 887–894. [Google Scholar] [CrossRef]
- Olsen, J.; Kirkeby, L.T.; Olsen, J.; Eiholm, S.; Jess, T.; Gögenur, I.; Troelsen, J.T. High interleukin-6 mRNA expression is a predictor of relapse in colon cancer. Anticancer Res. 2015, 35, 2235–2240. [Google Scholar]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef]
- Goodla, L.; Xue, X. The Role of Inflammatory Mediators in Colorectal Cancer Hepatic Metastasis. Cells 2022, 11, 2313. [Google Scholar] [CrossRef] [PubMed]
- Kiraly, O.; Gong, G.; Olipitz, W.; Muthupalani, S.; Engelward, B.P. Inflammation-Induced Cell Proliferation Potentiates DNA Damage-Induced Mutations In Vivo. PLoS Genet. 2015, 11, e1004901. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Aikemu, B.; Shao, Y.; Zhang, S.; Yang, G.; Hong, H.; Huang, L.; Jia, H.; Yang, X.; Zheng, M.; et al. Genomic signature of MTOR could be an immunogenicity marker in human colorectal cancer. BMC Cancer 2022, 22, 818. [Google Scholar] [CrossRef] [PubMed]
- Dorf, J.; Zaręba, K.; Matowicka-Karna, J.; Pryczynicz, A.; Guzińska-Ustymowicz, K.; Zalewska, A.; Maciejczyk, M. May the nitrosative and carbonyl stress promote inflammation in patients with colorectal cancer? J. Inflamm. Res. 2022, 15, 4585–4600. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, M.; Williams, R.; Wang, L.; Vogl, T.; Srikrishna, G. S100A8/A9 activate key genes and pathways in colon tumor progression. Mol. Cancer Res. 2011, 9, 133–148. [Google Scholar] [CrossRef]
- Nayak, J.; Mohanty, P.; Lenka, A.; Sahoo, N.; Agrawala, S.; Panigrahi, S.K. Histopathological and Immunohistochemical Evaluation of CDX2 and Ki67 in Colorectal Lesions with their Expression Pattern in Different Histologic Variants, Grade, and Stage of Colorectal Carcinomas. J. Microsc. Ultrastruct. 2021, 9, 183–189. [Google Scholar] [CrossRef]
- Kasprzak, A.; Siodła, E.; Andrzejewska, M.; Szmeja, J.; Seraszek-Jaros, A.; Cofta, S.; Szaflarski, W. Differential expression of mucin 1 and mucin 2 in colorectal cancer. World J. Gastroenterol. 2018, 24, 4164–4177. [Google Scholar] [CrossRef]
- Kesari, M.V.; Gaopande, V.L.; Joshi, A.R.; Babanagare, S.V.; Gogate, B.P.; Khadilkar, A.V. Immunohistochemical study of MUC1, MUC2 and MUC5AC in colorectal carcinoma and review of literature. Indian J. Gastroenterol. 2015, 34, 63–67. [Google Scholar] [CrossRef]
- Betge, J.; Schneider, N.I.; Harbaum, L.; Pollheimer, M.J.; Lindtner, R.A.; Kornprat, P.; Ebert, M.P.; Langner, C. MUC1, MUC2, MUC5AC, and MUC6 in colorectal cancer: Expression profiles and clinical significance. Virchows Arch. 2016, 469, 255–265. [Google Scholar] [CrossRef]
- Pujada, A.; Walter, L.; Patel, A.; Bui, T.A.; Zhang, Z.; Zhang, Y.; Denning, T.L.; Garg, P. Matrix metalloproteinase MMP9 maintains epithelial barrier function and preserves mucosal lining in colitis associated cancer. Oncotarget 2017, 8, 94650–94665. [Google Scholar] [CrossRef]
- Garg, P.; Ravi, A.; Patel, N.R.; Roman, J.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. Matrix Metalloproteinase-9 Regulates MUC-2 Expression Through Its Effect on Goblet Cell Differentiation. Gastroenterology 2007, 132, 1877–1889. [Google Scholar] [CrossRef] [PubMed]
- Horst, D.; Kriegl, L.; Engel, J.; Kirchner, T.; Jung, A. CD133 expression is an independent prognostic marker for low survival in colorectal cancer. Br. J. Cancer 2008, 99, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Mahara, S.; Sun, C.; Doan, A.; Chua, H.K.; Xu, D.; Bian, J.; Li, Y.; Zhu, D.; Sooraj, D.; et al. Genome-scale CRISPR-Cas9 screen of Wnt/β-catenin signaling identifies therapeutic targets for colorectal cancer. Sci. Adv. 2021, 7, eabf2567. [Google Scholar] [CrossRef] [PubMed]
- Mulyawan, I.M. Low Expression of Mucin 2, High Expression of Mucin 13, and High Expression of Nuclear Factor Kappa-Light-Enhancer of Activated B Cells were Significant Pathways in Colorectal Cancer Development. Asian Pac. J. Cancer Prev. 2023, 24, 1119–1123. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Elements | AV | SD | Normal Values | Biochemical Parameters | AV | SD | Normal Values |
