
Specific Substrate Activity of Lotus Root Polyphenol Oxidase: Insights from Gaussian-Accelerated Molecular Dynamics and Markov State Models



Abstract
:1. Introduction
2. Results
2.1. Molecular Docking Results
2.2. Structural Stability and Flexibility between Substrates and PPO
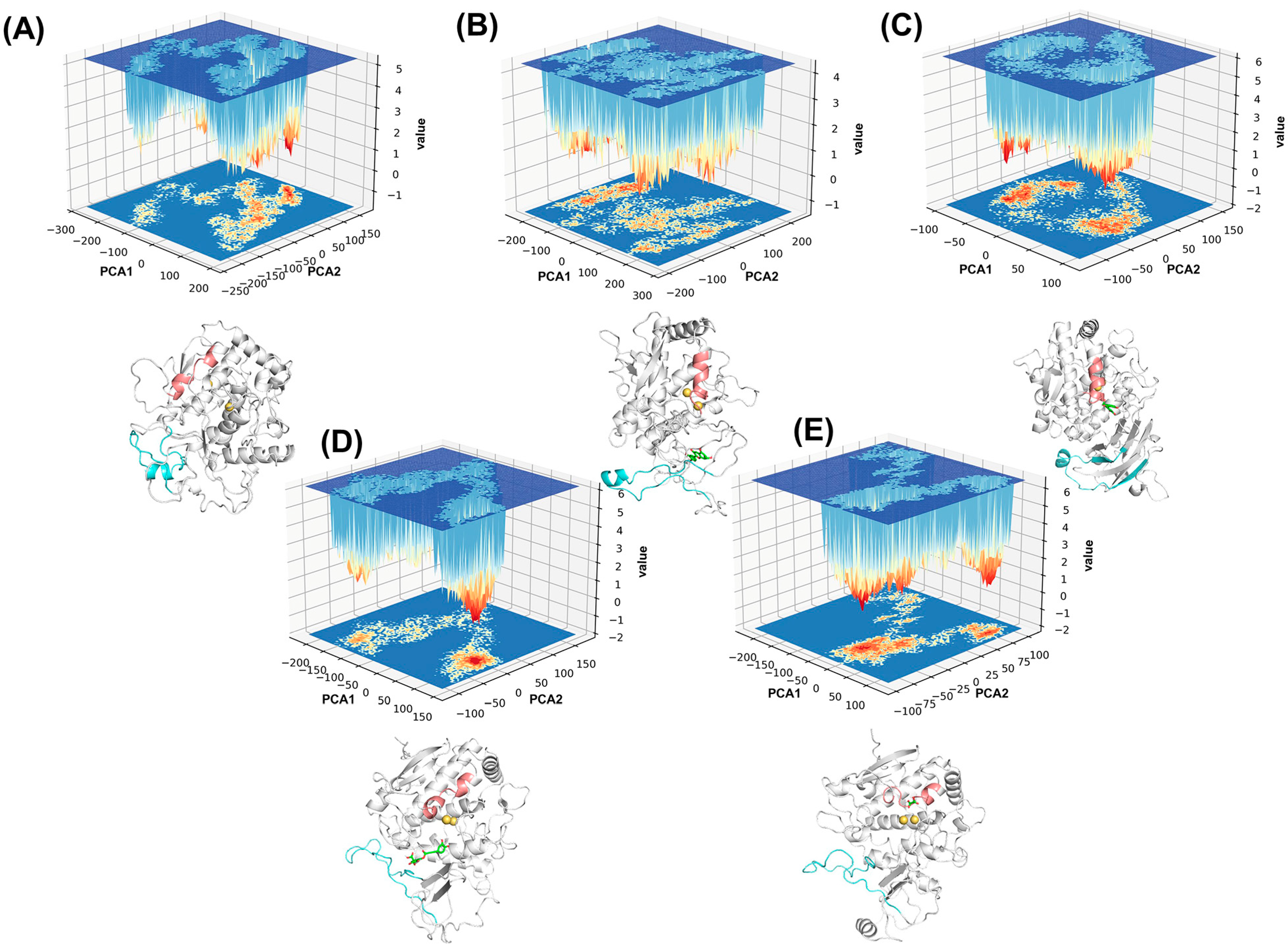
2.3. Analysis of Conformational Changes
2.4. MM-PBSA Analysis
3. Discussion
4. Materials and Methods
4.1. Preparation of Simulated Molecular Systems
4.2. Conventional Molecular Dynamics Simulations
4.3. Gaussian-Accelerated Molecular Dynamics Simulations
4.4. Trajectory Analysis
4.5. MM-PBSA Calculations
4.6. Markov Model Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMBER | Assisted Model Building with Energy Refinement |
| cMD | Conventional Molecular Dynamics |
| DCCM | Dynamic Cross-Correlation Matrix |
| DFT | Density Functional Theory |
| FEL | Free Energy Landscape |
| GaMD | Gaussian-Accelerated Molecular Dynamics |
| HOMO | Highest Occupied Molecular Orbital |
| ITS | Implied Timescales |
| LUMO | Lowest Unoccupied Molecular Orbital |
| MD | Molecular Dynamics |
| MFPT | Mean First Passage Time |
| MM/PBSA | Molecular Mechanics/Poisson–Boltzmann Surface Area |
| MSM | Markov State Model |
| NPT | Isothermal-Isobaric Ensemble (from French: Normale-Isobare) |
| NVT | Constant Number, Volume, and Temperature (from French: Normale-Volume-Température) |
| PCA | Principal Component Analysis |
| PME | Particle Mesh Ewald |
| PPO | Polyphenol Oxidase |
| PBC | Periodic Boundary Conditions |
| RMSD | Root Mean Square Deviation |
| RMSF | Root Mean Square Fluctuation |
| Rg | Radius of Gyration |
| SASA | Solvent Accessible Surface Area |
| TPT | Transition Path Theory |
| ΔEele | Electrostatic Energies |
| ΔEMM | Gas Phase Energy |
| ΔEvdW | Van der Waals Energies |
| ΔGsol | Solvation-Free Energy |
| Amino Acid Single-Letter Abbreviation | |
| F | Phenylalanine |
| G | Glycine |
| L | Leucine |
| V | Valine |
| N | Asparagine |
| Q | Glutamine |
| S | Serine |
References
- Zhang, S. Recent Advances of Polyphenol Oxidases in Plants. Molecules 2023, 28, 2158. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.M.; Kwon, E.B.; Lee, B.; Kim, C.Y. Recent Trends in Controlling the Enzymatic Browning of Fruit and Vegetable Products. Molecules 2020, 25, 2754. [Google Scholar] [CrossRef] [PubMed]
- Macheix, J.J.; Sapis, J.C.; Fleuriet, A. Phenolic compounds and polyphenoloxidase in relation to browning in grapes and wines. Crit. Rev. Food Sci. Nutr. 1991, 30, 441–486. [Google Scholar] [CrossRef]
- Mai, F.; Glomb, M.A. Isolation of phenolic compounds from iceberg lettuce and impact on enzymatic browning. J. Agric. Food Chem. 2013, 61, 2868–2874. [Google Scholar] [CrossRef]
- Friedman, M. Prevention of adverse effects of food browning. Adv. Exp. Med. Biol. 1991, 289, 171–215. [Google Scholar] [CrossRef]
- Ma, Y.; Hong, T.T.; Xu, D.; Wu, F.F.; Xu, X.M. Inhibition of PPO-related browning in fresh noodles: A combination of chemical and heat treatment. Food Chem. 2023, 404, 134549. [Google Scholar] [CrossRef]
