
DNA 5mC and RNA m6A Collaborate to Upregulate Phosphoenolpyruvate Carboxykinase 2 for Kupffer Cell Activation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Metabolic Reprogramming in LPS-Activated KCs Is Associated with PCK2 Upregulation
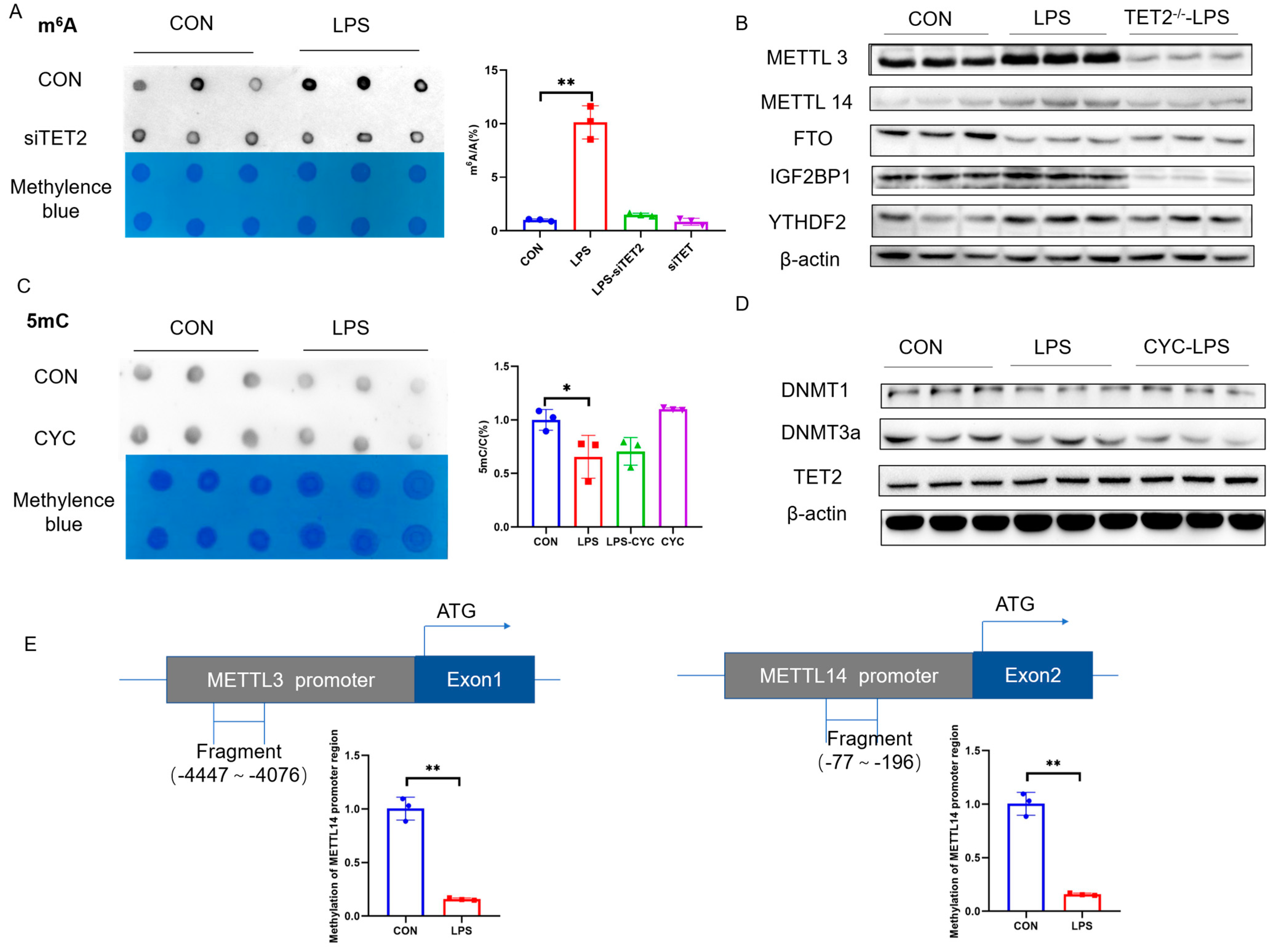
2.2. PCK2 Upregulation in LPS-Activated KCs Is Associated with Global DNA 5mC Demethylation and RNA m6A Hypermethylation
2.3. Suppression of 5mC Demethylation and m6A Hypermethylation Rectifies LPS-Induced PCK2 Upregulation
2.4. CpG Island of PCK2 Promoter Is 5mC-Demethylated and 3′UTR of PCK2 mRNA Is m6A-Hypermethylated in LPS-Activated KCs
2.5. 5mC Demethylation and m6A Hypermethylation Contribute Collectively to PCK2 Upregulation in LPS-Activated KCs
2.6. METTL3/METTL14 Genes Are Transactivated in LPS-Activated KCs via Promoter 5mC Hypomethylation
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cytokine Quantification
4.3. RNA Isolation and Real-Time qPCR
4.4. Protein Extraction and Western Blot Assay
4.5. DNA 5mC and RNA m6A Dot Blot Assay
4.6. Methylated DNA Immunoprecipitation (MeDIP) Assay
4.7. SELECT for Site-Specific Detection of m6A
4.8. siRNA Transfection and Inhibitor Treatment
