
Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury

 and
and
Abstract

1. Introduction
2. Results
2.1. Single Exposure with Nebulized LPS Leads to Prompt Induction of Inflammatory Response in the Lung
2.2. Single Exposure with Nebulized LPS Is Sufficient to Generate a Systemic Bloodstream Translocation
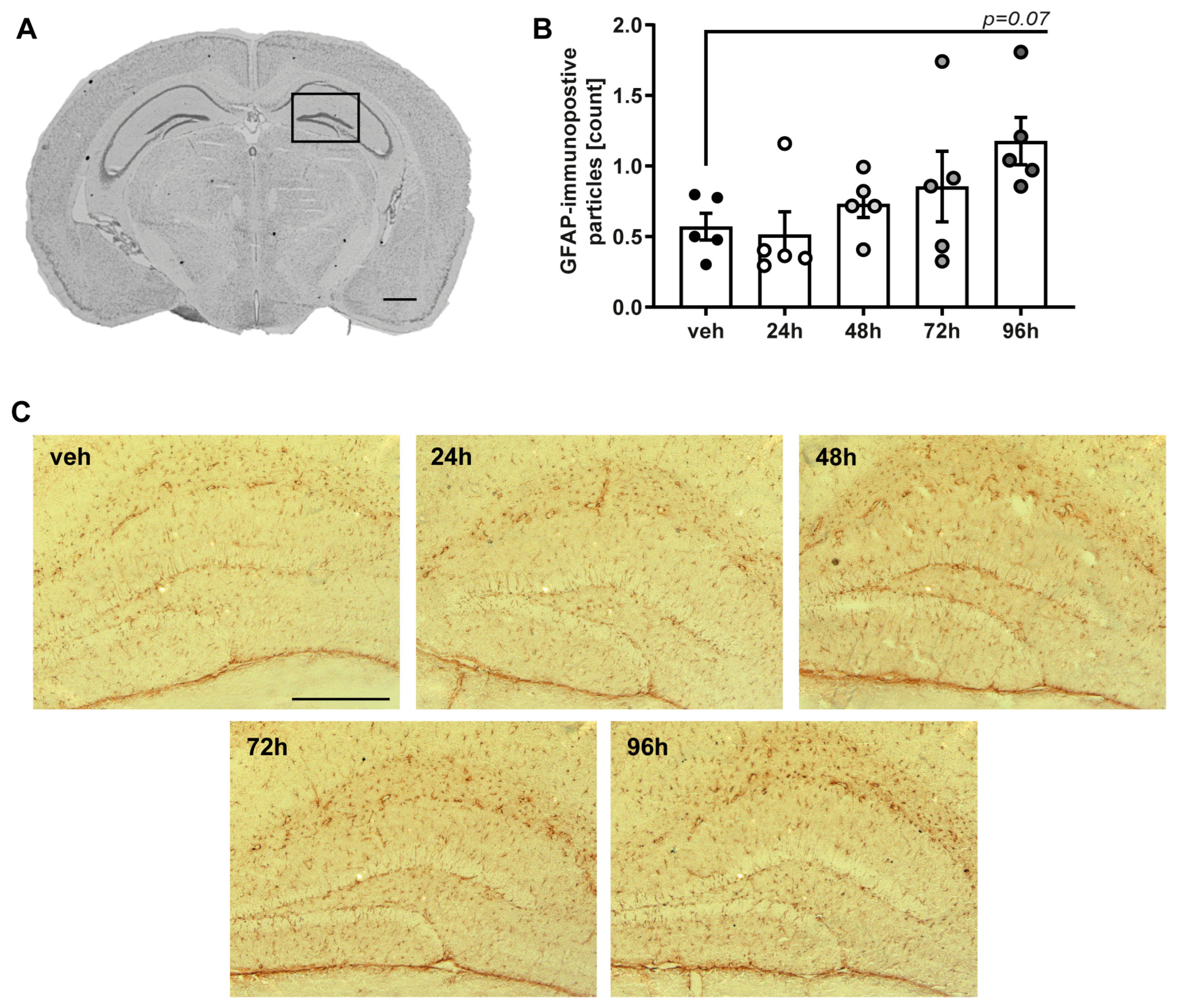
2.3. Single Exposure with Nebulized LPS Leads to Delayed Neuroinflammatory Response in the Cerebral Cortex and Hippocampus
3. Discussion
4. Materials and Methods
4.1. Animals and Study Groups
4.2. Histological Analyses
4.3. Gene Expression Analyses
4.4. Protein Assays
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| ALI | Acute lung injury |
| Arc | Activity-regulated cytoskeletal gene |
| ARDS | Acute respiratory distress syndrome |
| BDNF | Brain-derived neurotrophic factor |
| g | Gram |
| GFAP | Glial fibrillary acidic protein |
| Gadd45b | Growth arrest and DNA damage inducible beta |
| h | Hour |
| i.c.v. | Intracerebroventricular |
| IEGs | Immediate early genes |
| IL | Interleukin |
| Iba-1 | Ionized calcium binding adaptor molecule-1 |
| i.p. | Intraperitoneal |
| iNOS | Inducible nitric oxide synthase |
| i.v. | Intravenous |
| LBP | Lipopolysaccharide-binding protein |
| LPS | Lipopolysaccharide |
| min | Minute |
| MD2 | Myeloid differentiation protein 2 |
| NSS | Neurological Severity Score |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NO | Nitric oxide |
| PBS | Phosphate-buffered saline |
| PFA | Paraformaldehyde |
| RIPA | Radio-Immunoprecipitation assay |
| qPCR | Quantitative real-time polymerase chain reaction |
| ROI | Region of interest |
| RT | Room temperature |
| s | Second |
| SAE | Sepsis-associated encephalopathy |
| SAD | Sepsis-associated delirium |
| TLR4 | Toll-like receptor 4 |
| TNFα | Tumor necrosis factor α |
References
- Ferreira-Coimbra, J.; Sarda, C.; Rello, J. Burden of Community-Acquired Pneumonia and Unmet Clinical Needs. Adv. Ther. 2020, 37, 1302–1318. [Google Scholar] [CrossRef] [PubMed]
- Raoofi, S.; Pashazadeh Kan, F.; Rafiei, S.; Hosseinipalangi, Z.; Noorani Mejareh, Z.; Khani, S.; Abdollahi, B.; Seyghalani Talab, F.; Sanaei, M.; Zarabi, F.; et al. Global prevalence of nosocomial infection: A systematic review and meta-analysis. PLoS ONE 2023, 18, e0274248. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Kang, M.; Lim, J.; Lee, J.; Kang, D.; Kim, M.; Kim, J.; Park, H.; Min, K.H.; Cho, J.; et al. Comprehensive risk assessment for hospital-acquired pneumonia: Sociodemographic, clinical, and hospital environmental factors associated with the incidence of hospital-acquired pneumonia. BMC Pulm. Med. 2022, 22, 21. [Google Scholar] [CrossRef]
