
Optimization of Helicobacter pylori Biofilm Formation in In Vitro Conditions Mimicking Stomach



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Selection of H. pylori Strain
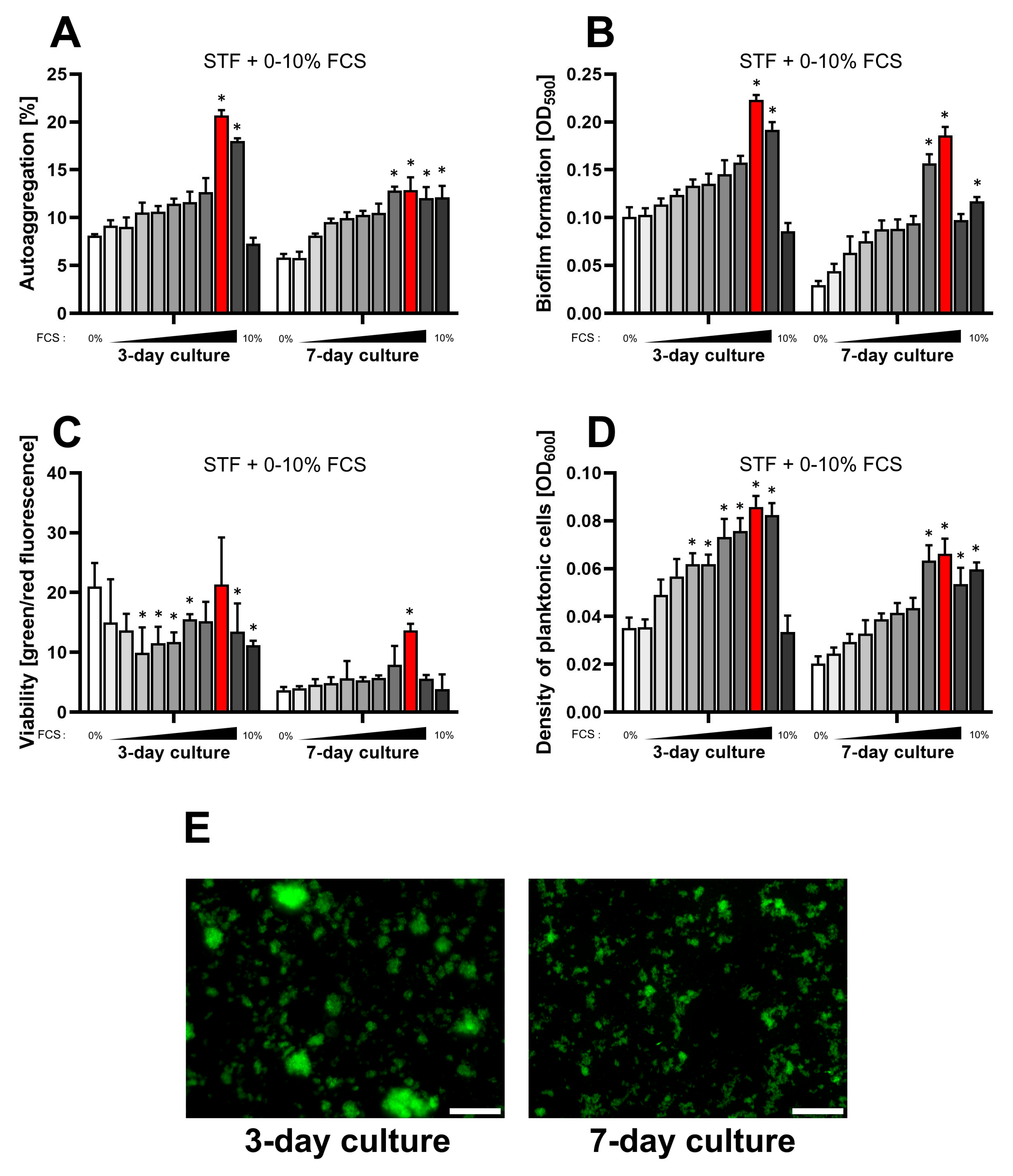
2.2. Effect of Simulated Transcellular Fluid (STF) on the Physiology of H. pylori
2.3. Effect of Simulated Gastric Fluid (SGF) on the Physiology of H. pylori
2.4. Summary of the Impact of STF and SGF on the H. pylori Physiology
2.5. Selection of a Gastric Cell Line
2.6. Establishment of H. pylori Co-Culture with KATO III in a Semi-Permeable Membrane Model
2.7. Establishment of H. pylori Co-Culture with KATO III in a Microfluidic Model
3. Materials and Methods
3.1. Storage and Revival of H. pylori
3.2. Influence of the Composition of Culture Media on the Physiology of H. pylori
3.3. Storage and Revival of KATO III
3.4. Co-Culture of H. pylori with KATO III in a Semi-Permeable Membrane Model
3.5. Co-Culture of H. pylori with KATO III in a Microfluidic System
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malfertheiner, P.; Camargo, M.C.; El-Omar, E.; Liou, J.M.; Peek, R.; Schulz, C.; Smith, S.I.; Suerbaum, S. Helicobacter pylori Infection. Nat. Rev. Dis. Primers 2023, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.T.; Issa, P.P.; Sinnathamby, E.S.; Granier, M.; Mayeux, H.; Eubanks, T.N.; Malone, K.; Ahmadzadeh, S.; Cornett, E.M.; Shekoohi, S.; et al. Helicobacter pylori: A Review of Current Treatment Options in Clinical Practice. Life 2022, 12, 2038. [Google Scholar] [CrossRef]
- Boyanova, L.; Hadzhiyski, P.; Gergova, R.; Markovska, R. Evolution of Helicobacter pylori Resistance to Antibiotics: A Topic of Increasing Concern. Antibiotics 2023, 12, 332. [Google Scholar] [CrossRef]
- Rokkas, T.; Ekmektzoglou, K. Advances in the Pharmacological and Regulatory Management of Multidrug Resistant Helicobacter pylori. Expert Rev. Clin. Pharmacol. 2023, 16, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Mladenova, I. Epidemiology of Helicobacter pylori Resistance to Antibiotics (A Narrative Review). Antibiotics 2023, 12, 1184. [Google Scholar] [CrossRef]
- Dascălu, R.I.; Bolocan, A.; Păduaru, D.N.; Constantinescu, A.; Mitache, M.M.; Stoica, A.D.; Andronic, O. Multidrug Resistance in Helicobacter pylori Infection. Front. Microbiol. 2023, 14, 1128497. [Google Scholar] [CrossRef] [PubMed]
- Moss, S.F.; Shah, S.C.; Tan, M.C.; El-Serag, H.B. Evolving Concepts in Helicobacter pylori Management. Gastroenterology 2024, 166, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Tshibangu-Kabamba, E.; Yamaoka, Y. Helicobacter pylori Infection and Antibiotic Resistance—From Biology to Clinical Implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 613–629. [Google Scholar] [CrossRef]