|---|---|---|---|---|---|---|---|
| CEA (ng/mL) | 9.08 | 20.91 | <2.5 | Urea (mmol/L) | 6.98 | 7.97 | 2.8–8.1 |
| CA19-9 (U/mL) | 13.79 | 12.97 | <37 | Glucose (mmol/L) | 6.59 | 2.82 | 4.1–5.9 |
| WBC (109/L) | 7.37 | 2.68 | 4.0–10.0 | Bilirubin direct (μmol/L) | 2.54 | 1.04 | <6.0 |
| RBC (1012/L) | 4.30 | 0.61 | 4.5–6.5 | Bilirubin total (μmol/L) | 11.13 | 5.88 | <21.0 |
| HGB (g/L) | 116.8 | 23.33 | 130–170 | AST (U/L) | 22.87 | 13.20 | <32.0 |
| HCT (%) | 36 | 6 | 39–50 | ALT (U/L) | 25.88 | 28.76 | <33.0 |
| MCV (fl) | 83.74 | 8.28 | 82–99 | ALP (U/L) | 141.5 | 182.4 | 35–105 |
| MCH (pg) | 27.17 | 3.70 | 27–32 | LDH (U/L) | 291.8 | 114.1 | 270–428 |
| MCHC (g/L) | 323.4 | 16.77 | 320–360 | K (mmol/L) | 4.45 | 0.59 | 3.5–5.1 |
| RDW-CV (%) | 15.57 | 3.11 | 12–15 | Na (mmol/L) | 139.6 | 3.45 | 136–145 |
| PLT (109/L) | 298.8 | 85.70 | 150–450 | Cl (mmol/L) | 100.5 | 4.45 | 98–107 |
| PCT % | 0.28 | 0.08 | 0.16–0.35 | Ca (mmol/L) | 2.39 | 0.17 | 2.1–2.6 |
| NEUT % | 66.13 | 9.61 | 44–77 | Mg (mmol/L) | 0.78 | 0.08 | 0.66–1.07 |
| LYM % | 22.34 | 7.89 | 20–46 | Phosphorus (mmol/L) | 1.04 | 0.15 | 0.8–1.5 |
| NEUT (109/L) | 4.96 | 2.30 | 2.0–7.5 | Creatinine (μmol/L) | 116.9 | 134 | 44–80 |
| LYM (109/L) | 1.58 | 0.69 | 1.0–4.0 | Amylase (mmol/L) | 74.57 | 28.31 | 28–100 |
| Age (years) | 67.38 | 9.84 | Protein (g/L) | 66.55 | 10.36 | 64–83 | |
| Body mass (kg) | 75.04 | 15.63 | Albumin (g/L) | 38.88 | 5.99 | 35–52 | |
| CRP (g/L) | 33.77 | 50.46 | <5.0 |
| Clinical and Laboratory Parameters | β-Catenin in Epithelium and Stroma | Ki67 S100 | GLCE MUC2 | pmTOR/mTOR Nitrotyrosine | pNFκB/pNFκB | |
|---|---|---|---|---|---|---|
| Age | r | 0.690 | ||||
| p | <0.0001 | |||||
| CA19-9 (U/mL) | r | 0.837 | ||||
| p | 0.006 | |||||
| Metastasis | r | 0.592 | ||||
| p | 0.004 | |||||
| Cancer stage ** | r * | 0.623 | ||||
| p | 0.017 | |||||
| WBC (109/L) | r | 0.621 | ||||
| p | 0.027 | |||||
| RBC (1012/L) | r | −0.624 | −0.703 | |||
| p | 0.015 | 0.009 | ||||
| HCT (%) | r * | −0.670 | −0.731 | |||
| p | 0.006 | 0.006 | ||||
| HGB (g/L) | r * | −0.669 | −0.661 | |||
| p | 0.006 | 0.016 | ||||
| MCH (pg) | r * | −0.507 | ||||
| p | 0.027 | |||||
| MCHC (g/L) | r * | −0.541 | −0.620 | −0.685 | ||
| p | 0.038 | 0.005 | 0.012 | |||
| RDW-CV (%) | r | −0.786 | 0.541 | 0.613 | ||
| p | 0.028 | 0.017 | 0.0287 | |||
| PCT (%) | r | 0.462 | 0.636 | |||
| p | 0.030 | 0.022 | ||||
| Lymphocyte (%) | r * | −0.734 | ||||
| p | 0.038 | |||||
| Neutrophile (109/L) | r | 0.769 | ||||
| p | 0.003 | |||||
| ALT (U/L) | r | −0.870 | ||||
| p | 0.033 | |||||
| AST (U/L) | r | 0.975 | ||||
| p | 0.033 | |||||
| Na (mmol/L) | r * | 0.975 | ||||
| p | 0.033 | |||||
| Cl (mmol/L) | r * | −0.998 | ||||
| p | 0.039 | |||||
| Creatinine (μmol/L) | r | 0.685 | ||||
| p | 0.035 | |||||
| CRP (g/L) | r | 0.975 | ||||
| p | 0.033 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todorović, S.; Ćeranić, M.S.; Tošković, B.; Diklić, M.; Mitrović Ajtić, O.; Subotički, T.; Vukotić, M.; Dragojević, T.; Živković, E.; Oprić, S.; et al. Proinflammatory Microenvironment in Adenocarcinoma Tissue of Colorectal Carcinoma. Int. J. Mol. Sci. 2024, 25, 10062. https://doi.org/10.3390/ijms251810062
Todorović S, Ćeranić MS, Tošković B, Diklić M, Mitrović Ajtić O, Subotički T, Vukotić M, Dragojević T, Živković E, Oprić S, et al. Proinflammatory Microenvironment in Adenocarcinoma Tissue of Colorectal Carcinoma. International Journal of Molecular Sciences. 2024; 25(18):10062. https://doi.org/10.3390/ijms251810062
Chicago/Turabian StyleTodorović, Slobodan, Miljan S. Ćeranić, Borislav Tošković, Miloš Diklić, Olivera Mitrović Ajtić, Tijana Subotički, Milica Vukotić, Teodora Dragojević, Emilija Živković, Svetlana Oprić, and et al. 2024. "Proinflammatory Microenvironment in Adenocarcinoma Tissue of Colorectal Carcinoma" International Journal of Molecular Sciences 25, no. 18: 10062. https://doi.org/10.3390/ijms251810062