4.9. RNA Decay Assay
4.10. Dual-Luciferase Reporter Assay
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shu, B.; Zhou, Y.X.; Li, H.; Zhang, R.Z.; He, C.; Yang, X. The METTL3/MALAT1/PTBP1/USP8/TAK1 axis promotes pyroptosis and M1 polarization of macrophages and contributes to liver fibrosis. Cell Death Discov. 2021, 7, 368. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yang, L.; Peng, X.; Mao, M.; Liu, X.; Song, J.; Li, H.; Chen, F. METTL3 promotes m6A hypermethylation of RBM14 via YTHDF1 leading to the progression of hepatocellular carcinoma. Hum. Cell 2022, 35, 1838–1855. [Google Scholar] [CrossRef] [PubMed]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.J.; Barnes, M.; Tang, H.; Pritchard, M.T.; Nagy, L.E. Kupffer cells in the liver. Compr. Physiol. 2013, 3, 785–797. [Google Scholar]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Palsson-McDermott, E.M.; Curtis, A.M.; Goel, G.; Lauterbach, M.A.R.; Sheedy, F.J.; Gleeson, L.E.; van den Bosch, M.W.M.; Quinn, S.R.; Domingo-Fernandez, R.; Johnston, D.G.W.; et al. Pyruvate Kinase M2 Regulates Hif-1alpha Activity and IL-1beta Induction and Is a Critical Determinant of the Warburg Effect in LPS-Activated Macrophages. Cell Metab. 