- Zhao, A.; Sun, J.; Liu, Y. Understanding Bacterial Biofilms: From Definition to Treatment Strategies. Front. Cell. Infect. Microbiol. 2023, 13, 1137947. [Google Scholar] [CrossRef]
- Flemming, H.C.; van Hullebusch, E.D.; Neu, T.R.; Nielsen, P.H.; Seviour, T.; Stoodley, P.; Wingender, J.; Wuertz, S. The Biofilm Matrix: Multitasking in a Shared Space. Nat. Rev. Microbiol. 2022, 21, 70–86. [Google Scholar] [CrossRef]
- Krzyżek, P.; Grande, R.; Migdał, P.; Paluch, E.; Gościniak, G. Biofilm Formation as a Complex Result of Virulence and Adaptive Responses of Helicobacter pylori. Pathogens 2020, 9, 1062. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Servetas, S.L.; Windham, I.; Merrell, D.S.; Ottemann, K.M. Helicobacter pylori Biofilm Formation and Its Potential Role in Pathogenesis. Microbiol. Mol. Biol. Rev. 2018, 82, e00001-18. [Google Scholar] [CrossRef] [PubMed]
- Rizzato, C.; Torres, J.; Kasamatsu, E.; Camorlinga-Ponce, M.; Bravo, M.M.; Canzian, F.; Kato, I. Potential Role of Biofilm Formation in the Development of Digestive Tract Cancer with Special Reference to Helicobacter pylori Infection. Front. Microbiol. 2019, 10, 846. [Google Scholar] [CrossRef] [PubMed]
- Elshenawi, Y.; Hu, S.; Hathroubi, S. Biofilm of Helicobacter pylori: Life Cycle, Features, and Treatment Options. Antibiotics 2023, 12, 1260. [Google Scholar] [CrossRef]
- Hou, C.; Yin, F.; Wang, S.; Zhao, A.; Li, Y.; Liu, Y. Helicobacter pylori Biofilm-Related Drug Resistance and New Developments in Its Anti-Biofilm Agents. Infect. Drug Resist. 2022, 15, 1561–1571. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Grande, R.; Gościniak, G. Biofilm Formation of Helicobacter pylori in Both Static and Microfluidic Conditions Is Associated with Resistance to Clarithromycin. Front. Cell. Infect. Microbiol. 2022, 12, 868905. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Gościniak, G. Impact of Sub-Inhibitory Concentrations of Antibiotics on the Biofilm Development of Helicobacter pylori. Abstr. Microb. Health Dis. 2023, 5, e857, P06.01. [Google Scholar]
- Yonezawa, H.; Osaki, T.; Hojo, F.; Kamiya, S. Effect of Helicobacter pylori Biofilm Formation on Susceptibility to Amoxicillin, Metronidazole and Clarithromycin. Microb. Pathog. 2019, 132, 100–108. [Google Scholar] [CrossRef]
- Hathroubi, S.; Zerebinski, J.; Clarke, A.; Ottemann, K.M. Helicobacter pylori Biofilm Confers Antibiotic Tolerance in Part via a Protein-Dependent Mechanism. Antibiotics 2020, 9, 355. [Google Scholar] [CrossRef]
- Fauzia, K.A.; Miftahussurur, M.; Syam, A.F.; Waskito, L.A.; Doohan, D.; Rezkitha, Y.A.A.; Matsumoto, T.; Tuan, V.P.; Akada, J.; Yonezawa, H.; et al. Biofilm Formation and Antibiotic Resistance Phenotype of Helicobacter pylori Clinical Isolates. Toxins 2020, 12, 473. [Google Scholar] [CrossRef]
