
Risk Assessment of Fenpropathrin: Cause Hepatotoxicity and Nephrotoxicity in Common Carp (Cyprinus carpio L.)

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Serum Biochemical Parameters
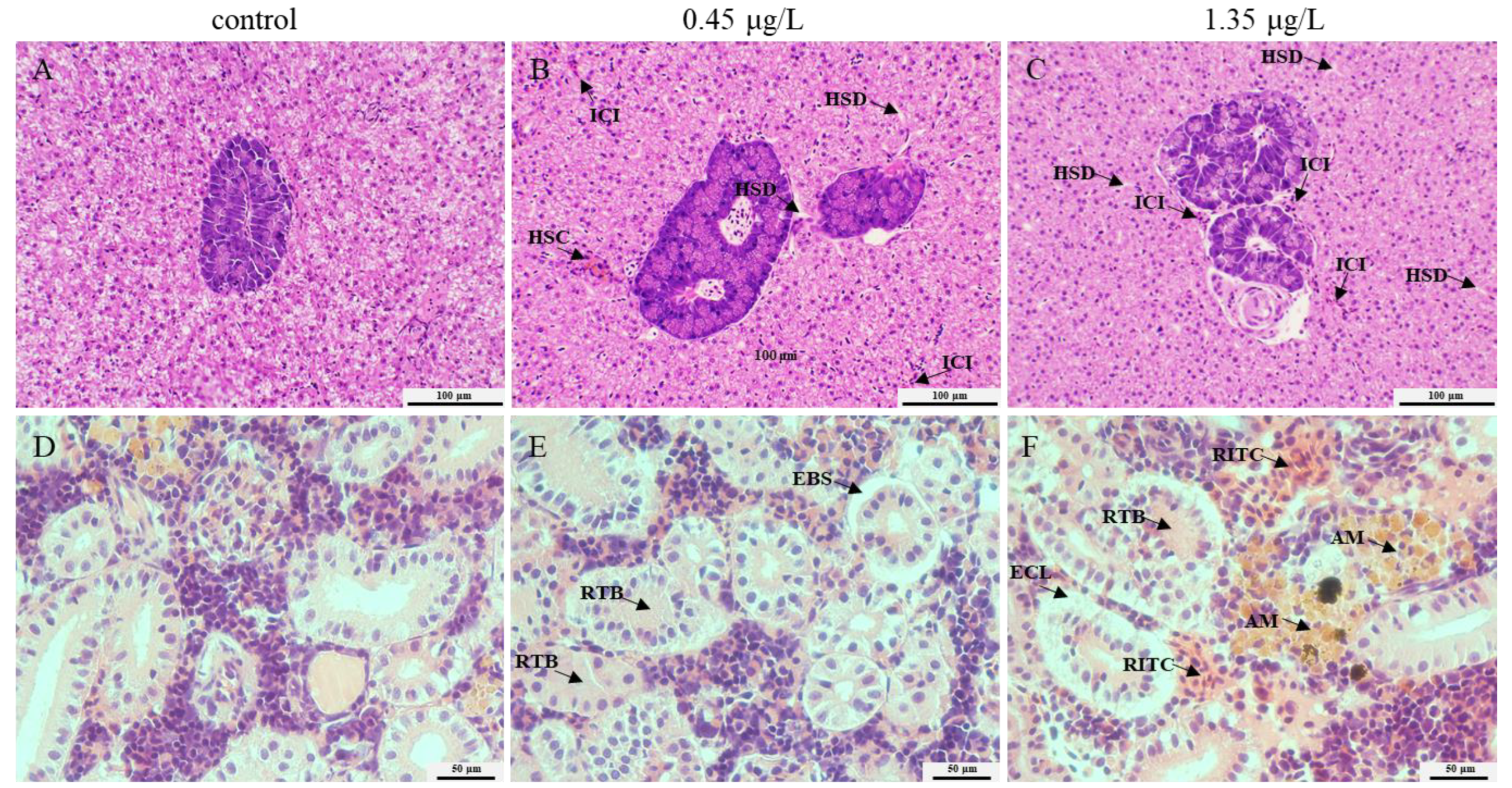
2.2. Histopahology
2.3. Oxidative Stress-Related Parameters in Fish Liver and Kidney
2.4. Inflammation-Related Parameters in Fish Liver and Kidney
2.5. DEGs and qPCR Validation
2.6. Gene Ontology (GO) Analysis of DEGs
2.7. Kyoto Encyclopedia of Genes and Genomes (KEGG) and Protein-Protein Interaction (PPI) Analysis of DEGs
3. Discussion
4. Materials and Methods
4.1. FEN and Common Carp
4.2. Experimental Design and Sample Collection
4.3. Biochemical Parameters Examination
4.4. Transcriptomic Analysis
4.5. Quantitative PCR (qPCR) Analysis
4.6. Histological Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shafer, T.J.; Meyer, D.A.; Crofton, K.M. Developmental neurotoxicity of pyrethroid insecticides: Critical review and future research needs. Environ. Health Perspect. 2005, 113, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Zhang, X.; Huang, J.; Chen, C.; Chen, Z.; Liu, L.; Zhang, G.; Yang, J.; Zhang, Z.; Zhang, Z.; et al. Fenpropathrin, a widely used pesticide, causes dopaminergic degeneration. Mol. Neurobiol. 2016, 53, 995–1008. [Google Scholar] [CrossRef] [PubMed]
- Breckenridge, C.B.; Holden, L.; Sturgess, N.; Weiner, M.; Sheets, L.; Sargent, D.; Soderlund, D.M.; Choi, J.S.; Symington, S.; Clark, J.M.; et al. Evidence for a separate mechanism of toxicity for the Type I and the Type II pyrethroid insecticides. Neurotoxicology 2009, 30, S17–S31. [Google Scholar] [CrossRef] [PubMed]
- Soderlund, D.M.; Clark, J.M.; Sheets, L.P.; Mullin, L.S.; Piccirillo, V.J.; Sargent, D.; Stevens, J.T.; Weiner, M.L. Mechanisms of pyrethroid neurotoxicity: Implications for cumulative risk assessment. Toxicology 2002, 171, 3–59. [Google Scholar] [CrossRef] [PubMed]
- Kanawi, E.; Budd, R.; Tjeerdema, R.S. Environmental fate and ecotoxicology of fenpropathrin. Rev. Environ. Contam. Toxicol. 2013, 225, 77–93. [Google Scholar]
- Tong, Z.; Duan, J.; Wu, Y.; Liu, Q.; He, Q.; Shi, Y.; Yu, L.; Cao, H. A survey of multiple pesticide residues in pollen and beebread collected in China. Sci. Total Environ. 2018, 640, 1578–1586. [Google Scholar] [CrossRef]
- Deng, F.; Sun, J.; Dou, R.; Yu, X.; Wei, Z.; Yang, C.; Zeng, X.; Zhu, L. Contamination of pyrethroids in agricultural soils from the Yangtze River Delta, China. Sci. Total Environ. 2020, 731, 139181. [Google Scholar] [CrossRef]
- Fang, S.H.; Chen, P.; Bian, J.N.; Zhong, W.Y.; Zhu, L.Y. Levels and toxicity assessment of pyrethroids in the surface sediments of Taihu Lake and Liaohe River. Acta Scient. Circum. 2012, 32, 2600–2606. (In Chinese) [Google Scholar]