- Tilley, A.; McHenry, M.P.; McHenry, J.A.; Solah, V.; Bayliss, K. Enzymatic browning: The role of substrates in polyphenol oxidase mediated browning. Curr. Res. Food Sci. 2023, 7, 100623. [Google Scholar] [CrossRef]
- Sui, X.; Meng, Z.; Dong, T.; Fan, X.; Wang, Q. Enzymatic browning and polyphenol oxidase control strategies. Curr. Opin. Biotechnol. 2023, 81, 102921. [Google Scholar] [CrossRef]
- Tang, M.G.; Zhang, S.; Xiong, L.G.; Zhou, J.H.; Huang, J.A.; Zhao, A.Q.; Liu, Z.H.; Liu, A.L. A comprehensive review of polyphenol oxidase in tea (Camellia sinensis): Physiological characteristics, oxidation manufacturing, and biosynthesis of functional constituents. Compr. Rev. Food Sci. Food Saf. 2023, 22, 2267–2291. [Google Scholar] [CrossRef]
- Cheng, K.; Dong, W.; Long, Y.; Zhao, J.; Hu, R.; Zhang, Y.; Zhu, K. Evaluation of the impact of different drying methods on the phenolic compounds, antioxidant activity, and in vitro digestion of green coffee beans. Food Sci. Nutr. 2019, 7, 1084–1095. [Google Scholar] [CrossRef]
- Toro-Uribe, S.; Godoy-Chivatá, J.; Villamizar-Jaimes, A.R.; Perea-Flores, M.J.; López-Giraldo, L.J. Insight of Polyphenol Oxidase Enzyme Inhibition and Total Polyphenol Recovery from Cocoa Beans. Antioxidants 2020, 9, 458. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Y.; Bi, J.F.; Hu, J.X.; Li, X. Metabolomics comparison of four varieties apple with different browning characters in response to pretreatment during pulp processing. Food Res. Int. 2024, 190, 114600. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Roh, S.H.; Min, S.C. Inactivation of Potato Polyphenol Oxidase Using Microwave Cold Plasma Treatment. J. Food Sci. 2019, 84, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Toledo, L.; Aguirre, C. Enzymatic browning in avocado (Persea americana) revisited: History, advances, and future perspectives. Crit. Rev. Food Sci. Nutr. 2017, 57, 3860–3872. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, F.; Niu, H.; Yuan, L.; Tian, J.; Cai, S.; Bi, X.; Zhou, L. Structural studies and molecular dynamic simulations of polyphenol oxidase treated by high pressure processing. Food Chem. 2022, 372, 131243. [Google Scholar] [CrossRef]
- Zhou, H.; Bie, S.; Li, Z.; Zhou, L. Comparing the Effect of HPP on the Structure and Stability of Soluble and Membrane-Bound Polyphenol Oxidase from ‘Lijiang Snow’ Peach: Multispectroscopic and Molecular Dynamics Simulation. Foods 2023, 12, 1820. [Google Scholar] [CrossRef]
- Hassan, M.; Abbas, Q.; Ashraf, Z.; Moustafa, A.A.; Seo, S.Y. Pharmacoinformatics exploration of polyphenol oxidases leading to novel inhibitors by virtual screening and molecular dynamic simulation study. Comput. Biol. Chem. 2017, 68, 131–142. [Google Scholar] [CrossRef]
- Gacche, R.N.; Gond, D.S.; Dhole, N.A.; Dawane, B.S. Coumarin Schiff-bases: As antioxidant and possibly anti-inflammatory agents. J. Enzym. Inhib. Med. Chem. 2006, 21, 157–161. [Google Scholar] [CrossRef]