- Wu, X.; Wu, D.; Cui, G.; Lee, K.H.; Yang, T.; Zhang, Z.; Liu, Q.; Zhang, J.; Chua, E.G.; Chen, Z. Association Between Biofilm Formation and Structure and Antibiotic Resistance in H. pylori. Infect. Drug Resist. 2024, 17, 2512. [Google Scholar] [CrossRef] [PubMed]
- Cammarota, G.; Branca, G.; Ardito, F.; Sanguinetti, M.; Ianiro, G.; Cianci, R.; Torelli, R.; Masala, G.; Gasbarrini, A.; Fadda, G.; et al. Biofilm Demolition and Antibiotic Treatment to Eradicate Resistant Helicobacter pylori: A Clinical Trial. Clin. Gastroenterol. Hepatol. 2010, 8, 817–820.e3. [Google Scholar] [CrossRef] [PubMed]
- Lebeaux, D.; Chauhan, A.; Rendueles, O.; Beloin, C. From In Vitro to In Vivo Models of Bacterial Biofilm-Related Infections. Pathogens 2013, 2, 288–356. [Google Scholar] [CrossRef] [PubMed]
- Vyas, H.K.N.; Xia, B.; Mai-Prochnow, A. Clinically Relevant In Vitro Biofilm Models: A Need to Mimic and Recapitulate the Host Environment. Biofilm 2022, 4, 100069. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Roy, S.; Ghosh, D.; Nandi, S.K. Role of Animal Models in Biomedical Research: A Review. Lab. Anim. Res. 2022, 38, 18. [Google Scholar] [CrossRef]
- Ersoy, S.C.; Heithoff, D.M.; Barnes, L.; Tripp, G.K.; House, J.K.; Marth, J.D.; Smith, J.W.; Mahan, M.J. Correcting a Fundamental Flaw in the Paradigm for Antimicrobial Susceptibility Testing. EBioMedicine 2017, 20, 181. [Google Scholar] [CrossRef]
- Belanger, C.R.; Hancock, R.E.W. Testing Physiologically Relevant Conditions in Minimal Inhibitory Concentration Assays. Nat. Protoc. 2021, 16, 3761–3774. [Google Scholar] [CrossRef]
- O’Gara, E.A.; Maslin, D.J.; Nevill, A.M.; Hill, D.J. The Effect of Simulated Gastric Environments on the Anti-Helicobacter Activity of Garlic Oil. J. Appl. Microbiol. 2008, 104, 1324–1331. [Google Scholar] [CrossRef]
- Liu, T.; Chai, S.; Li, M.; Chen, X.; Xie, Y.; Zhao, Z.; Xie, J.; Yu, Y.; Gao, F.; Zhu, F.; et al. A Nanoparticle-Based Sonodynamic Therapy Reduces Helicobacter pylori Infection in Mouse Without Disrupting Gut Microbiota. Nat. Commun. 2024, 15, 844. [Google Scholar] [CrossRef]
- Holz, C.; Busjahn, A.; Mehling, H.; Arya, S.; Boettner, M.; Habibi, H.; Lang, C. Significant Reduction in Helicobacter pylori Load in Humans with Non-viable Lactobacillus reuteri DSM17648: A Pilot Study. Probiotics Antimicrob. Proteins 2015, 7, 91–100. [Google Scholar] [CrossRef]
- Celli, J.P.; Turner, B.S.; Afdhal, N.H.; Keates, S.; Ghiran, I.; Kelly, C.P.; Ewoldt, R.H.; McKinley, G.H.; So, P.; Erramilli, S.; et al. Helicobacter pylori Moves through Mucus by Reducing Mucin Viscoelasticity. Proc. Natl. Acad. Sci. USA 2009, 106, 14321–14326. [Google Scholar] [CrossRef]
