
Integrated Metabolome, Transcriptome and Long Non-Coding RNA Analysis Reveals Potential Molecular Mechanisms of Sweet Cherry Fruit Ripening

 , ,
, ,
Abstract
1. Introduction
2. Results
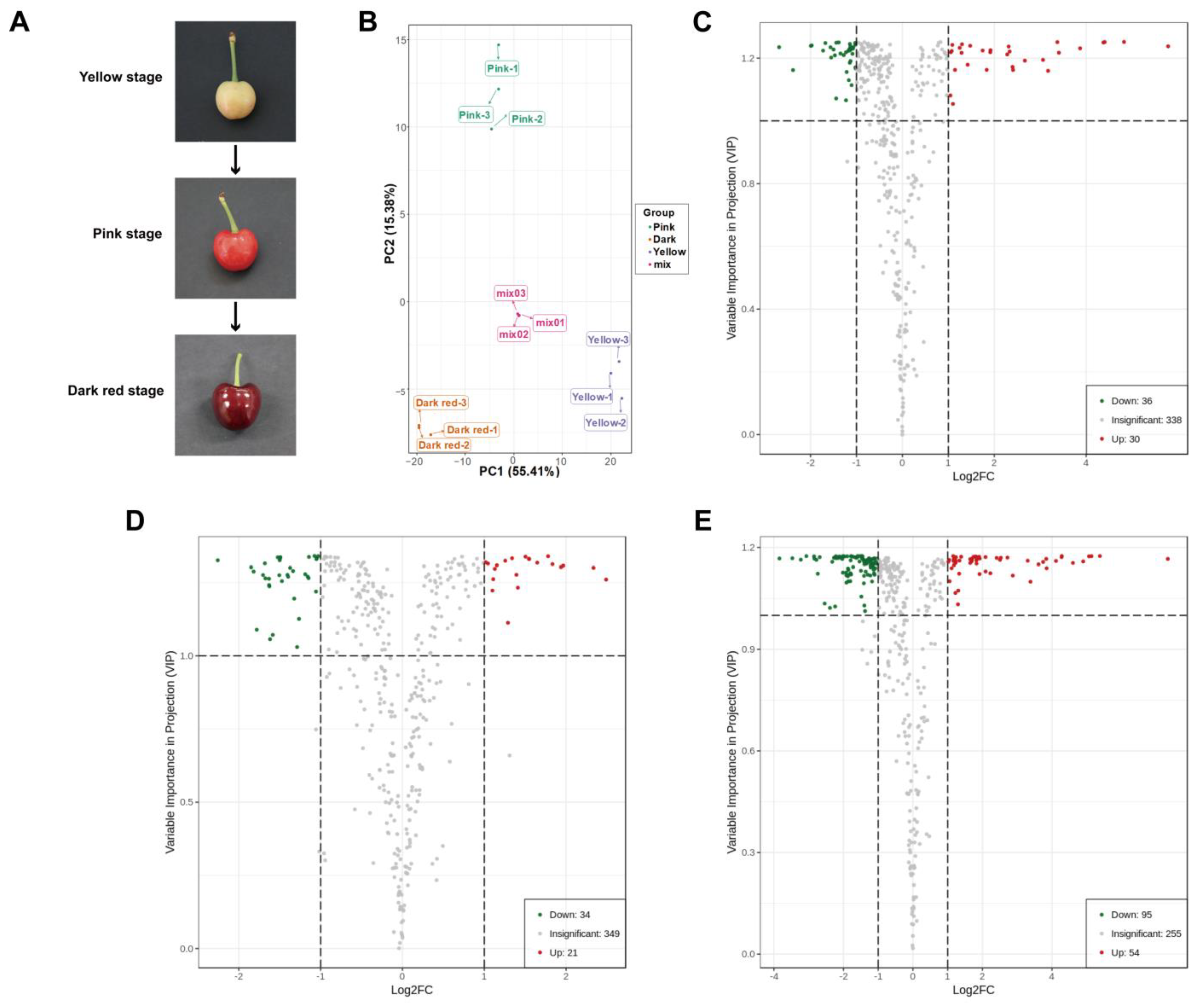
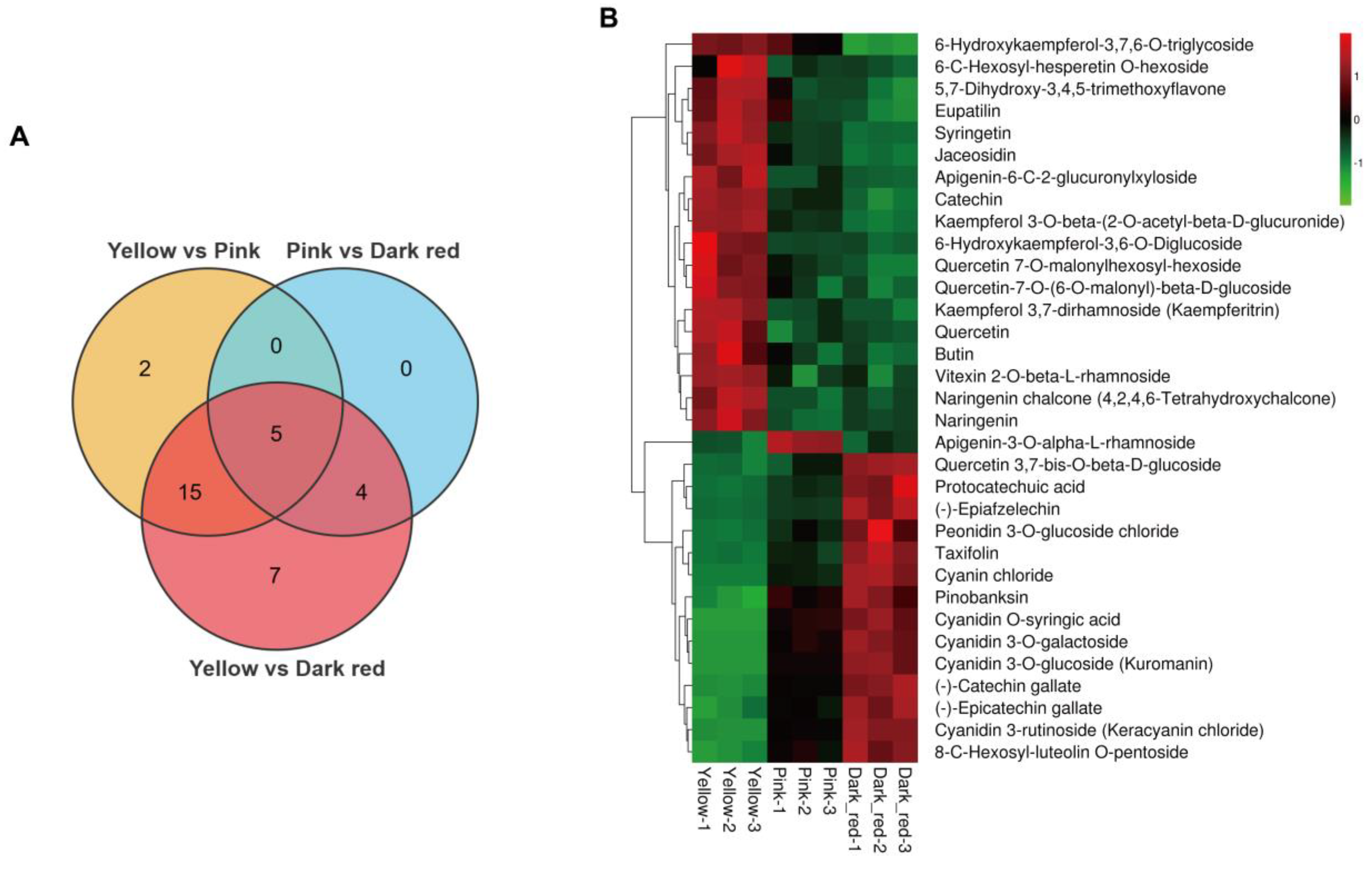
2.1. Widely Targeted Metabolomics Analysis
2.2. Statistics of Transcriptome and lncRNA Data
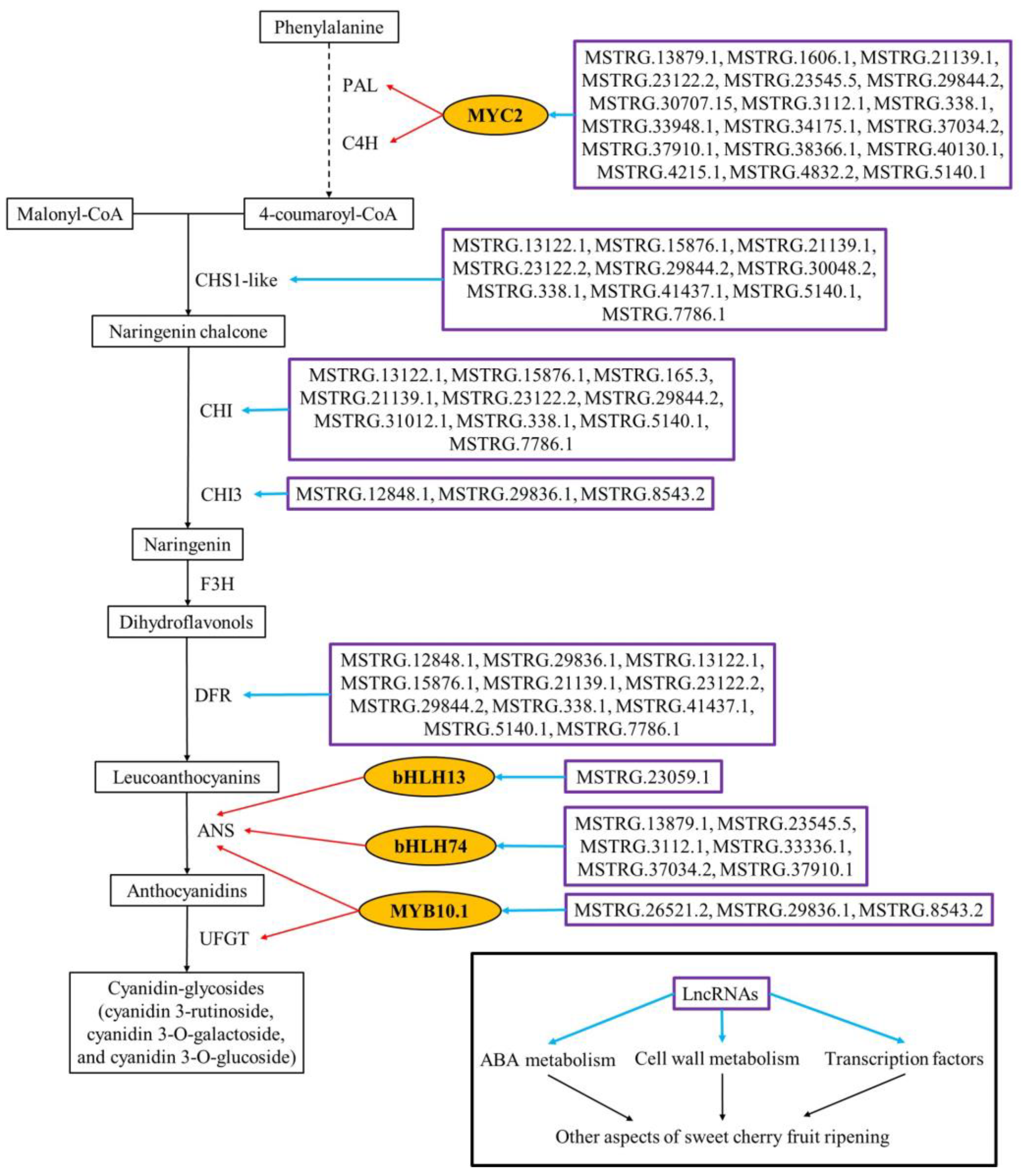
2.3. Spatio-Temporal Specific Expression of Ripening-Associated Genes Drives Sweet Cherry Fruit Ripening
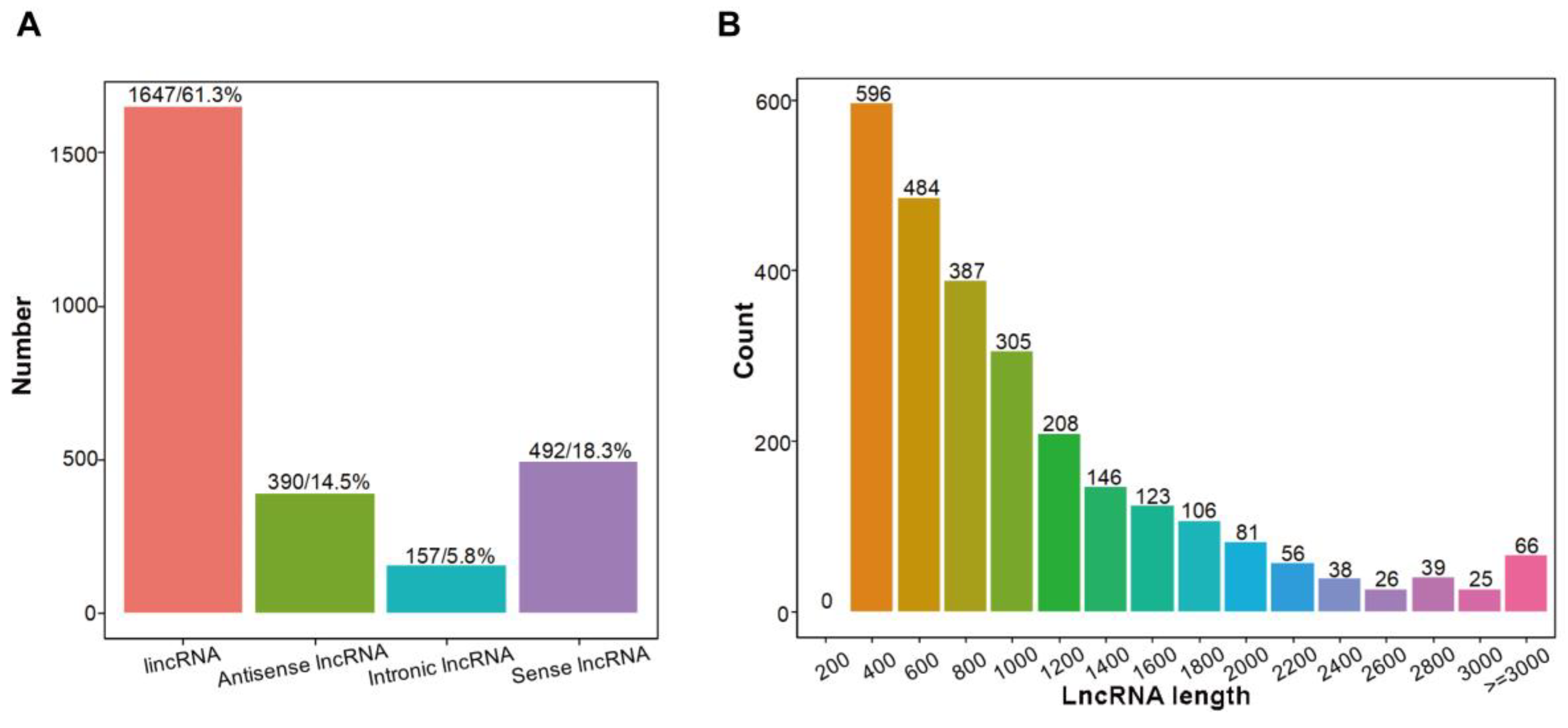
2.4. Identification of lncRNAs in Sweet Cherry Fruit
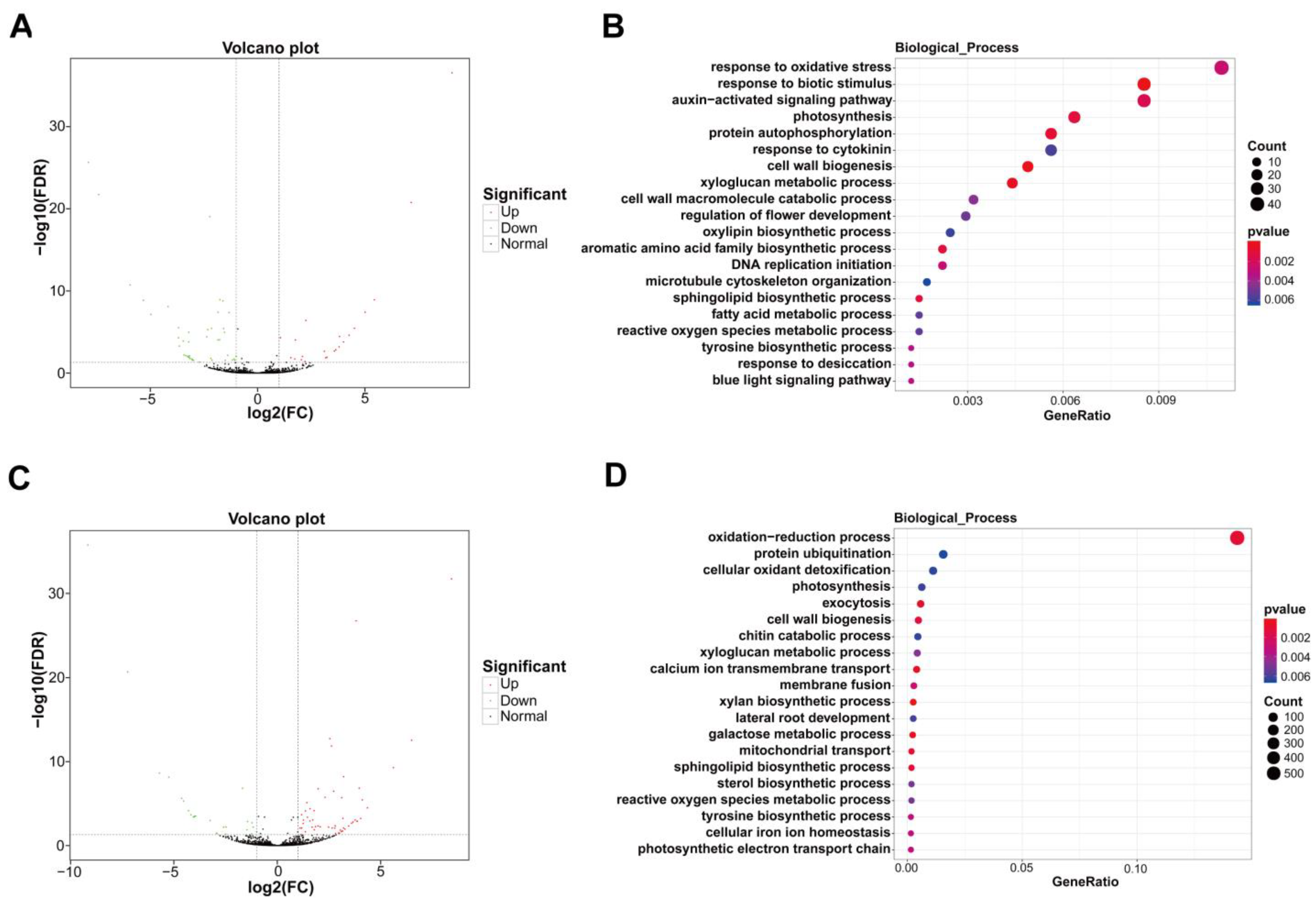
2.5. Differential Expression and Target Gene Enrichment of lncRNAs
2.6. Verification of Transcriptome and lncRNA Data
2.7. lncRNAs Were Involved in the Ripening Process of Sweet Cherry Fruit
2.8. Pearson’s Correlation Analysis of Anthocyanin Metabolites and Associated DEGs/DE-lncRNAs
3. Discussion
4. Materials and Methods
4.1. Fruit Material
4.2. Widely Targeted Metabolomics Detection
4.3. Transcriptome and lncRNA Sequencing
4.4. RT-qPCR
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Correia, S.; Schouten, R.; Silva, A.P.; Gonçalves, B. Factors affecting quality and health promoting compounds during growth and postharvest life of sweet cherry (Prunus avium L.). Front. Plant Sci. 2017, 8, 2166. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Li, W.S.; Chang, H.; Zhou, J.H.; Luo, Y.B.; Zhang, K.C.; Wang, B.G. Sweet cherry fruit miRNAs and effect of high CO2 on the profile associated with ripening. Planta 2019, 249, 1799–1810. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.L.; Dong, Y.X.; Liu, C.L.; Song, L.L.; Chen, L.; Li, M. The PavNAC56 transcription factor positively regulates fruit ripening and softening in sweet cherry (Prunus avium). Physiol. Plant. 2022, 174, e13834. [Google Scholar] [CrossRef]
- Giovannoni, J.; Nguyen, C.; Ampofo, B.; Zhong, S.L.; Fei, Z.J. The epigenome and transcriptional dynamics of fruit ripening. Annu. Rev. Genet. 2017, 68, 61–84. [Google Scholar] [CrossRef]
- Wang, W.; Fan, D.Y.; Hao, Q.; Jia, W.S. Signal transduction in non-climacteric fruit ripening. Hortic. Res. 2022, 9, uhac190. [Google Scholar] [CrossRef]
- Zhang, X.; Du, M.J.; Yang, Z.F.; Wang, Z.J.; Lim, K.J. Biogenesis, mode of action and the interactions of plant non-coding RNAs. Int. J. Mol. Sci. 2023, 24, 10664. [Google Scholar] [CrossRef]
- Chen, L.; Zhu, Q.H.; Kaufmann, K. Long non-coding RNAs in plants: Emerging modulators of gene activity in development and stress responses. Planta 2020, 252, 92. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, S.; Qi, H.R.; Cai, H.; Xu, M. Research progress on plant long non-coding RNA. Plants 2020, 9, 408. [Google Scholar] [CrossRef]
- Palos, K.; Yu, L.; Railey, C.E.; Dittrich, A.C.N.; Nelson, A.D.L. Linking discoveries, mechanisms, and technologies to develop a clearer perspective on plant long noncoding RNAs. Plant Cell 2023, 35, 1762–1786. [Google Scholar] [CrossRef]
- Zhu, B.Z.; Yang, Y.F.; Li, R.; Fu, D.Q.; Wen, L.W.; Luo, Y.B.; Zhu, H.L. RNA sequencing and functional analysis implicate the regulatory role of long non-coding RNAs in tomato fruit ripening. J. Exp. Bot. 2015, 66, 4483–4495. [Google Scholar] [CrossRef]
- Tang, W.; Zheng, Y.; Dong, J.; Yu, J.; Yue, J.Y.; Liu, F.F.; Guo, X.H.; Huang, S.X.; Wisniewski, M.; Sun, J.Q.; et al. Comprehensive transcriptome profiling reveals long noncoding RNA expression and alternative splicing regulation during fruit development and ripening in kiwifruit (Actinidia chinensis). Front. Plant Sci. 2016, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Y.; Chen, D.G.; Zhang, T.; Duan, A.G.; Zhang, J.G.; He, C.Y. Transcriptomic and functional analyses unveil the role of long non-coding RNAs in anthocyanin biosynthesis during sea buckthorn fruit ripening. DNA Res. 2018, 25, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.H.; Wang, Y.X.; Zhu, B.Z.; Luo, Y.; Wang, Q.; Gao, L.P. Analysis of the coding and non-coding RNA transcriptomes in response to bell pepper chilling. Int. J. Mol. Sci. 2018, 19, 2001. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.X.; Jiang, L.Y.; Chen, Q.; Li, Y.L.; Zhang, Y.T.; Luo, Y.; Zhang, Y.; Sun, B.; Wang, X.R.; Tang, H.R. Comparative transcriptome profiling analysis of red- and white-fleshed strawberry (Fragaria × ananassa) provides new insight into the regulation of the anthocyanin pathway. Plant Cell Physiol. 2018, 59, 1844–1859. [Google Scholar] [CrossRef]