- Mayer, A.M. Polyphenol oxidases in plants and fungi: Going places? A review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef]
- Yang, H.; He, S.; Feng, Q.; Liu, Z.; Xia, S.; Zhou, Q.; Wu, Z.; Zhang, Y. Lotus (Nelumbo nucifera): A multidisciplinary review of its cultural, ecological, and nutraceutical significance. Bioresour. Bioprocess. 2024, 11, 18. [Google Scholar] [CrossRef]
- Deng, X.; Huang, J.; Zhang, M.; Wei, X.; Song, H.; Wang, Y.; Xin, J.; Sun, H.; Liu, J.; Yang, D.; et al. Metabolite profiling and screening of callus browning-related genes in lotus (Nelumbo nucifera). Physiol. Plant. 2023, 175, e14027. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Deng, Z.Y.; Dong, H.; Tsao, R.; Liu, X. Substrate specificity of polyphenol oxidase and its selectivity towards polyphenols: Unlocking the browning mechanism of fresh lotus root (Nelumbo nucifera Gaertn.). Food Chem. 2023, 424, 136392. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Arantes, P.R.; Bhattarai, A.; Hsu, R.V.; Pawnikar, S.; Huang, Y.M.; Palermo, G.; Miao, Y. Gaussian accelerated molecular dynamics (GaMD): Principles and applications. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2021, 11, e1521. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, A.; Miao, Y. Gaussian accelerated molecular dynamics for elucidation of drug pathways. Expert. Opin. Drug Discov. 2018, 13, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Collier, T.A.; Piggot, T.J.; Allison, J.R. Molecular Dynamics Simulation of Proteins. Methods Mol. Biol. 2020, 2073, 311–327. [Google Scholar] [CrossRef]
- Lindorff-Larsen, K.; Piana, S.; Dror, R.O.; Shaw, D.E. How fast-folding proteins fold. Science 2011, 334, 517–520. [Google Scholar] [CrossRef]
- da Fonseca, A.M.; Caluaco, B.J.; Madureira, J.M.C.; Cabongo, S.Q.; Gaieta, E.M.; Djata, F.; Colares, R.P.; Neto, M.M.; Fernandes, C.F.C.; Marinho, G.S.; et al. Screening of Potential Inhibitors Targeting the Main Protease Structure of SARS-CoV-2 via Molecular Docking, and Approach with Molecular Dynamics, RMSD, RMSF, H-Bond, SASA and MMGBSA. Mol. Biotechnol. 2024, 66, 1919–1933. [Google Scholar] [CrossRef]
- Tian, M.; Xu, Y.-Y.; Li, Y.-N.; Yu, S.; Wang, Y.-L.; Ma, X.-L.; Zhang, Y.-W. Engineering of Substrate-Binding Domain to Improve Catalytic Activity of Chondroitin B Lyase with Semi-Rational Design. Curr. Issues Mol. Biol. 2024, 46, 9916–9927. [Google Scholar] [CrossRef]
- Kumar, S.U.; Sankar, S.; Kumar, D.T.; Younes, S.; Younes, N.; Siva, R.; Doss, C.G.P.; Zayed, H. Molecular dynamics, residue network analysis, and cross-correlation matrix to characterize the deleterious missense mutations in GALE causing galactosemia III. Cell Biochem. Biophys. 2021, 79, 201–219. [Google Scholar] [CrossRef]
- Blaschek, L.; Pesquet, E. Phenoloxidases in Plants—How Structural Diversity Enables Functional Specificity. Front. Plant Sci. 2021, 12, 754601. [Google Scholar] [CrossRef]