- Henderson, M.A.; Gillon, S.; Al-Haddad, M. Organization and Composition of Body Fluids. Anaesth. Intensive Care Med. 2021, 22, 511–517. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Q.-L.; Cheng, D.-D.; Xu, W.-T.; Lu, N.-H. Adhesion and Invasion of Gastric Mucosa Epithelial Cells by Helicobacter pylori. Front. Cell. Infect. Microbiol. 2016, 6, 159. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.M.; Gaddy, J.A.; Cover, T.L. Alterations in Helicobacter pylori Triggered by Contact with Gastric Epithelial Cells. Front. Cell. Infect. Microbiol. 2012, 2, 17. [Google Scholar] [CrossRef]
- Wood, J.M. Bacterial Responses to Osmotic Challenges. J. Gen. Physiol. 2015, 145, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-Mediated Signalling in Bacteria. FEMS Microbiol. Rev. 2013, 37, 156–181. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.K.; Huang, J.Y.; Wreden, C.; Sweeney, E.G.; Goers, J.; Remington, S.J.; Guillemin, K. Chemorepulsion from the Quorum Signal Autoinducer-2 Promotes Helicobacter pylori Biofilm Dispersal. mBio 2015, 6, e00379. [Google Scholar] [CrossRef]
- Rader, B.A.; Wreden, C.; Hicks, K.G.; Sweeney, E.G.; Ottemann, K.M.; Guillemin, K. Helicobacter pylori Perceives the Quorum-Sensing Molecule AI-2 as a Chemorepellent via the Chemoreceptor TlpB. Microbiology 2011, 157, 2445–2455. [Google Scholar] [CrossRef]
- Sweeney, E.G.; Nishida, A.; Weston, A.; Bañuelos, M.S.; Potter, K.; Conery, J.; Guillemin, K. Agent-Based Modeling Demonstrates How Local Chemotactic Behavior Can Shape Biofilm Architecture. mSphere 2019, 4, e00285-19. [Google Scholar] [CrossRef]
- Martinsen, T.C.; Bergh, K.; Waldum, H.L. Gastric Juice: A Barrier Against Infectious Diseases. Basic Clin. Pharmacol. Toxicol. 2005, 96, 94–102. [Google Scholar] [CrossRef]
- Martinsen, T.C.; Fossmark, R.; Waldum, H.L. The Phylogeny and Biological Function of Gastric Juice—Microbiological Consequences of Removing Gastric Acid. Int. J. Mol. Sci. 2019, 20, 6031. [Google Scholar] [CrossRef] [PubMed]
- Su-Arcaro, C.; Liao, W.; Bieniek, K.; Constantino, M.A.; Decker, S.M.; Turner, B.S.; Bansil, R. Unraveling the Intertwined Effect of pH on Helicobacter pylori Motility and the Microrheology of the Mucin-Based Medium It Swims in. Microorganisms 2023, 11, 2745. [Google Scholar] [CrossRef] [PubMed]
- Salama, N.R.; Hartung, M.L.; Müller, A. Life in the Human Stomach: Persistence Strategies of the Bacterial Pathogen Helicobacter pylori. Nat. Rev. Microbiol. 2013, 11, 385–399. [Google Scholar] [CrossRef]
- Cheng, H.M.; Mah, K.K.; Seluakumaran, K. Gastric Mucosal Barrier, Helicobacter pylori. In Defining Physiology: Principles, Themes, Concepts; Springer: Cham, Switzerland, 2020; Volume 2, pp. 19–20. ISBN 978-3-030-62285-5. [Google Scholar]