- Anderson, R.; Feldman, C. The Global Burden of Community-Acquired Pneumonia in Adults, Encompassing Invasive Pneumococcal Disease and the Prevalence of Its Associated Cardiovascular Events, with a Focus on Pneumolysin and Macrolide Antibiotics in Pathogenesis and Therapy. Int. J. Mol. Sci. 2023, 24, 11038. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990-2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef]
- Speiser, J.L.; Karvellas, C.J.; Shumilak, G.; Sligl, W.I.; Mirzanejad, Y.; Gurka, D.; Kumar, A.; Kumar, A. Predicting in-hospital mortality in pneumonia-associated septic shock patients using a classification and regression tree: A nested cohort study. J. Intensive Care 2018, 6, 66. [Google Scholar] [CrossRef]
- Chaudhry, N.; Duggal, A.K. Sepsis Associated Encephalopathy. Adv. Med. 2014, 2014, 762320. [Google Scholar] [CrossRef]
- Tokuda, R.; Nakamura, K.; Takatani, Y.; Tanaka, C.; Kondo, Y.; Ohbe, H.; Kamijo, H.; Otake, K.; Nakamura, A.; Ishikura, H.; et al. Sepsis-Associated Delirium: A Narrative Review. J. Clin. Med. 2023, 12, 1273. [Google Scholar] [CrossRef] [PubMed]
- Salluh, J.I.; Wang, H.; Schneider, E.B.; Nagaraja, N.; Yenokyan, G.; Damluji, A.; Serafim, R.B.; Stevens, R.D. Outcome of delirium in critically ill patients: Systematic review and meta-analysis. Bmj 2015, 350, h2538. [Google Scholar] [CrossRef]
- Maekawa, T.; Fujii, Y.; Sadamitsu, D.; Yokota, K.; Soejima, Y.; Ishikawa, T.; Miyauchi, Y.; Takeshita, H. Cerebral circulation and metabolism in patients with septic encephalopathy. Am. J. Emerg. Med. 1991, 9, 139–143. [Google Scholar] [CrossRef]
- Imamura, Y.; Wang, H.; Matsumoto, N.; Muroya, T.; Shimazaki, J.; Ogura, H.; Shimazu, T. Interleukin-1β causes long-term potentiation deficiency in a mouse model of septic encephalopathy. Neuroscience 2011, 187, 63–69. [Google Scholar] [CrossRef]
- Gofton, T.E.; Young, G.B. Sepsis-associated encephalopathy. Nat. Rev. Neurol. 2012, 8, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Mazeraud, A.; Righy, C.; Bouchereau, E.; Benghanem, S.; Bozza, F.A.; Sharshar, T. Septic-Associated Encephalopathy: A Comprehensive Review. Neurotherapeutics 2020, 17, 392–403. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Tsikis, S.T.; Fligor, S.C.; Hirsch, T.I.; Pan, A.; Yu, L.J.; Kishikawa, H.; Joiner, M.M.; Mitchell, P.D.; Puder, M. Lipopolysaccharide-induced murine lung injury results in long-term pulmonary changes and downregulation of angiogenic pathways. Sci. Rep. 2022, 12, 10245. [Google Scholar] [CrossRef]