2015, 21, 347. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Feng, Y.; Yang, Y.; Hu, Y.; Jia, Y.; Yang, S.; Zhao, N.; Zhao, R. A Novel Function of Mitochondrial Phosphoenolpyruvate Carboxykinase as a Regulator of Inflammatory Response in Kupffer Cells. Front. Cell Dev. Biol. 2021, 9, 726931. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Sun, Y.; Huang, C.; Li, J.; Wang, Y. New advances of DNA/RNA methylation modification in liver fibrosis. Cell. Signal. 2022, 92, 110224. [Google Scholar] [CrossRef]
- Choubey, P.; Kaur, H.; Bansal, K. Modulation of DNA/RNA Methylation Signaling Mediating Metabolic Homeostasis in Cancer. Subcell. Biochem. 2022, 100, 201–237. [Google Scholar]
- Zhang, Q.; Zhao, K.; Shen, Q.; Han, Y.; Gu, Y.; Li, X.; Zhao, D.; Liu, Y.; Wang, C.; Zhang, X.; et al. Tet2 is required to resolve inflammation by recruiting Hdac2 to specifically repress IL-6. Nature 2015, 525, 389–393. [Google Scholar] [CrossRef]
- Feng, Y.; Dong, H.; Sun, B.; Hu, Y.; Yang, Y.; Jia, Y.; Jia, L.; Zhong, X.; Zhao, R. METTL3/METTL14 Transactivation and m6A-Dependent TGF-beta1 Translation in Activated Kupffer Cells. Cell Mol. Gastroenterol. Hepatol. 2021, 12, 839–856. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.C.; Vatner, D.F.; Shulman, G.I. Regulation of hepatic glucose metabolism in health and disease. Nat. Rev. Endocrinol. 2017, 13, 572–587. [Google Scholar] [CrossRef]
- Wang, Z.; Dong, C. Gluconeogenesis in Cancer: Function and Regulation of PEPCK, FBPase, and G6Pase. Trends Cancer 2019, 5, 30–45. [Google Scholar] [CrossRef]
- Ye, Q.; Liu, Y.; Zhang, G.; Deng, H.; Wang, X.; Tuo, L.; Chen, C.; Pan, X.; Wu, K.; Fan, J.; et al. Deficiency of gluconeogenic enzyme PCK1 promotes metabolic-associated fatty liver disease through PI3K/AKT/PDGF axis activation in male mice. Nat. Commun. 2023, 14, 1402. [Google Scholar] [CrossRef]
- Zhou, B.; Magana, L.; Hong, Z.; Huang, L.S.; Chakraborty, S.; Tsukasaki, Y.; Huang, C.; Wang, L.; Di, A.; Ganesh, B.; et al. The angiocrine Rspondin3 instructs interstitial macrophage transition via metabolic-epigenetic reprogramming and resolves inflammatory injury. Nat. Immunol. 2020, 21, 1430–1443. [Google Scholar] [CrossRef]
- Cong, B.; Zhang, Q.; Cao, X. The function and regulation of TET2 in innate immunity and inflammation. Protein Cell 2021, 12, 165–173. [Google Scholar] [CrossRef]