- Lindén, S.; Mahdavi, J.; Hedenbro, J.; Borén, T.; Carlstedt, I. Effects of pH on Helicobacter pylori Binding to Human Gastric Mucins: Identification of Binding to Non-MUC5AC Mucins. Biochem. J. 2004, 384, 270. [Google Scholar] [CrossRef]
- Grande, R.; Di Giulio, M.; Bessa, L.J.; Di Campli, E.; Baffoni, M.; Guarnieri, S.; Cellini, L. Extracellular DNA in Helicobacter pylori Biofilm: A Backstairs Rumour. J. Appl. Microbiol. 2011, 110, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori Biofilm Involves a Multigene Stress-Biased Response, Including a Structural Role for Flagella. MBio 2018, 9, e01973-18. [Google Scholar] [CrossRef]
- Windham, I.H.; Servetas, S.L.; Whitmire, J.M.; Pletzer, D.; Hancock, R.E.W.; Merrell, D.S. Helicobacter pylori Biofilm Formation Is Differentially Affected by Common Culture Conditions, and Proteins Play a Central Role in the Biofilm Matrix. Appl. Environ. Microbiol. 2018, 84, e00391-18. [Google Scholar] [CrossRef]
- Rosenberg, M.; Azevedo, N.F.; Ivask, A. Propidium Iodide Staining Underestimates Viability of Adherent Bacterial Cells. Sci. Rep. 2019, 9, 6483. [Google Scholar] [CrossRef]
- Finkel, S.E.; Kolter, R. DNA as a Nutrient: Novel Role for Bacterial Competence Gene Homologs. J. Bacteriol. 2001, 183, 6293. [Google Scholar] [CrossRef]
- Corbinais, C.; Mathieu, A.; Damke, P.P.; Kortulewski, T.; Busso, D.; Prado-Acosta, M.; Radicella, J.P.; Marsin, S. ComB Proteins Expression Levels Determine Helicobacter pylori Competence Capacity. Sci. Rep. 2017, 7, 41495. [Google Scholar] [CrossRef]
- Salgar-Chaparro, S.J.; Lepkova, K.; Pojtanabuntoeng, T.; Darwin, A.; Machuca, L.L. Nutrient Level Determines Biofilm Characteristics and Subsequent Impact on Microbial Corrosion and Biocide Effectiveness. Appl. Environ. Microbiol. 2020, 86, e02885-19. [Google Scholar] [CrossRef] [PubMed]
- Van de Bovenkamp, J.H.B.; Mahdavi, J.; Korteland-Van Male, A.M.; Büller, H.A.; Einerhand, A.W.C.; Borén, T.; Dekker, J. The MUC5AC Glycoprotein Is the Primary Receptor for Helicobacter pylori in the Human Stomach. Helicobacter 2003, 8, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Niv, Y. Helicobacter pylori and Gastric Mucin Expression: A Systematic Review and Meta-Analysis. World J. Gastroenterol. 2015, 21, 9436. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, M.; Sakakibara, K.; Fujii, G. Establishment of Cultured Cell Lines Derived from a Human Gastric Carcinoma. Jpn. J. Exp. Med. 1978, 48, 61–68. [Google Scholar]
- Matsuda, K.; Yamauchi, K.; Matsumoto, T.; Sano, K.; Yamaoka, Y.; Ota, H. Quantitative Analysis of the Effect of Helicobacter pylori on the Expressions of SOX2, CDX2, MUC2, MUC5AC, MUC6, TFF1, TFF2, and TFF3 mRNAs in Human Gastric Carcinoma Cells. Scand. J. Gastroenterol. 2008, 43, 33. [Google Scholar] [CrossRef]