- D’Alessio, F.R. Mouse Models of Acute Lung Injury and ARDS. Methods Mol. Biol. 2018, 1809, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Rissel, R.; Renz, M.; Mohnke, K.; Riedel, J.; Ritter, K.; Ziebart, A.; Ruemmler, R.; Hartmann, E.K.; Kamuf, J. Comparison of two porcine acute lung injury models: A post-hoc analysis. Intensive Care Med. Exp. 2022, 10, 37. [Google Scholar] [CrossRef]
- Aeffner, F.; Bolon, B.; Davis, I.C. Mouse Models of Acute Respiratory Distress Syndrome: A Review of Analytical Approaches, Pathologic Features, and Common Measurements. Toxicol. Pathol. 2015, 43, 1074–1092. [Google Scholar] [CrossRef]
- de Souza Xavier Costa, N.; Ribeiro Júnior, G.; Dos Santos Alemany, A.A.; Belotti, L.; Zati, D.H.; Frota Cavalcante, M.; Matera Veras, M.; Ribeiro, S.; Kallás, E.G.; Nascimento Saldiva, P.H.; et al. Early and late pulmonary effects of nebulized LPS in mice: An acute lung injury model. PLoS ONE 2017, 12, e0185474. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef]
- Wiener-Kronish, J.P.; Albertine, K.H.; Matthay, M.A. Differential responses of the endothelial and epithelial barriers of the lung in sheep to Escherichia coli endotoxin. J. Clin. Investig. 1991, 88, 864–875. [Google Scholar] [CrossRef]
- Skrzypczak-Wiercioch, A.; Sałat, K. Lipopolysaccharide-Induced Model of Neuroinflammation: Mechanisms of Action, Research Application and Future Directions for Its Use. Molecules 2022, 27, 5481. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Bi, W.; Xiao, S.; Lan, X.; Cheng, X.; Zhang, J.; Lu, D.; Wei, W.; Wang, Y.; Li, H.; et al. Neuroinflammation induced by lipopolysaccharide causes cognitive impairment in mice. Sci. Rep. 2019, 9, 5790. [Google Scholar] [CrossRef] [PubMed]
- Murtaj, V.; Belloli, S.; Di Grigoli, G.; Pannese, M.; Ballarini, E.; Rodriguez-Menendez, V.; Marmiroli, P.; Cappelli, A.; Masiello, V.; Monterisi, C.; et al. Age and Sex Influence the Neuro-inflammatory Response to a Peripheral Acute LPS Challenge. Front. Aging Neurosci. 2019, 11, 299. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, Q.; Li, B.; Li, S.; Yang, Z.; Yuan, F.; Liu, Z. A single dose of lipopolysaccharide elicits autofluorescence in the mouse brain. Front. Aging Neurosci. 2023, 15, 1126273. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Zhang, K.; Li, Y.; Tang, Z.; Zheng, R.; Ma, Y.; Chen, Z.; Lei, N.; Xiong, L.; Guo, P.; et al. Lipopolysaccharide-induced depression-like model in mice: Meta-analysis and systematic evaluation. Front. Immunol. 2023, 14, 1181973. [Google Scholar] [CrossRef]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; de Oliveira, A.C.P. Lipopolysaccharide-Induced Neuroinflammation as a Bridge to Understand Neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef]