- Liu, W.H.; Cheng, C.Z.; Lin, Y.L.; Xu, X.H.; Lai, Z.X. Genome-wide identification and characterization of mRNAs and lncRNAs involved in cold stress in the wild banana (Musa itinerans). PLoS ONE 2018, 13, e0200002. [Google Scholar] [CrossRef]
- Yang, T.; Ma, H.Y.; Zhang, J.; Wu, T.; Song, T.T.; Tian, J.; Yao, Y.C. Systematic identification of long noncoding RNAs expressed during light-induced anthocyanin accumulation in apple fruit. Plant J. 2019, 100, 572–590. [Google Scholar] [CrossRef]
- Tian, Y.Y.; Bai, S.L.G.; Dang, Z.H.; Hao, J.F.; Zhang, J.; Hasi, A. Genome-wide identification and characterization of long non-coding RNAs involved in fruit ripening and the climacteric in Cucumis melo. BMC Plant Biol. 2019, 19, 369. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, G.; Sharma, S.; Upadhyay, S.K.; Singh, K. Long non-coding RNAs coordinate developmental transitions and other key biological processes in grapevine. Sci. Rep. 2019, 9, 3552. [Google Scholar] [CrossRef]
- Wu, Y.W.; Xu, J.; Han, X.M.; Qiao, G.; Yang, K.; Wen, Z.; Wen, X.P. Comparative transcriptome analysis combining SMRT- and Illumina-based RNA-Seq identifies potential candidate genes involved in betalain biosynthesis in pitaya fruit. Int. J. Mol. Sci. 2020, 21, 3288. [Google Scholar] [CrossRef]
- Yao, S.X.; Wang, Z.M.; Cao, Q.; Xie, J.; Wang, X.R.; Zhang, R.; Deng, L.L.; Ming, J.; Zeng, K.F. Molecular basis of postharvest granulation in orange fruit revealed by metabolite, transcriptome and methylome profiling. Postharvest Biol. Technol. 2020, 166, 111205. [Google Scholar] [CrossRef]
- Ge, Y.; Dong, X.S.; Liu, Y.Z.; Yang, Y.; Zhan, R.L. Molecular and biochemical analyses of avocado (Persea americana) reveal differences in the oil accumulation pattern between the mesocarp and seed during the fruit developmental period. Sci. Hortic. 2021, 276, 109717. [Google Scholar] [CrossRef]
- Bordoloi, K.S.; Baruah, P.M.; Das, M.; Agarwala, N. Unravelling lncRNA mediated gene expression as potential mechanism for regulating secondary metabolism in Citrus limon. Food Biosci. 2022, 46, 101448. [Google Scholar] [CrossRef]
- Dey, S.S.; Sharma, P.K.; Munshi, A.D.; Jaiswal, S.; Behera, T.K.; Kumari, K.; Boopalakrishnan, G.; Iquebal, M.A.; Bhattacharya, R.C.; Rai, A.; et al. Genome wide identification of lncRNAs and circRNAs having regulatory role in fruit shelf life in health crop cucumber (Cucumis sativus L.). Front. Plant Sci. 2022, 13, 884476. [Google Scholar] [CrossRef]
- Li, S.G.; Zhang, J.Y.; Zhang, L.Q.; Fang, X.P.; Luo, J.; An, H.S.; Zhang, X.Y. Genome-wide identification and comprehensive analysis reveal potential roles of long non-coding RNAs in fruit development of southern highbush blueberry (Vaccinium corymbosum L.). Front. Plant Sci. 2022, 13, 1078085. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Zhao, Y.J.; Wu, Y.Q.; Zhao, X.Q.; Hao, Z.X.; Luo, H.; Yuan, Z.H. Transcriptional profiling of long non-coding RNAs regulating fruit cracking in Punica granatum L. under bagging. Front. Plant Sci. 2022, 13, 943547. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, X.Q.; Wang, H.J.; Ye, M.; Liu, Y.T.; Song, Z.H.; Du, T.T.; Cao, H.Y.; Song, L.Q.; Xiao, X.; et al. Identification and analysis of long non-coding RNAs related to UV-B-induced anthocyanin biosynthesis during blood-fleshed peach (Prunus persica) ripening. Front Genet. 2022, 13, 932207. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Saha, B.; Jaiswal, S.; Angadi, U.B.; Rai, A.; Iquebal, M.A. Genome-wide identification and characterization of tissue-specific non-coding RNAs in black pepper (Piper nigrum L.). Front. Plant Sci. 2023, 14, 1079221. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, C.Y.; Cheng, W.Q.; Wu, K.Y.; Gong, B.C. Full-length transcriptome profiling for fruit development in Diospyros oleifera using nanopore sequencing. BMC Genom. Data 2023, 24, 17. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Wen, Y.J.; Liu, J.G.; Jiang, Y.H.; Shi, F.C.; Chen, J.Z.; Cai, C.H.; Ou, L.X.; Yan, Q.; Liu, H.L. The comprehensive detection of mRNAs, lncRNAs, circRNAs, and miRNAs in lychee response to lychee downy blight. Agronomy 2023, 13, 1904. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Wang, X.P.; Qi, K.J.; Bao, J.P.; Zhang, S.L.; Gu, C. Involvement of long non-coding RNAs in pear fruit senescence under high- and low-temperature conditions. Hortic. Plant J. 2023, 9, 224–236. [Google Scholar] [CrossRef]
- Yu, T.T.; Tzeng, D.T.W.; Li, R.; Chen, J.Y.; Zhong, S.L.; Fu, D.Q.; Zhu, B.Z.; Luo, Y.B.; Zhu, H.L. Genome-wide identification of long non-coding RNA targets of the tomato MADS box transcription factor RIN and function analysis. Ann. Bot. 2019, 123, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.Y.; Yang, T.; Li, Y.; Zhang, J.; Wu, T.; Song, T.T.; Yao, Y.C.; Tian, J. The long noncoding RNA MdLNC499 bridges MdWRKY1 and MdERF109 function to regulate early-stage light-induced anthocyanin accumulation in apple fruit. Plant Cell 2021, 33, 3309–3330. [Google Scholar] [CrossRef]
- Tang, Y.J.; Qu, Z.P.; Lei, J.J.; He, R.Q.; Adelson, D.L.; Zhu, Y.L.; Yang, Z.B.; Wang, D. The long noncoding RNA FRILAIR regulates strawberry fruit ripening by functioning as a noncanonical target mimic. PLoS Genet. 2021, 17, e1009461. [Google Scholar] [CrossRef]
- Vignati, E.; Lipska, M.; Dunwell, J.M.; Caccamo, M.; Simkin, A.J. Fruit development in sweet cherry. Plants 2022, 11, 1531. [Google Scholar] [CrossRef] [PubMed]
- Leng, P.; Yuan, B.; Guo, Y.D. The role of abscisic acid in fruit ripening and responses to abiotic stress. J. Exp. Bot. 2014, 65, 4577–4588. [Google Scholar] [CrossRef]
- Peng, Z.Z.; Liu, G.S.; Li, H.L.; Wang, Y.X.; Gao, H.Y.; Jemric, T.; Fu, D.Q. Molecular and genetic events determining the softening of fleshy fruits: A comprehensive review. Int. J. Mol. Sci. 2022, 23, 12482. [Google Scholar] [CrossRef]
- Pelloux, J.; Rusterucci, C.; Mellerowicz, E.J. New insights into pectin methylesterase structure and function. Trends Plant Sci. 2007, 12, 267–277. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruit. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.M.; Wang, H.; Li, M.F.; Wang, J.; Yang, Y.; Zhang, X.M.; Yan, G.H.; Zhang, H.; Liu, J.S.; Zhang, K.C. The R2R3 MYB transcription factor PavMYB10.1 involves in anthocyanin biosynthesis and determines fruit skin colour in sweet cherry (Prunus avium L.). Plant Biotechnol. J. 2016, 14, 2120–2133. [Google Scholar] [CrossRef]
- Starkevic, P.; Paukstyte, J.; Kazanaviciute, V.; Denkovskiene, E.; Stanys, V.; Bendokas, V.; Siksnianas, T.; Razanskiene, A.; Razanskas, R. Expression and anthocyanin biosynthesis-modulating potential of sweet cherry (Prunus avium L.) MYB10 and bHLH genes. PLoS ONE 2015, 10, e0126991. [Google Scholar] [CrossRef]
- Zhai, Z.F.; Xiao, Y.Q.; Wang, Y.Y.; Sun, Y.T.; Peng, X.; Feng, C.; Zhang, X.; Du, B.Y.; Zhou, X.; Wang, C.; et al. Abscisic acid-responsive transcription factors PavDof2/6/15 mediate fruit softening in sweet cherry. Plant Physiol. 2022, 190, 2501–2518. [Google Scholar] [CrossRef] [PubMed]
- Tucker, G.; Yin, X.R.; Zhang, A.D.; Wang, M.M.; Zhu, Q.G.; Liu, X.F.; Xie, X.L.; Chen, K.S.; Grierson, D. Ethylene and fruit softening. Food Qual. Saf. 2017, 1, 253–267. [Google Scholar] [CrossRef]
- Rathinam, M.; Rao, U.; Sreevathsa, R. Novel biotechnological strategies to combat biotic stresses: Polygalacturonase inhibitor (PGIP) proteins as a promising comprehensive option. Appl. Microbiol. Biotechnol. 2020, 104, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Tian, C.P.; Li, X.W.; Gong, H.S.; Zhang, A.D. Transcriptome co-expression network analysis identifies key genes and regulators of sweet cherry anthocyanin biosynthesis. Horticulturae 2021, 7, 123. [Google Scholar] [CrossRef]