- Perrais, M.; Pigny, P.; Buisine, M.P.; Porchet, N.; Aubert, J.P.; Van Seuningen-Lempire, I. Aberrant Expression of Human Mucin Gene MUC5B in Gastric Carcinoma and Cancer Cells. Identification and Regulation of a Distal Promoter. J. Biol. Chem. 2001, 276, 15386–15396. [Google Scholar] [CrossRef]
- Chung, H.H.; Mireles, M.; Kwarta, B.J.; Gaborski, T.R. Use of Porous Membranes in Tissue Barrier and Co-Culture Models. Lab Chip 2018, 18, 1689. [Google Scholar] [CrossRef]
- Johnson, K.S.; Ottemann, K.M. Colonization, Localization, and Inflammation: The Roles of H. pylori Chemotaxis In Vivo. Curr. Opin. Microbiol. 2018, 41, 51–57. [Google Scholar] [CrossRef]
- Li, R.; Lv, X.; Zhang, X.; Saeed, O.; Deng, Y. Microfluidics for Cell-Cell Interactions: A Review. Front. Chem. Sci. Eng. 2016, 10, 90–98. [Google Scholar] [CrossRef]
- Coluccio, M.L.; Perozziello, G.; Malara, N.; Parrotta, E.; Zhang, P.; Gentile, F.; Limongi, T.; Raj, P.M.; Cuda, G.; Candeloro, P.; et al. Microfluidic Platforms for Cell Cultures and Investigations. Microelectron. Eng. 2019, 208, 14–28. [Google Scholar] [CrossRef]
- Badilescu, S.; Parimalam, S.S.; Packirisamy, M. Culturing Cells for Life: Innovative Approaches in Macroscopic and Microfluidic Cultures, with an Emphasis on Stem Cells. Front. Nanotechnol. 2023, 5, 1264498. [Google Scholar] [CrossRef]
- Benoit, M.R.; Conant, C.G.; Ionescu-Zanetti, C.; Schwartz, M.; Matin, A. New Device for High-Throughput Viability Screening of Flow Biofilms. Appl. Environ. Microbiol. 2010, 76, 4136–4142. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Pocard, M.; Mirshahi, M. Targeting the Differentiation of Gastric Cancer Cells (KATO-III) Downregulates Epithelial-Mesenchymal and Cancer Stem Cell Markers. Oncol. Rep. 2019, 42, 678. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, M.; Krzyżek, P.; Dworniczek, E.; Adamski, R.; Sroka, Z. In Silico Screening and In Vitro Assessment of Natural Products with Anti-Virulence Activity against Helicobacter pylori. Molecules 2022, 27, 20. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Guo, Z.; Yan, J.; Bu, C.; Peng, C.; Li, C.; Mao, R.; Zhang, J.; Wang, Z.; Chen, S.; et al. Gastric Acid-Responsive ROS Nanogenerators for Effective Treatment of Helicobacter pylori Infection without Disrupting Homeostasis of Intestinal Flora. Adv. Sci. 2023, 10, e2206957. [Google Scholar] [CrossRef]
- Hałasa, R.; Turecka, K.; Mizerska, U.; Krauze-Baranowska, M. Anti-Helicobacter pylori Biofilm Extracts from Rubus idaeus and Rubus occidentalis. Pharmaceutics 2024, 16, 501. [Google Scholar] [CrossRef]
- Hochvaldova, L.; Posselt, G.; Wessler, S.; Kvítek, L.; Panáček, A. Implications of Silver Nanoparticles for H. pylori Infection: Modulation of CagA Function and Signaling. Front. Cell. Infect. Microbiol. 2024, 14, 1419568. [Google Scholar] [CrossRef]