- Singh, S.; Sahu, K.; Singh, C.; Singh, A. Lipopolysaccharide induced altered signaling pathways in various neurological disorders. Naunyn Schmiedebergs Arch. Pharmacol. 2022, 395, 285–294. [Google Scholar] [CrossRef]
- Eng, L.F.; Ghirnikar, R.S.; Lee, Y.L. Glial fibrillary acidic protein: GFAP-thirty-one years (1969–2000). Neurochem. Res. 2000, 25, 1439–1451. [Google Scholar] [CrossRef]
- Chen, T.; Xu, Y.-P.; Chen, Y.; Sun, S.; Yan, Z.-Z.; Wang, Y.-H. Arc regulates brain damage and neuroinflammation via Sirt1 signaling following subarachnoid hemorrhage. Brain Res. Bull. 2023, 203, 110780. [Google Scholar] [CrossRef]
- Cruz-Mendoza, F.; Jauregui-Huerta, F.; Aguilar-Delgadillo, A.; García-Estrada, J.; Luquin, S. Immediate Early Gene c-fos in the Brain: Focus on Glial Cells. Brain Sci. 2022, 12, 687. [Google Scholar] [CrossRef]
- Salerno, D.M.; Tront, J.S.; Hoffman, B.; Liebermann, D.A. Gadd45a and Gadd45b modulate innate immune functions of granulocytes and macrophages by differential regulation of p38 and JNK signaling. J. Cell. Physiol. 2012, 227, 3613–3620. [Google Scholar] [CrossRef]
- Medzhitov, R. Recognition of microorganisms and activation of the immune response. Nature 2007, 449, 819–826. [Google Scholar] [CrossRef]
- Domscheit, H.; Hegeman, M.A.; Carvalho, N.; Spieth, P.M. Molecular Dynamics of Lipopolysaccharide-Induced Lung Injury in Rodents. Front. Physiol. 2020, 11, 36. [Google Scholar] [CrossRef]
- Menezes, S.L.; Bozza, P.T.; Neto, H.C.; Laranjeira, A.P.; Negri, E.M.; Capelozzi, V.L.; Zin, W.A.; Rocco, P.R. Pulmonary and extrapulmonary acute lung injury: Inflammatory and ultrastructural analyses. J. Appl. Physiol. 2005, 98, 1777–1783. [Google Scholar] [CrossRef]
- Al-Ani, B.; ShamsEldeen, A.M.; Kamar, S.S.; Haidara, M.A.; Al-Hashem, F.; Alshahrani, M.Y.; Al-Hakami, A.M.; Kader, D.H.A.; Maarouf, A. Lipopolysaccharide induces acute lung injury and alveolar haemorrhage in association with the cytokine storm, coagulopathy and AT1R/JAK/STAT augmentation in a rat model that mimics moderate and severe Covid-19 pathology. Clin. Exp. Pharmacol. Physiol. 2022, 49, 483–491. [Google Scholar] [CrossRef]
- Pelgrim, C.E.; van Ark, I.; Leusink-Muis, T.; Brans, M.A.D.; Braber, S.; Garssen, J.; van Helvoort, A.; Kraneveld, A.D.; Folkerts, G. Intratracheal administration of solutions in mice; development and validation of an optimized method with improved efficacy, reproducibility and accuracy. J. Pharmacol. Toxicol. Methods 2022, 114, 107156. [Google Scholar] [CrossRef]
- Raduolovic, K.; Mak’Anyengo, R.; Kaya, B.; Steinert, A.; Niess, J.H. Injections of Lipopolysaccharide into Mice to Mimic Entrance of Microbial-derived Products After Intestinal Barrier Breach. J. Vis. Exp. 2018, 135, 57610. [Google Scholar] [CrossRef]