- Ma, S.; Wan, X.; Deng, Z.; Shi, L.; Hao, C.; Zhou, Z.; Zhou, C.; Fang, Y.; Liu, J.; Yang, J.; et al. Epigenetic regulator CXXC5 recruits DNA demethylase Tet2 to regulate TLR7/9-elicited IFN response in pDCs. J. Exp. Med. 2017, 214, 1471–1491. [Google Scholar] [CrossRef]
- Seenappa, V.; Das, B.; Joshi, M.B.; Satyamoorthy, K. Context Dependent Regulation of Human Phosphoenolpyruvate Carboxykinase Isoforms by DNA Promoter Methylation and RNA Stability. J. Cell Biochem. 2016, 117, 2506–2520. [Google Scholar] [CrossRef]
- Meng, J.; Liu, X.; Tang, S.; Liu, Y.; Zhao, C.; Zhou, Q.; Li, N.; Hou, S. METTL3 inhibits inflammation of retinal pigment epithelium cells by regulating NR2F1 in an m6A-dependent manner. Front. Immunol. 2022, 13, 905211. [Google Scholar] [CrossRef]
- Gan, X.; Dai, Z.; Ge, C.; Yin, H.; Wang, Y.; Tan, J.; Sun, S.; Zhou, W.; Yuan, S.; Yang, F. FTO promotes liver inflammation by suppressing m6A mRNA methylation of IL-17RA. Front. Oncol. 2022, 12, 989353. [Google Scholar] [CrossRef]
- Du, J.; Liao, W.; Liu, W.; Deb, D.K.; He, L.; Hsu, P.J.; Nguyen, T.; Zhang, L.; Bissonnette, M.; He, C.; et al. N6-Adenosine Methylation of Socs1 mRNA Is Required to Sustain the Negative Feedback Control of Macrophage Activation. Dev. Cell 2020, 55, 737–753.e737. [Google Scholar] [CrossRef]