- Chen, C.Q.; Chen, H.X.; Yang, W.L.; Li, J.; Tang, W.J.; Gong, R.G. Transcriptomic and metabolomic analysis of quality changes during sweet cherry fruit development and mining of related genes. Int. J. Mol. Sci. 2022, 23, 7402. [Google Scholar] [CrossRef]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Lv, J.; Cui, W.; Liu, H.; He, H.J.; Xiu, Y.C.; Guo, J.; Liu, H.; Liu, Q.; Zeng, T.B.; Chen, Y.; et al. Identification and characterization of long non-coding RNAs related to mouse embryonic brain development from available transcriptomic data. PLoS ONE 2013, 8, e71152. [Google Scholar] [CrossRef]
- Kelley, D.; Rinn, J. Transposable elements reveal a stem cell-specific class of long noncoding RNAs. Genome Biol. 2012, 13, R107. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.G.; Yu, K.T.; Zhang, C.H.; Liu, Y.N.; Chen, R.S.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.Q.; Kocher, J.P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2013, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integrat. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Clean Reads (million) | Mapped Reads (million) | Mapping Rate |
|---|---|---|---|
| Y1 | 113.55 | 90.09 | 79.34% |
| Y2 | 114.69 | 91.6 | 79.87% |
| Y3 | 112.88 | 96.36 | 78.42% |
| P1 | 133.52 | 106.04 | 79.42% |
| P2 | 117.96 | 96.62 | 81.91% |
| P3 | 114.27 | 94.32 | 82.54% |
| DR1 | 130.05 | 107.59 | 82.72% |
| DR2 | 114.4 | 94.67 | 82.75% |
| DR3 | 109.79 | 90.26 | 82.21% |
| Gene ID | Log2 (Fold Change) | Annotation | Upstream DE-lncRNAs |
|---|---|---|---|
| Pav_sc0001440.1_g080.1.mk | −2.08 | CYP707A1-like | MSTRG.3112.1 (−1.85), MSTRG.37034.2 (−1.86) |
| Pav_sc0000852.1_g900.1.mk | −2.50 | CYP707A4-like1 | MSTRG.10166.2 (2.03), MSTRG.11213.10 (2.24), MSTRG.26521.2 (−2.33), MSTRG.3040.1 (−1.55), MSTRG.30960.1 (−3.27), MSTRG.34331.1 (−3.22), MSTRG.36057.7 (−3.69), MSTRG.36335.1 (−2.14), MSTRG.40513.1 (−2.24), MSTRG.8543.2 (−1.07) |
| Pav_sc0000071.1_g720.1.mk | −4.26 | CYP707A4-like2 | MSTRG.11213.10 (2.24), MSTRG.26521.2 (−2.33), MSTRG.3040.1 (−1.55), MSTRG.30960.1 (−3.27), MSTRG.34331.1 (−3.22), MSTRG.36335.1 (−2.14), MSTRG.8543.2 (−1.07) |
| Pav_sc0000020.1_g060.1.mk | 1.17 | PG QRT3 | MSTRG.12848.1 (2.08), MSTRG.37034.2 (−1.86) |
| Pav_sc0000212.1_g1900.1.mk | 1.23 | TBG | MSTRG.30065.2 (7.15), MSTRG.3112.1 (−1.85), MSTRG.35294.2 (4.50), MSTRG.38400.3 (3.10), MSTRG.9626.1 (−3.69) |
| Pav_sc0000028.1_g320.1.mk | −1.99 | PME inhibitor1 | MSTRG.3112.1 (−1.85), MSTRG.37034.2 (−1.86) |
| Pav_sc0004475.1_g020.1.br | −1.10 | PME inhibitor22 | MSTRG.18052.1 (−1.40), MSTRG.2169.2 (−1.59), MSTRG.25930.8 (−3.20), MSTRG.2981.10 (−1.62), MSTRG.30065.2 (7.15), MSTRG.33128.1 (−1.77), MSTRG.33241.2 (−1.18), MSTRG.35294.2 (4.50), MSTRG.37117.4 (5.00), MSTRG.38400.3 (3.10), MSTRG.40513.1 (−2.24), MSTRG.41450.1 (−2.38) |
| Pav_sc0001335.1_g240.1.mk | −1.38 | PME inhibitor34 | MSTRG.10166.2 (2.03), MSTRG.11213.10 (2.24), MSTRG.26521.2 (−2.33), MSTRG.2981.4 (−1.01), MSTRG.29836.1 (4.25), MSTRG.30960.1 (−3.27), MSTRG.33370.3 (−1.1), MSTRG.34331.1 (−3.22), MSTRG.8543.2 (−1.07) |
| Pav_sc0000045.1_g280.1.mk | 1.96 | CHS1-like | |
| Pav_sc0005746.1_g030.1.mk | 1.83 | CHI3 | MSTRG.12848.1 (2.08), MSTRG.29836.1 (4.25), MSTRG.8543.2 (−1.07) |
| Pav_sc0002208.1_g840.1.mk | 2.16 | DFR | MSTRG.12848.1 (2.08), MSTRG.29836.1 (4.25) |
| Pav_sc0000464.1_g130.1.br | 1.97 | MYB10.1 | MSTRG.26521.2 (−2.33), MSTRG.29836.1 (4.25), MSTRG.8543.2 (−1.07) |
| Pav_sc0000102.1_g430.1.mk | −1.39 | bHLH33 | MSTRG.10166.2 (2.03), MSTRG.11213.10 (2.24), MSTRG.13193.1 (1.06), MSTRG.26521.2 (−2.33), MSTRG.3040.1 (−1.55), MSTRG.30960.1 (−3.27), MSTRG.33370.3 (−1.10), MSTRG.8543.2 (−1.07) |