- Hummel, R.; Lang, M.; Walderbach, S.; Wang, Y.; Tegeder, I.; Gölz, C.; Schäfer, M.K.E. Single intracerebroventricular progranulin injection adversely affects the blood-brain barrier in experimental traumatic brain injury. J. Neurochem. 2021, 158, 342–357. [Google Scholar] [CrossRef]
- Wang, Y.; Wernersbach, I.; Strehle, J.; Li, S.; Appel, D.; Klein, M.; Ritter, K.; Hummel, R.; Tegeder, I.; Schäfer, M.K.E. Early posttraumatic CSF1R inhibition via PLX3397 leads to time- and sex-dependent effects on inflammation and neuronal maintenance after traumatic brain injury in mice. Brain Behav. Immun. 2022, 106, 49–66. [Google Scholar] [CrossRef]
- Ritter, K.; Vetter, D.; Wernersbach, I.; Schwanz, T.; Hummel, R.; Schäfer, M.K.E. Pre-traumatic antibiotic-induced microbial depletion reduces neuroinflammation in acute murine traumatic brain injury. Neuropharmacology 2023, 237, 109648. [Google Scholar] [CrossRef]
- Kang, J.-B.; Park, D.-J.; Shah, M.-A.; Kim, M.-O.; Koh, P.-O. Lipopolysaccharide induces neuroglia activation and NF-κB activation in cerebral cortex of adult mice. Lab. Anim. Res. 2019, 35, 19. [Google Scholar] [CrossRef]
- Che, D.N.; Cho, B.O.; Kim, J.S.; Shin, J.Y.; Kang, H.J.; Jang, S.I. Luteolin and Apigenin Attenuate LPS-Induced Astrocyte Activation and Cytokine Production by Targeting MAPK, STAT3, and NF-κB Signaling Pathways. Inflammation 2020, 43, 1716–1728. [Google Scholar] [CrossRef]
- Tarassishin, L.; Suh, H.S.; Lee, S.C. LPS and IL-1 differentially activate mouse and human astrocytes: Role of CD14. Glia 2014, 62, 999–1013. [Google Scholar] [CrossRef]
- Simi, A.; Edling, Y.; Ingelman-Sundberg, M.; Tindberg, N. Activation of c-fos by lipopolysaccharide in glial cells via p38 mitogen-activated protein kinase-dependent activation of serum or cyclic AMP/calcium response element. J. Neurochem. 2005, 92, 915–924. [Google Scholar] [CrossRef]
- Suh, H.W.; Choi, S.S.; Lee, J.K.; Lee, H.K.; Han, E.J.; Lee, J. Regulation of c-fos and c-jun gene expression by lipopolysaccharide and cytokines in primary cultured astrocytes: Effect of PKA and PKC pathways. Arch. Pharm. Res. 2004, 27, 396–401. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Y.; Guo, T.; Tang, C.; Liu, L.; Gao, D. High Concentration of Glial Cell Line-Derived Neurotrophic Factor Protects Primary Astrocytes from Apoptosis. Dev. Neurosci. 2018, 40, 134–144. [Google Scholar] [CrossRef]
- Yakout, D.W.; Shree, N.; Mabb, A.M. Effect of pharmacological manipulations on Arc function. Curr. Res. Pharmacol. Drug Discov. 2021, 2, 100013. [Google Scholar] [CrossRef]