- Wang, J.N.; Wang, F.; Ke, J.; Li, Z.; Xu, C.H.; Yang, Q.; Chen, X.; He, X.Y.; He, Y.; Suo, X.G.; et al. Inhibition of METTL3 attenuates renal injury and inflammation by alleviating TAB3 m6A modifications via IGF2BP2-dependent mechanisms. Sci. Transl. Med. 2022, 14, 2709. [Google Scholar] [CrossRef]
- Luo, G.; Zhu, T.; Ren, Z. METTL3 Regulated the Meat Quality of Rex Rabbits by Controlling PCK2 Expression via a YTHDF2-N6-Methyladenosine Axis. Foods 2022, 11, 1549. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xiao, H.; Yang, Y.; Zhang, P.; Yuan, J.; Zhang, W.; Chen, L.; Fan, Y.; Zhang, J.; Cheng, H.; et al. Crosstalk between 5-methylcytosine and N6-methyladenosine machinery defines disease progression, therapeutic response and pharmacogenomic landscape in hepatocellular carcinoma. Mol. Cancer 2023, 22, 5. [Google Scholar] [CrossRef]
- Zhang, D.; Wu, S.; Zhang, X.; Ren, S.; Tang, Z.; Gao, F. Coordinated transcriptional and post-transcriptional epigenetic regulation during skeletal muscle development and growth in pigs. J. Anim. Sci. Biotechnol. 2022, 13, 146. [Google Scholar] [CrossRef]