| Pav_sc0001900.1_g150.1.mk | −1.34 | ARF8 | MSTRG.10166.2 (2.03), MSTRG.11213.10 (2.24), MSTRG.26521.2 (−2.33), MSTRG.2981.4 (−1.01), MSTRG.29836.1 (4.25), MSTRG.3040.1 (−1.55), MSTRG.30960.1 (−3.27), MSTRG.33370.3 (−1.10), MSTRG.34331.1 (−3.22), MSTRG.36057.7 (−3.69), MSTRG.8543.2 (−1.07) |
| Pav_sc0000358.1_g850.1.mk | −1.39 | Dof15 | MSTRG.18052.1 (−1.40), MSTRG.2169.2 (−1.59), MSTRG.25930.8 (−3.20), MSTRG.2642.1 (5.43), MSTRG.2981.10 (−1.62), MSTRG.30065.2 (7.15), MSTRG.33128.1 (−1.77), MSTRG.33241.2 (−1.18), MSTRG.35294.2 (4.50), MSTRG.37117.4 (5.00), MSTRG.38400.3 (3.10), MSTRG.40513.1 (−2.24) |
| Gene ID | Log2 (Fold Change) | Annotation | Upstream DE-lncRNAs |
|---|---|---|---|
| Pav_sc0000020.1_g060.1.mk | 2.39 | PG QRT3 | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.165.3 (1.84), MSTRG.21139.1 (3.20), MSTRG.22209.11 (−4.63), MSTRG.23122.2 (1.24), MSTRG.23545.5 (2.47), MSTRG.29844.2 (1.97), MSTRG.30707.15 (−5.26), MSTRG.31012.1 (2.79), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.37034.2 (3.80), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.4832.2 (4.35), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0003823.1_g150.1.mk | 1.09 | PG-like | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.338.1 (3.11), MSTRG.41437.1 (1.45), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0000428.1_g510.1.mk | 3.29 | XTH2-like1 | MSTRG.15876.1 (−1.69), MSTRG.1606.1 (1.57), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.34175.1 (1.78), MSTRG.35257.14 (6.49), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.38366.1 (3.16), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.4215.1 (1.35), MSTRG.4832.2 (4.35), MSTRG.5140.1 (8.41) |
| Pav_sc0000428.1_g520.1.mk | 2.49 | XTH2-like2 | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.1606.1 (1.57), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.29844.2 (1.97), MSTRG.30536.1 (3.65), MSTRG.30707.15 (−5.26), MSTRG.31012.1 (2.79), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.34175.1 (1.78), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.38366.1 (3.16), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.4215.1 (1.35), MSTRG.4832.2 (4.35), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0000428.1_g530.1.mk | 3.61 | XTH2-like3 | MSTRG.15876.1 (−1.69), MSTRG.1606.1 (1.57), MSTRG.21139.1 (3.20), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.34175.1 (1.78), MSTRG.35257.14 (6.49), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.38366.1 (3.16), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.4215.1 (1.35), MSTRG.4832.2 (4.35), MSTRG.5140.1 (8.41) |
| Pav_sc0000910.1_g790.1.mk | 1.15 | XTH8 | MSTRG.33128.1 (1.17), MSTRG.40513.1 (1.42), MSTRG.41450.1 (1.71) |
| Pav_sc0000308.1_g610.1.mk | 1.70 | XTH23 | MSTRG.25299.1 (3.76), MSTRG.30248.1 (4.10), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35) |
| Pav_sc0000359.1_g040.1.mk | 3.67 | XTH24 | MSTRG.13549.1 (1.91), MSTRG.25299.1 (3.76), MSTRG.30065.2 (−7.24), MSTRG.30248.1 (4.10), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.35294.2 (−4.54), MSTRG.4215.1 (1.35) |
| Pav_sc0000354.1_g310.1.mk | 1.32 | XTH33 | MSTRG.2169.2 (1.13), MSTRG.30065.2 (−7.24), MSTRG.35294.2 (−4.54), MSTRG.38400.3 (−2.61) |
| Pav_sc0000582.1_g630.1.mk | 2.06 | CEL9-like | MSTRG.13879.1 (1.09), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.34175.1 (1.78), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35) |
| Pav_sc0000652.1_g760.1.mk | 1.83 | CEL CX | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.165.3 (1.84), MSTRG.21139.1 (3.20), MSTRG.22209.11 (−4.63), MSTRG.23122.2 (1.24), MSTRG.23545.5 (2.47), MSTRG.29844.2 (1.97), MSTRG.30536.1 (3.65), MSTRG.30707.15 (−5.26), MSTRG.31012.1 (2.79), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0000244.1_g040.1.mk | 1.12 | EXP-like A2.1 | MSTRG.13549.1 (1.91), MSTRG.22060.23 (2.06), MSTRG.25299.1 (3.76), MSTRG.25638.2 (5.61), MSTRG.30248.1 (4.10), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35) |
| Pav_co4016743.1_g010.1.br | 1.32 | EXP-like A2.2 | MSTRG.22060.23 (2.06), MSTRG.25299.1 (3.76), MSTRG.25638.2 (5.61), MSTRG.30248.1 (4.10), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35) |