- Wall, M.J.; Collins, D.R.; Chery, S.L.; Allen, Z.D.; Pastuzyn, E.D.; George, A.J.; Nikolova, V.D.; Moy, S.S.; Philpot, B.D.; Shepherd, J.D.; et al. The Temporal Dynamics of Arc Expression Regulate Cognitive Flexibility. Neuron 2018, 98, 1124–1132. [Google Scholar] [CrossRef]
- Rosi, S. Neuroinflammation and the plasticity-related immediate-early gene Arc. Brain Behav. Immun. 2011, 25 (Suppl. 1), S39–S49. [Google Scholar] [CrossRef]
- Rosi, S.; Ramirez-Amaya, V.; Vazdarjanova, A.; Worley, P.F.; Barnes, C.A.; Wenk, G.L. Neuroinflammation alters the hippocampal pattern of behaviorally induced Arc expression. J. Neurosci. 2005, 25, 723–731. [Google Scholar] [CrossRef]
- Anderson, S.T.; Commins, S.; Moynagh, P.N.; Coogan, A.N. Lipopolysaccharide-induced sepsis induces long-lasting affective changes in the mouse. Brain Behav. Immun. 2015, 43, 98–109. [Google Scholar] [CrossRef]
- Peng, X.; Luo, Z.; He, S.; Zhang, L.; Li, Y. Blood-Brain Barrier Disruption by Lipopolysaccharide and Sepsis-Associated Encephalopathy. Front. Cell. Infect. Microbiol. 2021, 11, 768108. [Google Scholar] [CrossRef]
- Huang, C.; Ye, T.; Chen, B.; Chen, Z.; Ye, Y.; Liu, H. Intranasal administration of lipopolysaccharide reverses chronic stress-induced depression-like behavior in mice by microglial stimulation. Int. Immunopharmacol. 2023, 120, 110347. [Google Scholar] [CrossRef]
- Jiao, Y.; Tong, C.S.W.; Zhao, L.; Zhang, Y.; Nicholls, J.M.; Rainer, T.H. Intraperitoneal versus intranasal administration of lipopolysaccharide in causing sepsis severity in a murine model: A preliminary comparison. Lab. Anim. Res. 2024, 40, 18. [Google Scholar] [CrossRef]
- McCarthy, G.M.; Bridges, C.R.; Blednov, Y.A.; Harris, R.A. CNS cell-type localization and LPS response of TLR signaling pathways. F1000Research 2017, 6, 1144. [Google Scholar] [CrossRef]
- Vargas-Caraveo, A.; Sayd, A.; Robledo-Montaña, J.; Caso, J.R.; Madrigal, J.L.M.; García-Bueno, B.; Leza, J.C. Toll-like receptor 4 agonist and antagonist lipopolysaccharides modify innate immune response in rat brain circumventricular organs. J. Neuroinflamm. 2020, 17, 6. [Google Scholar] [CrossRef]
- Da Ré, C.; Souza, J.M.; Fróes, F.; Taday, J.; dos Santos, J.P.; Rodrigues, L.; Sesterheim, P.; Gonçalves, C.A.; Leite, M.C. Neuroinflammation induced by lipopolysaccharide leads to memory impairment and alterations in hippocampal leptin signaling. Behav. Brain Res. 2020, 379, 112360. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Yuki, K.; Koutsogiannaki, S. Translational Role of Rodent Models to Study Ventilator-Induced Lung Injury. Transl. Perioper. Pain Med. 2021, 8, 404–415. [Google Scholar]