- Zhao, L.; Lai, Z.; Li, X.; Wang, C.; Shuai, F.; Zeng, Y.; Yang, W. Contamination and toxicity evaluation of pyrethroids in sediments of the Pearl River Estuary. Ecol. Environ. Sci. 2013, 22, 1408–1413. [Google Scholar]
- Ni, Y.N. Research on Pesticide Pollution Control Technology at the Source of Drinking Water Sources in Guiyang. Master’s Thesis, Guizhou University, Guiyang, China, 2018. (In Chinese). [Google Scholar]
- Zhao, Y.Q.; Li, L.N.; Li, J.H. Study of acute and joint toxicity of common pyrethroid and organophosphate pesticides to fish. Environ. Pollut. 2008, 11, 53–57. (In Chinese) [Google Scholar]
- Kouzayha, A.; Al Ashi, A.; Al Akoum, R.; Al Iskandarani, M.; Budzinski, H.; Jaber, F. Occurrence of pesticide residues in Lebanon’s water resources. Bull. Environ. Contam. Toxicol. 2013, 91, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Al-Makkawy, H.K.; Madbouly, M.D. Persistence and accumulation of some organic insecticides in Nile water and fish. Resour. Conserv. Recycl. 1999, 27, 105–115. [Google Scholar] [CrossRef]
- Banaee, M.; Sureda, A.; Zohiery, F.; Hagi, B.N.; Garanzini, D.S. Alterations in biochemical parameters of the freshwater fish, Alburnus mossulensis, exposed to sub-lethal concentrations of Fenpropathrin. Int. J. Aquat. Bio. 2014, 2, 58–68. [Google Scholar]
- Yu, T.; Xu, X.; Mao, H.; Han, X.; Liu, Y.; Zhang, H.; Lai, J.; Gu, J.; Xia, M.; Hu, C.; et al. Fenpropathrin exposure induces neurotoxicity in zebrafish embryos. Fish Physiol. Biochem. 2022, 48, 1539–1554. [Google Scholar] [CrossRef]
- Sarmah, R.; Pokhrel, H.; Ameen, R.; Nath, D.; Bhagabati, S.K.; Dutta, R. Comparative toxicity analysis of fenpropathrin with its two commercial formulations on developing zebrafish embryos. Turk. J. Fish. Aquat. Sci. 2023, 23, 23405. [Google Scholar] [CrossRef]
- Authman, M.M.; Zaki, M.S.; Khallaf, E.A.; Abbas, H.H. Use of fish as bio-indicator of the effects of heavy metals pollution. J. Aquac. Res. Dev. 2015, 6, 1000328. [Google Scholar] [CrossRef]
- Miyamoto, J. Degradation, metabolism and toxicity of synthetic pyrethroids. Environ. Health Perspect. 1976, 14, 15–28. [Google Scholar] [CrossRef]
- Ahmad, F.; Ali, S.S.; Shakoori, A.R. Sublethal effects of Danitol (Fenpropathrin), a synthetic pyrethroid, on Chinese grass carp, Ctenopharyngodon idella. Folia Biol. 1995, 43, 151–160. [Google Scholar]
- Wang, Y.; Yang, G.; Shen, W.; Xu, C.; Wang, Q. Synergistic effect of fenpropathrin and paclobutrazol on early life stages of zebrafish (Danio rerio). Environ. Pollut. 2020, 266, 115067. [Google Scholar] [CrossRef]