- Khan, M.T.; Ali, S.; Zeb, M.T.; Kaushik, A.C.; Malik, S.I.; Wei, D.Q. Gibbs Free Energy Calculation of Mutation in PncA and RpsA Associated With Pyrazinamide Resistance. Front. Mol. Biosci. 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Golla, V.K.; Prajapati, J.D.; Joshi, M.; Kleinekathöfer, U. Exploration of Free Energy Surfaces Across a Membrane Channel Using Metadynamics and Umbrella Sampling. J. Chem. Theory Comput. 2020, 16, 2751–2765. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res 2023, 51, D523–D531. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Senoo, N.; Chinthapalli, D.K.; Baile, M.G.; Golla, V.K.; Saha, B.; Oluwole, A.O.; Ogunbona, O.B.; Saba, J.A.; Munteanu, T.; Valdez, Y.; et al. Functional diversity among cardiolipin binding sites on the mitochondrial ADP/ATP carrier. EMBO J. 2024, 43, 2979–3008. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Jabłońska, J.; Pravda, L.; Vařeková, R.S.; Thornton, J.M. PDBsum: Structural summaries of PDB entries. Protein Sci. 2018, 27, 129–134. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, J.; Suzek, T.O.; Zhang, J.; Wang, J.; Zhou, Z.; Han, L.; Karapetyan, K.; Dracheva, S.; Shoemaker, B.A.; et al. PubChem’s BioAssay Database. Nucleic Acids Res. 2012, 40, D400–D412. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Case, D.A.; Belfon, K.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; Ghoreishi, D.; et al. AMBER 2022; University of California: San Francisco, CA, USA, 2022. [Google Scholar]
- Harvey, M.J.; Giupponi, G.; Fabritiis, G.D. ACEMD: Accelerating Biomolecular Dynamics in the Microsecond Time Scale. J. Chem. Theory Comput. 2009, 5, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.; Kasavajhala, K.; Belfon, K.A.A.; Raguette, L.; Huang, H.; Migues, A.N.; Bickel, J.; Wang, Y.; Pincay, J.; Wu, Q.; et al. ff19SB: Amino-Acid-Specific Protein Backbone Parameters Trained against Quantum Mechanics Energy Surfaces in Solution. J. Chem. Theory Comput. 2020, 16, 528–552. [Google Scholar] [CrossRef] [PubMed]
- Izadi, S.; Anandakrishnan, R.; Onufriev, A.V. Building Water Models: A Different Approach. J. Phys. Chem. Lett. 2014, 5, 3863–3871. [Google Scholar] [CrossRef] [PubMed]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N.log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E., 3rd. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Dynamics Trajectory Data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef] [PubMed]
- Roe, D.R.; Cheatham, T.E., 3rd. Parallelization of CPPTRAJ enables large scale analysis of molecular dynamics trajectory data. J. Comput. Chem. 2018, 39, 2110–2117. [Google Scholar] [CrossRef]
- Ichiye, T.; Karplus, M. Collective motions in proteins: A covariance analysis of atomic fluctuations in molecular dynamics and normal mode simulations. Proteins 1991, 11, 205–217. [Google Scholar] [CrossRef]
- Srivastava, G.; Tripathi, S.; Kumar, A.; Sharma, A. Molecular investigation of active binding site of isoniazid (INH) and insight into resistance mechanism of S315T-MtKatG in Mycobacterium tuberculosis. Tuberculosis 2017, 105, 18–27. [Google Scholar] [CrossRef]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]