- Krzyżek, P.; Migdał, P.; Paluch, E.; Karwańska, M.; Wieliczko, A.; Gościniak, G. Myricetin as an Antivirulence Compound Interfering with a Morphological Transformation into Coccoid Forms and Potentiating Activity of Antibiotics against Helicobacter pylori. Int. J. Mol. Sci. 2021, 22, 2695. [Google Scholar] [CrossRef]
- Chen, X.; Chen, W.; Zhao, Y.; Wang, Q.; Wang, W.; Xiang, Y.; Yuan, H.; Xie, Y.; Zhou, J. Interplay of Helicobacter pylori, Fibroblasts, and Cancer Cells Induces Fibroblast Activation and Serpin E1 Expression by Cancer Cells to Promote Gastric Tumorigenesis. J. Transl. Med. 2022, 20, 322. [Google Scholar] [CrossRef]
- Hathroubi, S.; Hu, S.; Ottemann, K.M. Genetic Requirements and Transcriptomics of Helicobacter pylori Biofilm Formation on Abiotic and Biotic Surfaces. npj Biofilms Microbiomes 2020, 6, 56. [Google Scholar] [CrossRef]
- Gajewski, A.; Mnich, E.; Szymański, K.; Hinc, K.; Obuchowski, M.; Moran, A.P.; Chmiela, M. Helicobacter pylori Antigens, Acetylsalicylic Acid, LDL and 7-ketocholesterol—Their Potential Role in Destabilizing the Gastric Epithelial Cell Barrier. An In Vitro Model of Kato III Cells. Acta Biochim. Pol. 2016, 63, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K.; Tanaka, T.; McNeil, P.L. Disruption-Induced Mucus Secretion: Repair and Protection. PLoS Biol. 2006, 4, 1525–1531. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, U.; Duku, M.; Katoh, M.; Jörns, J.; Krause, W.J. Histochemical Similarities of Mucins Produced by Brunner’s Glands and Pyloric Glands: A Comparative Study. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 2004, 278, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Duda-Madej, A.; Kozłowska, J.; Baczyńska, D.; Krzyżek, P. Ether Derivatives of Naringenin and Their Oximes as Factors Modulating Bacterial Adhesion. Antibiotics 2023, 12, 1076. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyżek, P.; Migdał, P.; Krzyżanowska, B.; Duda-Madej, A. Optimization of Helicobacter pylori Biofilm Formation in In Vitro Conditions Mimicking Stomach. Int. J. Mol. Sci. 2024, 25, 9839. https://doi.org/10.3390/ijms25189839
Krzyżek P, Migdał P, Krzyżanowska B, Duda-Madej A. Optimization of Helicobacter pylori Biofilm Formation in In Vitro Conditions Mimicking Stomach. International Journal of Molecular Sciences. 2024; 25(18):9839. https://doi.org/10.3390/ijms25189839
Chicago/Turabian StyleKrzyżek, Paweł, Paweł Migdał, Barbara Krzyżanowska, and Anna Duda-Madej. 2024. "Optimization of Helicobacter pylori Biofilm Formation in In Vitro Conditions Mimicking Stomach" International Journal of Molecular Sciences 25, no. 18: 9839. https://doi.org/10.3390/ijms25189839