- Ma, J.; Xiu, W.; Diao, C.; Miao, Y.; Feng, Y.; Ding, W.; Li, Y.; Sultan, Y.; Li, X. Fenpropathrin induces neurotoxic effects in common carp (Cyprinus carpio L.). Pestic. Biochem. Physiol. 2023, 197, 105644. [Google Scholar] [CrossRef]
- Tlili, S.; Jebali, J.; Banni, M.; Haouas, Z.; Mlayah, A.; Helal, A.N.; Boussetta, H. Multimarker approach analysis in common carp Cyprinus carpio sampled from three freshwater sites. Environ. Monit. Assess. 2010, 168, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Y.; Li, W.; Li, X. Hepatotoxicity of paraquat on common carp (Cyprinus carpio L.). Sci. Total Environ. 2018, 616, 889–898. [Google Scholar] [CrossRef]
- Ma, J.; Li, Y.; Wu, M.; Zhang, C.; Che, Y.; Li, W.; Li, X. Serum immune responses in common carp (Cyprinus carpio L.) to paraquat exposure: The traditional parameters and circulating microRNAs. Fish Shellfish Immun. 2018, 76, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Yki-Järvinen, H. Glucose toxicity. Endocr. Rev. 1992, 13, 415–431. [Google Scholar]
- Borges, A.; Scotti, L.V.; Siqueira, D.R.; Zanini, R.; Amaral, F.; Jurinitz, D.F.; Wassermann, G.F. Changes in hematological and serum biochemical values in jundia Rhamdia quelen due to sub-lethal toxicity of cypermethrin. Chemosphere 2007, 69, 920–926. [Google Scholar] [CrossRef]
- Whalan, J.E. A toxicologist’s guide to clinical pathology in animals. In Hematology, Clinical Chemistry, Urinalysis; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Banihashemi, E.A.; Soltanian, S.; Gholamhosseini, A.; Banaee, M. Effect of microplastics on Yersinia ruckeri infection in rainbow trout (Oncorhynchus mykiss). Environ. Sci. Pollut. Res. Int. 2022, 29, 11939–11950. [Google Scholar] [CrossRef]
- Simons, K.; Ikoen, E. How cells handle cholesterol. Science 2000, 290, 1721–1726. [Google Scholar] [CrossRef]
- Tunçsoy, M. Impacts of titanium dioxide nanoparticles on serum parameters and enzyme activities of Clarias gariepinus. Bull. Environ. Contam. Toxicol. 2021, 106, 629–636. [Google Scholar] [CrossRef]
- Busher, J.T. Serum albumin and globulin. Clin. Methods Hist. Phys. Lab. Exam. 1990, 3, 497–499. [Google Scholar]
- Chatterjee, A.; Bhattacharya, R.; Chatterjee, S.; Saha, N.C. λ cyhalothrin induced toxicity and potential attenuation of hematological, biochemical, enzymological and stress biomarkers in Cyprinus carpio L. at environmentally relevant concentrations: A multiple biomarker approach. Comp. Biochem. Phys. C 2021, 250, 109164. [Google Scholar] [CrossRef]