- Deng, S.; Zhang, J.; Su, J.; Zuo, Z.; Zeng, L.; Liu, K.; Zheng, Y.; Huang, X.; Bai, R.; Zhuang, L.; et al. RNA m6A regulates transcription via DNA demethylation and chromatin accessibility. Nat. Genet. 2022, 54, 1427–1437. [Google Scholar] [CrossRef]
- Yu, J.T.; Hu, X.W.; Chen, H.Y.; Yang, Q.; Li, H.D.; Dong, Y.H.; Zhang, Y.; Wang, J.N.; Jin, J.; Wu, Y.G.; et al. DNA methylation of FTO promotes renal inflammation by enhancing m6A of PPAR-alpha in alcohol-induced kidney injury. Pharmacol. Res. 2021, 163, 105286. [Google Scholar] [CrossRef]
- Cai, D.; Yuan, M.; Jia, Y.; Liu, H.; Hu, Y.; Zhao, R. Maternal gestational betaine supplementation-mediated suppression of hepatic cyclin D2 and presenilin1 gene in newborn piglets is associated with epigenetic regulation of the STAT3-dependent pathway. J. Nutr. Biochem. 2015, 26, 1622–1631. [Google Scholar] [CrossRef]
- Fan, C.; Ma, Y.; Chen, S.; Zhou, Q.; Jiang, H.; Zhang, J.; Wu, F. Comprehensive Analysis of the Transcriptome-Wide m6A Methylation Modification Difference in Liver Fibrosis Mice by High-Throughput m6A Sequencing. Front. Cell Dev. Biol. 2021, 9, 767051. [Google Scholar] [CrossRef]