- Ritter, K.; Jung, K.; Dolderer, C.; Appel, D.; Oswald, C.C.; Ritz, U.; Schäfer, M.K.E. Early Reciprocal Effects in a Murine Model of Traumatic Brain Injury and Femoral Fracture. Mediat. Inflamm. 2021, 2021, 8835730. [Google Scholar] [CrossRef]
- Huang, C.; Sakry, D.; Menzel, L.; Dangel, L.; Sebastiani, A.; Krämer, T.; Karram, K.; Engelhard, K.; Trotter, J.; Schäfer, M.K. Lack of NG2 exacerbates neurological outcome and modulates glial responses after traumatic brain injury. Glia 2016, 64, 507–523. [Google Scholar] [CrossRef]
- Pöttker, B.; Stöber, F.; Hummel, R.; Angenstein, F.; Radyushkin, K.; Goldschmidt, J.; Schäfer, M.K.E. Traumatic brain injury causes long-term behavioral changes related to region-specific increases of cerebral blood flow. Brain Struct. Funct. 2017, 222, 4005–4021. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name, (Amplicon Size, bp) | Oligonucleotide Sequences 5′–3′ (fw: Forward, rev: Reverse) | Gene Bank Number |
|---|---|---|
| Arc (192) | fw-CTCAACTTCCGGGGATGCAG rev-CTGGTATGAATCACTGGGGGC | NM_001276684 |
| Fos (165) | fw-CGGGTTTCAACGCCGACTA Rev-TGGCACTAGAGACGGACAGAT | NM_010234 |
| Gadd45b (113) | fw-CCTCCTGGTCACGAACTGTC rev-TGGGTCTCAGCGTTCCTCTA | NM_008655 |
| Gfap (120) | fw-CGGAGACGCATCACCTCTG rev-TGGAGGAGTCATTCGAGACAA | NM_001131020 |
| IL-6 (471) | fw-CATAAAATAGTCCTTCCTACCCCAATTTCC-FL rev-TATGCTTAGGCATAACGCACTAG | NM_031168 |
| Ppia (146) | fw-GCGTCTSCTTCGAGCTGTT rev-RAAGTCACCCTGGCA | NM_008907 |
| TLR4 (134) | fw-GCTTTCACCTCTGCCTTCAC rev-CCAACGGCTCTGAATAAAGTG | NM_021297.3 |
| TNFalpha (212) | fw-TCTCATCAGTTCTATGGCCC rev-GGGAGTAGACAAGGTACAAC | NM_013693 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritter, K.; Rissel, R.; Renz, M.; Ziebart, A.; Schäfer, M.K.E.; Kamuf, J. Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury. Int. J. Mol. Sci. 2024, 25, 10117. https://doi.org/10.3390/ijms251810117
Ritter K, Rissel R, Renz M, Ziebart A, Schäfer MKE, Kamuf J. Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury. International Journal of Molecular Sciences. 2024; 25(18):10117. https://doi.org/10.3390/ijms251810117
Chicago/Turabian StyleRitter, Katharina, René Rissel, Miriam Renz, Alexander Ziebart, Michael K. E. Schäfer, and Jens Kamuf. 2024. "Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury" International Journal of Molecular Sciences 25, no. 18: 10117. https://doi.org/10.3390/ijms251810117
APA StyleRitter, K., Rissel, R., Renz, M., Ziebart, A., Schäfer, M. K. E., & Kamuf, J. (2024). Nebulized Lipopolysaccharide Causes Delayed Cortical Neuroinflammation in a Murine Model of Acute Lung Injury. International Journal of Molecular Sciences, 25(18), 10117. https://doi.org/10.3390/ijms251810117