- Homeyer, N.; Gohlke, H. Free Energy Calculations by the Molecular Mechanics Poisson-Boltzmann Surface Area Method. Mol. Inf. 2012, 31, 114–122. [Google Scholar] [CrossRef]
- Wang, C.; Greene, D.; Xiao, L.; Qi, R.; Luo, R. Recent Developments and Applications of the MMPBSA Method. Front. Mol. Biosci. 2017, 4, 87. [Google Scholar] [CrossRef] [PubMed]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W.; et al. Calculating structures and free energies of complex molecules: Combining molecular mechanics and continuum models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, K.; Ma, Y.; Han, W. Probing the Mechanisms of Inhibitors Binding to Presenilin Homologue Using Molecular Dynamics Simulations. Molecules 2023, 28, 2076. [Google Scholar] [CrossRef] [PubMed]
- Scherer, M.K.; Trendelkamp-Schroer, B.; Paul, F.; Pérez-Hernández, G.; Hoffmann, M.; Plattner, N.; Wehmeyer, C.; Prinz, J.H.; Noé, F. PyEMMA 2: A Software Package for Estimation, Validation, and Analysis of Markov Models. J. Chem. Theory Comput. 2015, 11, 5525–5542. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1–S2 | S1–S3 | S1–S4 | S2–S3 | S2–S4 | S3–S4 | |
|---|---|---|---|---|---|---|
| Ligand-free protein | 3.95 | 3.69 | 3.18 | 2.83 | 3.23 | 3.09 |
| Catechin-bound protein | 3.78 | 3.70 | 3.82 | 3.86 | 3.77 | 3.96 |
| Epicatechin-bound protein | 2.31 | 4.28 | 4.26 | 4.13 | 4.17 | 4.25 |
| Chlorogenic acid-bound protein | 2.96 | 2.62 | 4.36 | 1.78 | 4.33 | 4.25 |
| Oxalic acid-bound protein | 2.83 | 3.99 | 4.18 | 3.77 | 4.04 | 2.87 |
| Catechin | Epicatechin | Chlorogenic Acid | Oxalic Acid | |
|---|---|---|---|---|
| ∆EvdW | −19.32 ± 1.17 | −22.50 ± 2.10 | −51.40 ± 1.17 | −0.76 ± 0.44 |
| ∆Eele | −51.65 ± 2.97 | −46.71 ± 5.13 | −22.35 ± 3.00 | −3.57 ± 2.53 |
| ∆Ggas | −70.96 ± 2.83 | −69.21 ± 3.91 | −73.76 ± 3.82 | −4.33 ± 2.94 |
| ∆Gsolv | 59.43 ± 2.07 | 58.46 ± 2.80 | 45.73 ± 3.27 | 4.02 ± 2.62 |
| ∆Gtotal | −11.53 ± 1.00 | −10.75 ± 1.29 | −28.03 ± 1.04 | −0.31 ± 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Zheng, S.; Tang, Y.; Han, W.; Li, W.; Li, T. Specific Substrate Activity of Lotus Root Polyphenol Oxidase: Insights from Gaussian-Accelerated Molecular Dynamics and Markov State Models. Int. J. Mol. Sci. 2024, 25, 10074. https://doi.org/10.3390/ijms251810074
Liu M, Zheng S, Tang Y, Han W, Li W, Li T. Specific Substrate Activity of Lotus Root Polyphenol Oxidase: Insights from Gaussian-Accelerated Molecular Dynamics and Markov State Models. International Journal of Molecular Sciences. 2024; 25(18):10074. https://doi.org/10.3390/ijms251810074
Chicago/Turabian StyleLiu, Minghao, Siyun Zheng, Yijia Tang, Weiwei Han, Wannan Li, and Tao Li. 2024. "Specific Substrate Activity of Lotus Root Polyphenol Oxidase: Insights from Gaussian-Accelerated Molecular Dynamics and Markov State Models" International Journal of Molecular Sciences 25, no. 18: 10074. https://doi.org/10.3390/ijms251810074