- Xiao, Y.; Wang, Y.; Tang, Q.; Wei, L.; Zhang, X.; Jia, G. An Elongation- and Ligation-Based qPCR Amplification Method for the Radiolabeling-Free Detection of Locus-Specific N6-Methyladenosine Modification. Angew. Chem. Int. Ed. Engl. 2018, 57, 15995–16000. [Google Scholar] [CrossRef]
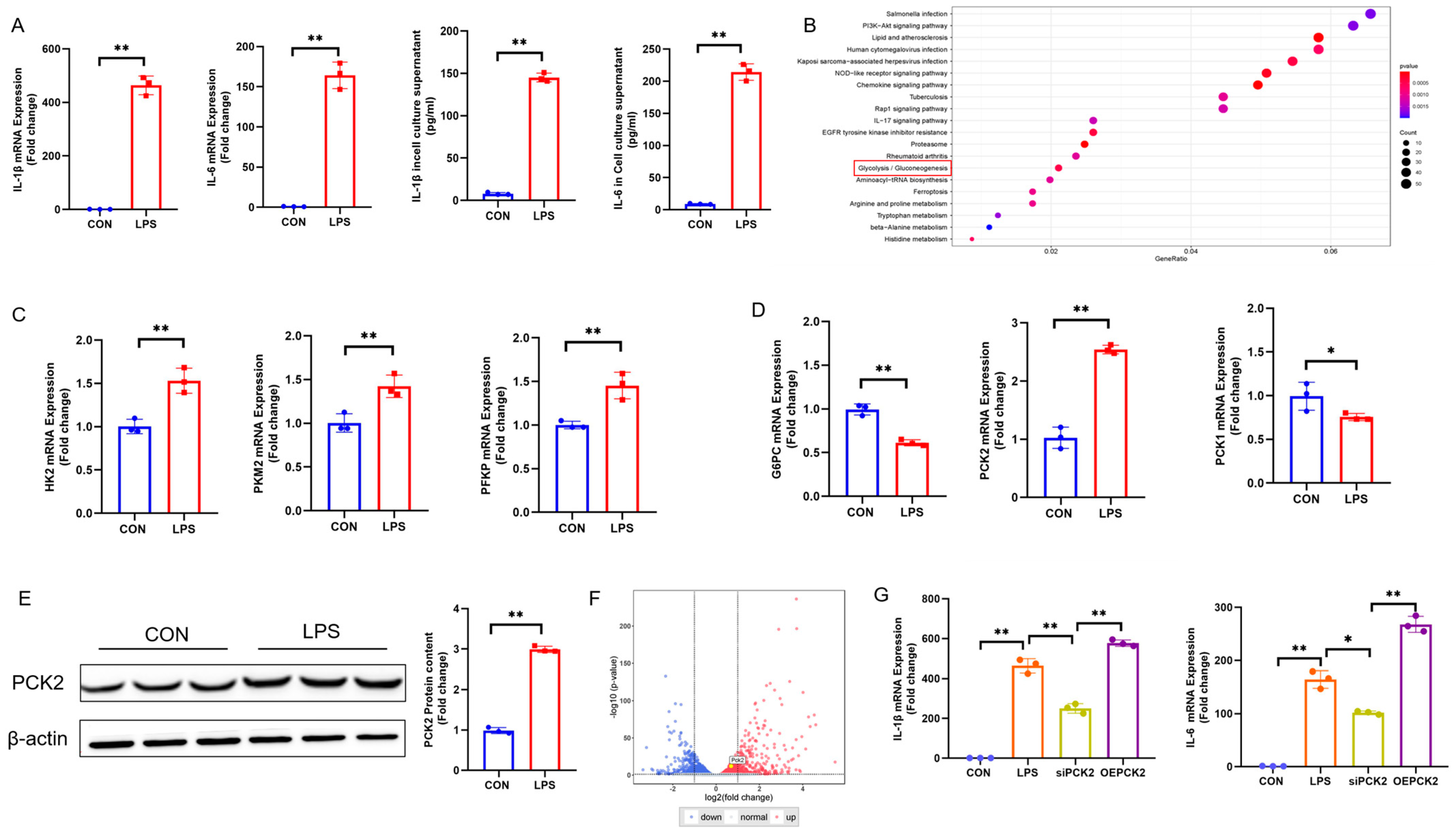
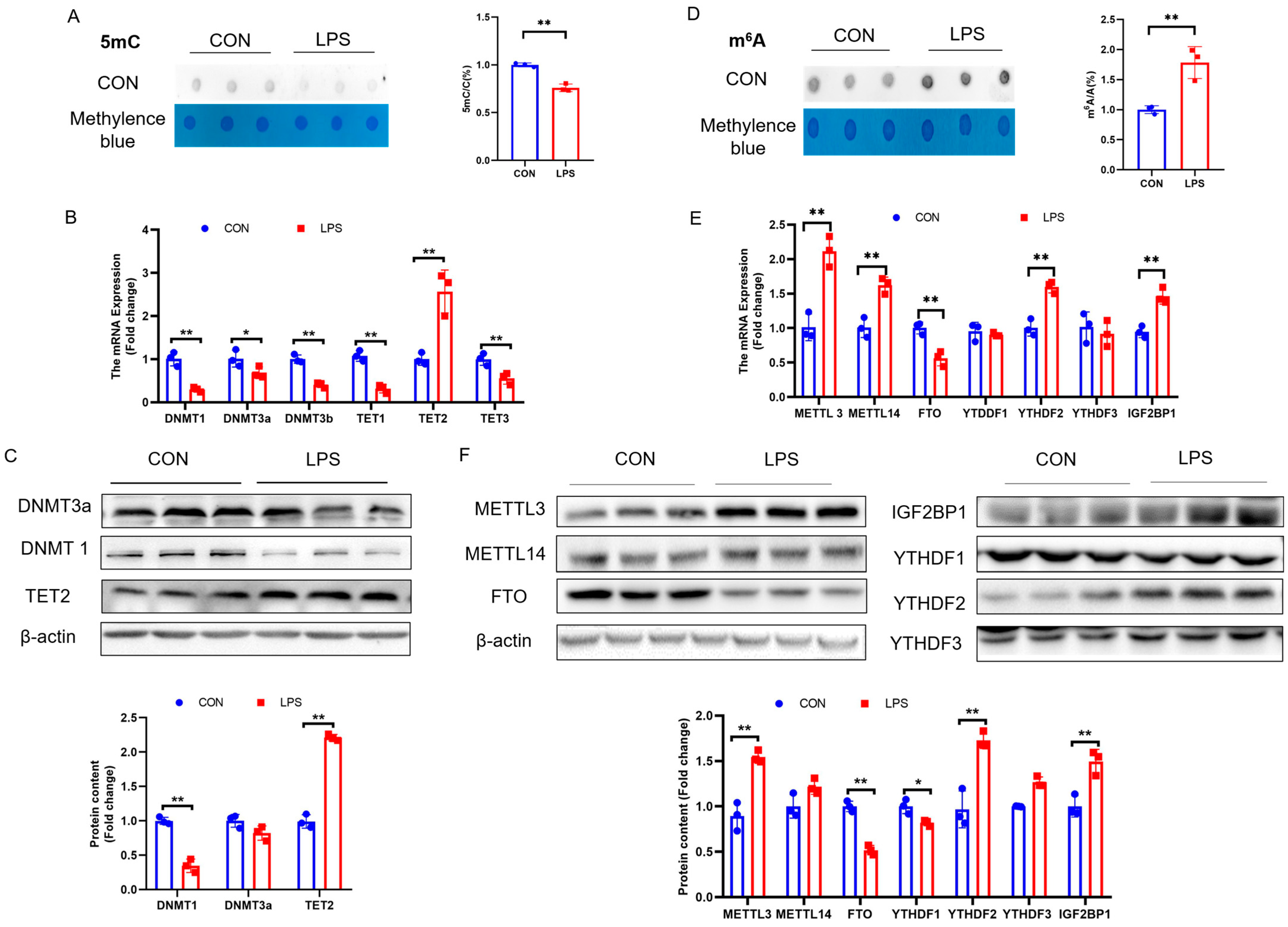
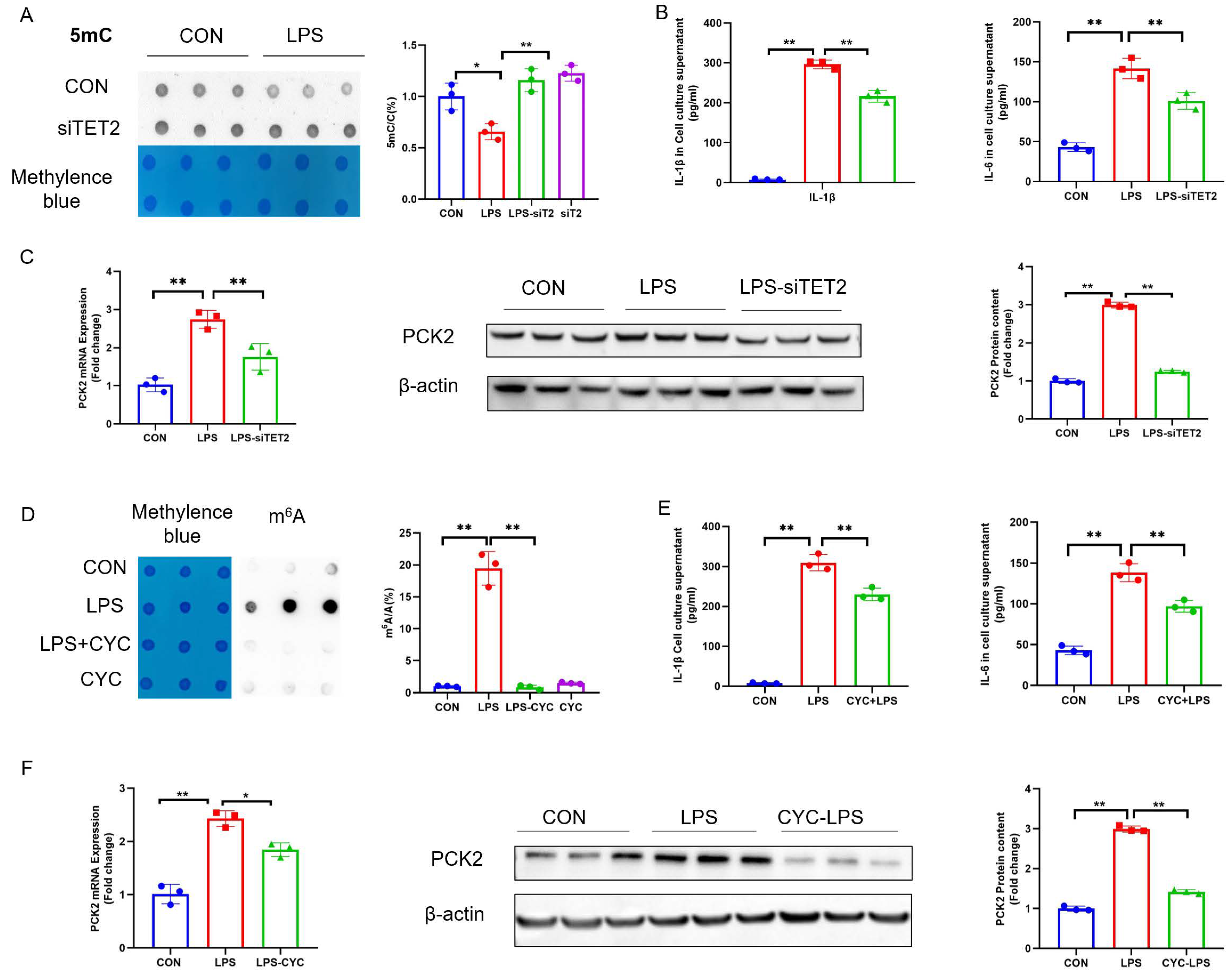
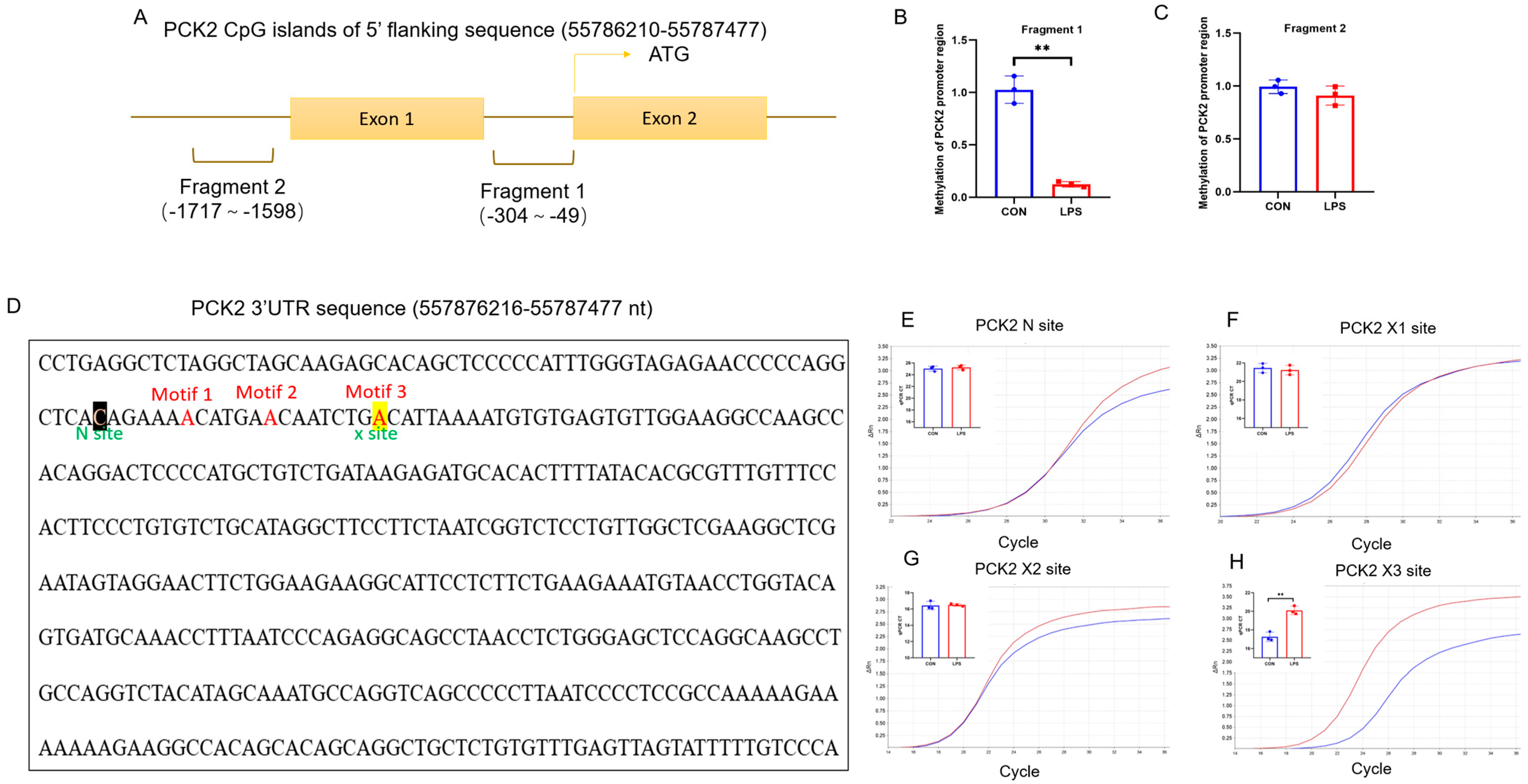

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Yuan, W.; Feng, Y.; Zhao, R. DNA 5mC and RNA m6A Collaborate to Upregulate Phosphoenolpyruvate Carboxykinase 2 for Kupffer Cell Activation. Int. J. Mol. Sci. 2024, 25, 9894. https://doi.org/10.3390/ijms25189894
Zhao Y, Yuan W, Feng Y, Zhao R. DNA 5mC and RNA m6A Collaborate to Upregulate Phosphoenolpyruvate Carboxykinase 2 for Kupffer Cell Activation. International Journal of Molecular Sciences. 2024; 25(18):9894. https://doi.org/10.3390/ijms25189894
Chicago/Turabian StyleZhao, Yulan, Wenbo Yuan, Yue Feng, and Ruqian Zhao. 2024. "DNA 5mC and RNA m6A Collaborate to Upregulate Phosphoenolpyruvate Carboxykinase 2 for Kupffer Cell Activation" International Journal of Molecular Sciences 25, no. 18: 9894. https://doi.org/10.3390/ijms25189894