- Hatami, M.; Banaee, M.; Haghi, B.N. Sub-lethal toxicity of chlorpyrifos alone and in combination with polyethylene glycol to common carp (Cyprinus carpio). Chemosphere 2019, 219, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Navruz, F.Z.; Acar, Ü.; Yılmaz, S.; Kesbiç, O.S. Dose-dependent stress response of esfenvalerate insecticide on common carp (Cyprinus carpio): Evaluating blood parameters and gene expression. Comp. Biochem. Physiol. C 2023, 272, 109711. [Google Scholar] [CrossRef]
- Reddy, C.P.K.; Manikandavelu, D.; Arisekar, U.; Ahilan, B.; Uma, A.; Jayakumar, N.; Kim, W.; Govarthanan, M.; Harini, C.; Vidya, R.S.; et al. Toxicological effect of endocrine disrupting insecticide (deltamethrin) on enzymatical, haematological and histopathological changes in the freshwater iridescent shark, Pangasius hypothalamus. Environ. Toxicol. Pharmacol. 2023, 101, 104201. [Google Scholar] [CrossRef] [PubMed]
- Poupon, R. Liver alkaline phosphatase: A missing link between choleresis and biliary inflammation. Hepatology 2015, 61, 2080–2090. [Google Scholar] [CrossRef]
- Yancheva, V.; Velcheva, I.; Stoyanova, S.; Georgieva, E. Histological biomarkers in fish as a tool in ecological risk assessment and monitoring programs: A review. Appl. Ecol. Environ. Res. 2016, 14, 47–75. [Google Scholar] [CrossRef]
- Khafaga, A.F.; Naiel, M.A.; Dawood, M.A.; Abdel-Latif, H.M. Dietary Origanum vulgare essential oil attenuates cypermethrin-induced biochemical changes, oxidative stress, histopathological alterations, apoptosis, and reduces DNA damage in Common carp (Cyprinus carpio). Aquat. Toxicol. 2020, 228, 105624. [Google Scholar] [CrossRef]
- Afzal, G.; Ahmad, H.I.; Hussain, R.; Jamal, A.; Kiran, S.; Hussain, T.; Saeed, S. Bisphenol A induces histopathological, hematobiochemical alterations, oxidative stress, and genotoxicity in common carp (Cyprinus carpio L.). Oxid. Med. Cell. Longev. 2022, 2022, 5450421. [Google Scholar] [CrossRef]
- Wang, W.; Mou, S.; Xiu, W.; Li, Y.; Liu, Z.; Feng, Y.; Ma, J.; Li, X. Fenpropathrin disrupted the gills of common carp (Cyprinus carpio L.) through oxidative stress, inflammatory responses, apoptosis, and transcriptional alterations. Ecotoxicol. Environ. Saf. 2024, 271, 116007. [Google Scholar] [CrossRef]
- Ma, J.; Zhu, J.; Wang, W.; Ruan, P.; Rajeshkumar, S.; Li, X. Biochemical and molecular impacts of glyphosate-based herbicide on the gills of common carp. Environ. Pollut. 2019, 252, 1288–1300. [Google Scholar] [CrossRef]
- Liu, Z.; Shangguan, Y.; Zhu, P.; Sultan, Y.; Feng, Y.; Li, X.; Ma, J. Developmental toxicity of glyphosate on embryo-larval zebrafish (Danio rerio). Ecotox. Environ. Saf. 2022, 236, 113493. [Google Scholar] [CrossRef]