| Pav_sc0000582.1_g890.1.mk | −1.76 | PG inhibitor 1-like | MSTRG.13879.1 (1.09), MSTRG.18349.5 (−1.23), MSTRG.30248.1 (4.10), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.338.1 (3.11), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35), MSTRG.5140.1 (8.41) |
| Pav_sc0000843.1_g400.1.mk | −2.52 | PME inhibitor2 | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.21139.1 (3.20), MSTRG.22209.11 (−4.63), MSTRG.23122.2 (1.24), MSTRG.29844.2 (1.97), MSTRG.30707.15 (−5.26), MSTRG.31012.1 (2.79), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0001335.1_g240.1.mk | −1.15 | PME inhibitor34 | |
| Pav_sc0000045.1_g280.1.mk | 2.14 | CHS1-like | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.29844.2 (1.97), MSTRG.30048.2 (3.09), MSTRG.338.1 (3.11), MSTRG.41437.1 (1.45), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0007510.1_g020.1.mk | 1.33 | CHI | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.165.3 (1.84), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.29844.2 (1.97), MSTRG.31012.1 (2.79), MSTRG.338.1 (3.11), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0002208.1_g840.1.mk | 1.26 | DFR | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.29844.2 (1.97), MSTRG.338.1 (3.11), MSTRG.41437.1 (1.45), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0000625.1_g100.1.mk | 2.74 | MYB44-like | MSTRG.15876.1 (−1.69), MSTRG.1606.1 (1.57), MSTRG.21139.1 (3.20), MSTRG.22209.11 (−4.63), MSTRG.23122.2 (1.24), MSTRG.23545.5 (2.47), MSTRG.29844.2 (1.97), MSTRG.30707.15 (−5.26), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.34175.1 (1.78), MSTRG.35257.14 (6.49), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.38366.1 (3.16), MSTRG.40130.1 (2.53), MSTRG.41437.1 (1.45), MSTRG.4215.1 (1.35), MSTRG.4832.2 (4.35), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0000766.1_g080.1.mk | 1.49 | MYB306-like1 | MSTRG.2169.2 (1.13), MSTRG.30065.2 (−7.24), MSTRG.33128.1 (1.17), MSTRG.35294.2 (−4.54), MSTRG.38400.3 (−2.61), MSTRG.40513.1 (1.42), MSTRG.41450.1 (1.71), MSTRG.41822.2 (1.17) |
| Pav_sc0000877.1_g610.1.mk | 1.59 | MYB306-like2 | MSTRG.23059.1 (−3.96), MSTRG.38366.1 (3.16) |
| Pav_sc0000464.1_g210.1.br | 1.11 | MYB10 V1-3 | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.29844.2 (1.97), MSTRG.31012.1 (2.79), MSTRG.338.1 (3.11), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30), |
| Pav_sc0000586.1_g190.1.mk | 1.87 | bHLH13 | MSTRG.23059.1 (−3.96) |
| Pav_sc0000624.1_g1350.1.mk | 2.66 | bHLH35 | MSTRG.2169.2 (1.13), MSTRG.30065.2 (−7.24), MSTRG.33128.1 (1.17), MSTRG.35294.2 (−4.54), MSTRG.38400.3 (−2.61) |
| Pav_sc0000143.1_g320.1.mk | −1.48 | bHLH51 | MSTRG.13122.1 (1.26), MSTRG.15876.1 (−1.69), MSTRG.21139.1 (3.20), MSTRG.30707.15 (−5.26), MSTRG.338.1 (3.11), MSTRG.41437.1 (1.45), MSTRG.4882.1 (−1.47), MSTRG.5140.1 (8.41), MSTRG.7786.1 (2.30) |
| Pav_sc0001519.1_g020.1.mk | 1.29 | bHLH74 | MSTRG.13879.1 (1.09), MSTRG.23545.5 (2.47), MSTRG.3112.1 (2.62), MSTRG.33336.1 (1.35), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.4215.1 (1.35) |
| Pav_sc0000998.1_g640.1.mk | 3.00 | bHLH92 | MSTRG.40513.1 (1.42) |
| Pav_sc0001422.1_g020.1.mk | 1.02 | bHLH113 | MSTRG.15876.1 (−1.69), MSTRG.1606.1 (1.57), MSTRG.21139.1 (3.20), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.338.1 (3.11), MSTRG.35257.14 (6.49), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.38366.1 (3.16), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35), MSTRG.5140.1 (8.41) |
| Pav_sc0001313.1_g380.1.mk | 3.07 | bHLH162-like1 | MSTRG.30065.2 (−7.24), MSTRG.35294.2 (−4.54), MSTRG.38400.3 (−2.61), MSTRG.9626.1 (3.95) |
| Pav_sc0001309.1_g720.1.mk | 3.41 | bHLH162-like2 | MSTRG.13549.1 (1.91), MSTRG.25299.1 (3.76), MSTRG.27214.15 (−2.50), MSTRG.30065.2 (−7.24), MSTRG.3112.1 (2.62), MSTRG.35294.2 (−4.54) |
| Pav_sc0006499.1_g050.1.mk | 2.15 | MYC2 | MSTRG.13879.1 (1.09), MSTRG.1606.1 (1.57), MSTRG.21139.1 (3.20), MSTRG.23122.2 (1.24), MSTRG.23545.5 (2.47), MSTRG.29844.2 (1.97), MSTRG.30707.15 (−5.26), MSTRG.3112.1 (2.62), MSTRG.338.1 (3.11), MSTRG.33948.1 (1.57), MSTRG.34175.1 (1.78), MSTRG.37034.2 (3.80), MSTRG.37910.1 (2.72), MSTRG.38366.1 (3.16), MSTRG.40130.1 (2.53), MSTRG.4215.1 (1.35), MSTRG.4832.2 (4.35), MSTRG.5140.1 (8.41) |
| Pav_sc0001557.1_g070.1.mk | 3.08 | WD40.1 | MSTRG.25299.1 (3.76), MSTRG.30065.2 (−7.24), MSTRG.3112.1 (2.62), MSTRG.35294.2 (−4.54), MSTRG.38400.3 (−2.61) |
| Pav_sc0001557.1_g120.1.br | 3.05 | WD40.2 | |
| Pav_sc0001963.1_g370.1.mk | 1.42 | WD40.3 | MSTRG.30065.2 (−7.24), MSTRG.35294.2 (−4.54), MSTRG.38400.3 (−2.61) |
| Cyanidin 3-Rutinoside | Cyanin Chloride | Cyanidin 3-O-Galactoside | Peonidin 3-O-Glucoside Chloride | Cyanidin 3-O-Glucoside | Cyanidin O-Syringic Acid | |
|---|---|---|---|---|---|---|
| CYP707A1-like | 0.667 * | 0.788 * | 0.582 | 0.726 * | 0.591 * | 0.506 |
| CYP707A4-like1 | −0.813 * | −0.691 * | −0.858 * | −0.630 * | −0.850 * | −0.882 * |
| CYP707A4-like2 | −0.849 * | −0.742 * | −0.893 * | −0.692 * | −0.887 * | −0.915 * |
| CHS1-like | 0.894 * | 0.965 * | 0.848 * | 0.968 * | 0.874 * | 0.839 * |
| CHI | 0.895 * | 0.913 * | 0.836 * | 0.847 * | 0.845 * | 0.813 * |
| CHI3 | 0.981 * | 0.956 * | 0.973 * | 0.902 * | 0.986 * | 0.977 * |
| DFR | 0.964 * | 0.980 * | 0.930 * | 0.937 * | 0.945 * | 0.921 * |
| MYB10.1 | 0.971 * | 0.953 * | 0.967 * | 0.932 * | 0.980 * | 0.979 * |
| MYB44-like | 0.818 * | 0.910 * | 0.751 * | 0.860 * | 0.762 * | 0.695 * |
| MYB306-like1 | −0.292 | −0.105 | −0.392 | −0.077 | −0.374 | −0.451 |
| MYB306-like2 | 0.498 | 0.577 | 0.420 | 0.487 | 0.401 | 0.321 |
| MYB10 V1-3 | 0.967 * | 0.962 * | 0.927 * | 0.875 * | 0.937 * | 0.907 * |
| bHLH13 | 0.118 | 0.255 | 0.031 | 0.252 | 0.050 | −0.020 |
| bHLH33 | −0.890 * | −0.799 * | −0.921 * | −0.754 * | −0.925 * | −0.952 * |
| bHLH35 | −0.238 | −0.058 | −0.333 | −0.066 | −0.327 | −0.418 |
| bHLH51 | −0.833 * | −0.903 * | −0.759 * | −0.848 * | −0.787 * | −0.731 * |
| bHLH74 | 0.540 | 0.684 * | 0.435 | 0.629 * | 0.465 | 0.376 |
| bHLH92 | −0.788 * | −0.657 * | −0.849 * | −0.613 * | −0.842 * | −0.886 * |
| bHLH113 | 0.696 * | 0.829 * | 0.622 * | 0.801 * | 0.648 * | 0.571 |
| bHLH162-like1 | −0.001 | 0.179 | −0.115 | 0.160 | −0.103 | −0.197 |
| bHLH162-like2 | 0.200 | 0.392 | 0.096 | 0.412 | 0.124 | 0.039 |
| MYC2 | 0.696 * | 0.810 * | 0.609 * | 0.756 * | 0.620 * | 0.542 |
| WD40.1 | 0.180 | 0.365 | 0.064 | 0.366 | 0.091 | 0.004 |
| WD40.2 | 0.242 | 0.360 | 0.131 | 0.251 | 0.131 | 0.036 |
| WD40.3 | −0.083 | 0.098 | −0.197 | 0.088 | −0.178 | −0.267 |
| MSTRG.11213.10 | 0.780 * | 0.650 * | 0.831 * | 0.606 * | 0.829 * | 0.874 * |
| MSTRG.13193.1 | 0.687 * | 0.547 | 0.738 * | 0.496 | 0.744 * | 0.797 * |
| MSTRG.15876.1 | −0.856 * | −0.930 * | −0.798 * | −0.899 * | −0.815 * | −0.765 * |
| MSTRG.1606.1 | 0.637 * | 0.757 * | 0.563 | 0.714 * | 0.582 * | 0.505 |
| MSTRG.165.3 | 0.777 * | 0.815 * | 0.704 * | 0.762 * | 0.711 * | 0.674 * |
| MSTRG.21139.1 | 0.813 * | 0.899 * | 0.738 * | 0.865 * | 0.761 * | 0.707 * |
| MSTRG.22209.11 | −0.837 * | −0.890 * | −0.788 * | −0.846 * | −0.795 * | −0.751 * |
| MSTRG.23122.2 | 0.842 * | 0.893 * | 0.785 * | 0.841 * | 0.788 * | 0.741 * |
| MSTRG.23545.5 | 0.709 * | 0.782 * | 0.609 * | 0.675 * | 0.620 * | 0.544 |
| MSTRG.25299.1 | 0.313 | 0.490 | 0.224 | 0.527 | 0.260 | 0.188 |
| MSTRG.26521.2 | −0.859 * | −0.769 * | −0.897 * | −0.742 * | −0.905 * | −0.937 * |
| MSTRG.27214.15 | −0.233 | −0.357 | −0.134 | −0.296 | −0.180 | −0.096 |
| MSTRG.29836.1 | 0.949 * | 0.900 * | 0.921 * | 0.791 * | 0.927 * | 0.911 * |
| MSTRG.29844.2 | 0.829 * | 0.886 * | 0.747 * | 0.791 * | 0.761 * | 0.699 * |
| MSTRG.30048.2 | 0.716 * | 0.845 * | 0.689 * | 0.929 * | 0.728 * | 0.693 * |
| MSTRG.30065.2 | 0.010 | −0.182 | 0.129 | −0.174 | 0.102 | 0.196 |
| MSTRG.3040.1 | −0.809 * | −0.678 * | −0.857 * | −0.581 | −0.844 * | −0.867 * |
| MSTRG.30707.15 | −0.723 * | −0.820 * | −0.644 * | −0.769 * | −0.661 * | −0.596 * |
| MSTRG.30960.1 | −0.828 * | −0.712 * | −0.877 * | −0.672 * | −0.874 * | −0.913 * |