- Xiu, W.; Ding, W.; Mou, S.; Li, Y.; Sultan, Y.; Ma, J.; Li, X. Adverse effects of fenpropathrin on the intestine of common carp (Cyprinus carpio L.) and the mechanism involved. Pestic. Biochem. Physiol. 2024, 199, 105799. [Google Scholar] [CrossRef] [PubMed]
- Gaweł, S.; Wardas, M.; Niedworok, E.; Wardas, P. Malondialdehyde (MDA) as a lipid peroxidation marker. Wiad. Lek. 2004, 57, 453–455. [Google Scholar] [PubMed]
- Xu, X.; Yu, Y.; Ling, M.; Ares, I.; Martínez, M.; Lopez-Torres, B.; Maximiliano, J.E.; Martínez-Larrañaga, M.R.; Wang, X.; Anadón, A.; et al. Oxidative stress and mitochondrial damage in lambda-cyhalothrin toxicity: A comprehensive review of antioxidant mechanisms. Environ. Pollut. 2023, 338, 122694. [Google Scholar] [CrossRef]
- Dinarello, C.A. Proinflammatory cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef]
- Murray, A.W. Recycling the cell cycle cyclins revisited. Cell 2004, 116, 221–234. [Google Scholar] [CrossRef]
- Xu, R.; Xu, P.; Wei, H.; Huang, Y.; Zhu, X.; Lin, C.; Yan, Z.; Xin, L.; Li, L.; Lv, W.; et al. Ticlopidine induces embryonic development toxicity and hepatotoxicity in zebrafish by upregulating the oxidative stress signaling pathway. Ecotoxicol. Environ. Saf. 2023, 262, 115283. [Google Scholar] [CrossRef]
- Gutierrez, C. DNA replication and cell cycle in plants learning from geminiviruses. EMBO J. 2000, 19, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Romanowski, P.; Madine, M.A. Mechanisms restricting DNA replication to once per cell cycle: MCMS, pre-replicative complexes and kinases. Trends Cell Biol. 1996, 6, 184–188. [Google Scholar] [CrossRef]
- Liu, D.; Keijzers, G.; Rasmussen, L.J. DNA mismatch repair and its many roles in eukaryotic cells. Mutat. Res. 2017, 773, 174–187. [Google Scholar] [CrossRef]
- Kusakabe, M.; Onishi, Y.; Tada, H.; Kurihara, F.; Kusao, K.; Furukawa, M.; Iwai, S.; Yokoi, M.; Sakai, W.; Sugasawa, K. Mechanism and regulation of DNA damage recognition in nucleotide excision repair. Genes Environ. 2019, 41, 2. [Google Scholar] [CrossRef]
- Janus, F.; Albrechtsen, N.; Dornreiter, I.; Wiesmüller, L.; Grosse, F.; Deppert, W. The dual role model for p53 in maintaining genomic integrity. Cell. Mol. Life Sci. 1999, 55, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.B.; Schumacher, B. p53 in the DNA-damage-repair process. Cold Spring Harb. Perspect. Med. 2016, 6, a026070. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Lech, J.J.; Buhler, D.R. Xenobiotics and xenoestrogens in fish: Modulation of cytochrome P450 and carcinogenesis. Mutat. Res. 1998, 399, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Wassmur, B.; Gräns, J.; Kling, P.; Celander, M.C. Interactions of pharmaceuticals and other xenobiotics on hepatic pregnane X receptor and cytochrome P450 3A signaling pathway in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2010, 100, 91–100. [Google Scholar] [CrossRef]