| MSTRG.31012.1 | 0.839 * | 0.860 * | 0.758 * | 0.717 * | 0.755 * | 0.690 * |
| MSTRG.3112.1 | 0.454 | 0.636 * | 0.363 | 0.671 * | 0.397 | 0.320 |
| MSTRG.33128.1 | −0.503 | −0.323 | −0.587 * | −0.259 | −0.570 | −0.630 * |
| MSTRG.33336.1 | 0.502 | 0.651 * | 0.388 | 0.600 * | 0.425 | 0.334 |
| MSTRG.33370.3 | −0.891 * | −0.814 * | −0.924 * | −0.732 * | −0.924 * | −0.926 * |
| MSTRG.338.1 | 0.867 * | 0.946 * | 0.805 * | 0.906 * | 0.821 * | 0.765 * |
| MSTRG.33948.1 | 0.856 * | 0.902 * | 0.790 * | 0.787 * | 0.793 * | 0.724 * |
| MSTRG.34175.1 | 0.658 * | 0.731 * | 0.578 | 0.597 * | 0.562 | 0.470 |
| MSTRG.34331.1 | −0.841 * | −0.738 * | −0.881 * | −0.690 * | −0.874 * | −0.901 * |
| MSTRG.35257.14 | 0.714 * | 0.825 * | 0.682 * | 0.825 * | 0.694 * | 0.631 * |
| MSTRG.35294.2 | 0.009 | −0.178 | 0.118 | −0.185 | 0.101 | 0.187 |
| MSTRG.36057.7 | −0.707 * | −0.569 | −0.769 * | −0.537 | −0.767 * | −0.821 * |
| MSTRG.36335.1 | −0.760 * | −0.663 * | −0.797 * | −0.646 * | −0.791 * | −0.828 * |
| MSTRG.37034.2 | 0.729 * | 0.820 * | 0.633 * | 0.741 * | 0.646 * | 0.574 |
| MSTRG.37910.1 | 0.613 * | 0.758 * | 0.526 | 0.758 * | 0.555 | 0.487 |
| MSTRG.38366.1 | 0.702 * | 0.782 * | 0.624 * | 0.668 * | 0.621 * | 0.534 |
| MSTRG.38400.3 | 0.045 | −0.133 | 0.166 | −0.115 | 0.151 | 0.243 |
| MSTRG.40130.1 | 0.721 * | 0.829 * | 0.638 * | 0.771 * | 0.649 * | 0.573 |
| MSTRG.40513.1 | −0.563 | −0.384 | −0.646 * | −0.314 | −0.626 * | −0.683 * |
| MSTRG.41437.1 | 0.822 * | 0.909 * | 0.761 * | 0.895 * | 0.786 * | 0.736 * |
| MSTRG.41450.1 | −0.462 | −0.308 | −0.539 | −0.285 | −0.531 | −0.593 * |
| MSTRG.41822.2 | −0.198 | 0.009 | −0.301 | 0.106 | −0.254 | −0.312 |
| MSTRG.4215.1 | 0.587 * | 0.734 * | 0.485 | 0.701 * | 0.514 | 0.429 |
| MSTRG.4832.2 | 0.776 * | 0.828 * | 0.705 * | 0.691 * | 0.695 * | 0.613 * |
| MSTRG.4882.1 | −0.801 * | −0.863 * | −0.740 * | −0.808 * | −0.767 * | −0.716 * |
| MSTRG.5140.1 | 0.855 * | 0.936 * | 0.786 * | 0.892 * | 0.804 * | 0.747 * |
| MSTRG.7786.1 | 0.872 * | 0.930 * | 0.804 * | 0.889 * | 0.831 * | 0.789 * |
| MSTRG.8543.2 | −0.898 * | −0.817 * | −0.921 * | −0.777 * | −0.930 * | −0.956 * |
| MSTRG.9626.1 | 0.090 | 0.241 | −0.032 | 0.192 | −0.021 | −0.110 |
| MSTRG.10166.2 | 0.826 * | 0.713 * | 0.862 * | 0.661 * | 0.863 * | 0.899 * |
| MSTRG.12848.1 | 0.959 * | 0.934 * | 0.932 * | 0.806 * | 0.930 * | 0.893 * |
| MSTRG.13122.1 | 0.819 * | 0.884 * | 0.744 * | 0.842 * | 0.771 * | 0.726 * |
| MSTRG.13549.1 | 0.343 | 0.505 | 0.269 | 0.566 | 0.301 | 0.246 |
| MSTRG.13879.1 | 0.476 | 0.602 * | 0.350 | 0.499 | 0.376 | 0.279 |
| MSTRG.2169.2 | −0.392 | −0.202 | −0.463 | −0.140 | −0.451 | −0.522 |
| MSTRG.23059.1 | −0.431 | −0.532 | −0.360 | −0.521 | −0.368 | −0.309 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Fu, D.; Duan, X.; Zhou, J.; Chang, H.; Xu, R.; Wang, B.; Wang, Y. Integrated Metabolome, Transcriptome and Long Non-Coding RNA Analysis Reveals Potential Molecular Mechanisms of Sweet Cherry Fruit Ripening. Int. J. Mol. Sci. 2024, 25, 9860. https://doi.org/10.3390/ijms25189860
Liu G, Fu D, Duan X, Zhou J, Chang H, Xu R, Wang B, Wang Y. Integrated Metabolome, Transcriptome and Long Non-Coding RNA Analysis Reveals Potential Molecular Mechanisms of Sweet Cherry Fruit Ripening. International Journal of Molecular Sciences. 2024; 25(18):9860. https://doi.org/10.3390/ijms25189860
Chicago/Turabian StyleLiu, Gangshuai, Daqi Fu, Xuwei Duan, Jiahua Zhou, Hong Chang, Ranran Xu, Baogang Wang, and Yunxiang Wang. 2024. "Integrated Metabolome, Transcriptome and Long Non-Coding RNA Analysis Reveals Potential Molecular Mechanisms of Sweet Cherry Fruit Ripening" International Journal of Molecular Sciences 25, no. 18: 9860. https://doi.org/10.3390/ijms25189860
APA StyleLiu, G., Fu, D., Duan, X., Zhou, J., Chang, H., Xu, R., Wang, B., & Wang, Y. (2024). Integrated Metabolome, Transcriptome and Long Non-Coding RNA Analysis Reveals Potential Molecular Mechanisms of Sweet Cherry Fruit Ripening. International Journal of Molecular Sciences, 25(18), 9860. https://doi.org/10.3390/ijms25189860