- Xu, X.; Cui, Z.; Wang, X.; Wang, X.; Zhang, S. Toxicological responses on cytochrome P450 and metabolic transferases in liver of goldfish (Carassius auratus) exposed to lead and paraquat. Ecotoxicol. Environ. Saf. 2018, 151, 161–169. [Google Scholar] [CrossRef]
- OECD. Guideline for Testing of Chemicals No. 204: Fish, Prolonged Toxicity Test: 14-Day Study; OECD Publishing: Paris, France, 1984. [Google Scholar]
- Wu, F.; Shao, A.; Xu, X. Simultaneous determination of atrazine, dichloropermethrin, cypermethrin, mepermethrin and permethrin in drinking water by HPLC. Mod. Prevent. Med. 2020, 47, 3619–3622. [Google Scholar]
- Ding, W.; Shangguan, Y.; Zhu, Y.; Sultan, Y.; Feng, Y.; Zhang, B.; Liu, Y.; Ma, J.; Li, X. Negative impacts of microcystin-LR and glyphosate on zebrafish intestine: Linked with gut microbiota and microRNAs? Environ. Pollut. 2021, 286, 117685. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Roberts, A.; Pachter, L. Streaming fragment assignment for real-time analysis of sequencing experiments. Nat. Methods 2013, 10, 71–73. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression of RNA-Seq data at the gene level - the DESeq package. Heidelb. Ger. Eur. Mol. Biol. Lab. 2012, 10, 1000. [Google Scholar]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Yuan, G.L.; Zhu, L.; Li, L.; Pei, C.; Hou, L.; Li, C.; Jiang, X.Y.; Kong, X.H. Molecular characteristics of interleukin (IL)-17A/F3 and its immune response on the pathogen and functional regulation on cytokines in common carp Cyprinus carpio L. Dev. Comp. Immunol. 2023, 139, 104566. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2015, 55, 611–622. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, G.; Liu, Z.; Wang, H.; Mou, S.; Li, Y.; Ma, J.; Li, X. Risk Assessment of Fenpropathrin: Cause Hepatotoxicity and Nephrotoxicity in Common Carp (Cyprinus carpio L.). Int. J. Mol. Sci. 2024, 25, 9822. https://doi.org/10.3390/ijms25189822
Zhu G, Liu Z, Wang H, Mou S, Li Y, Ma J, Li X. Risk Assessment of Fenpropathrin: Cause Hepatotoxicity and Nephrotoxicity in Common Carp (Cyprinus carpio L.). International Journal of Molecular Sciences. 2024; 25(18):9822. https://doi.org/10.3390/ijms25189822
Chicago/Turabian StyleZhu, Gongming, Zhihui Liu, Hao Wang, Shaoyu Mou, Yuanyuan Li, Junguo Ma, and Xiaoyu Li. 2024. "Risk Assessment of Fenpropathrin: Cause Hepatotoxicity and Nephrotoxicity in Common Carp (Cyprinus carpio L.)" International Journal of Molecular Sciences 25, no. 18: 9822. https://doi.org/10.3390/ijms25189822