
Defenders of the Transcriptome: Guard Protein-Mediated mRNA Quality Control in Saccharomyces cerevisiae


Abstract
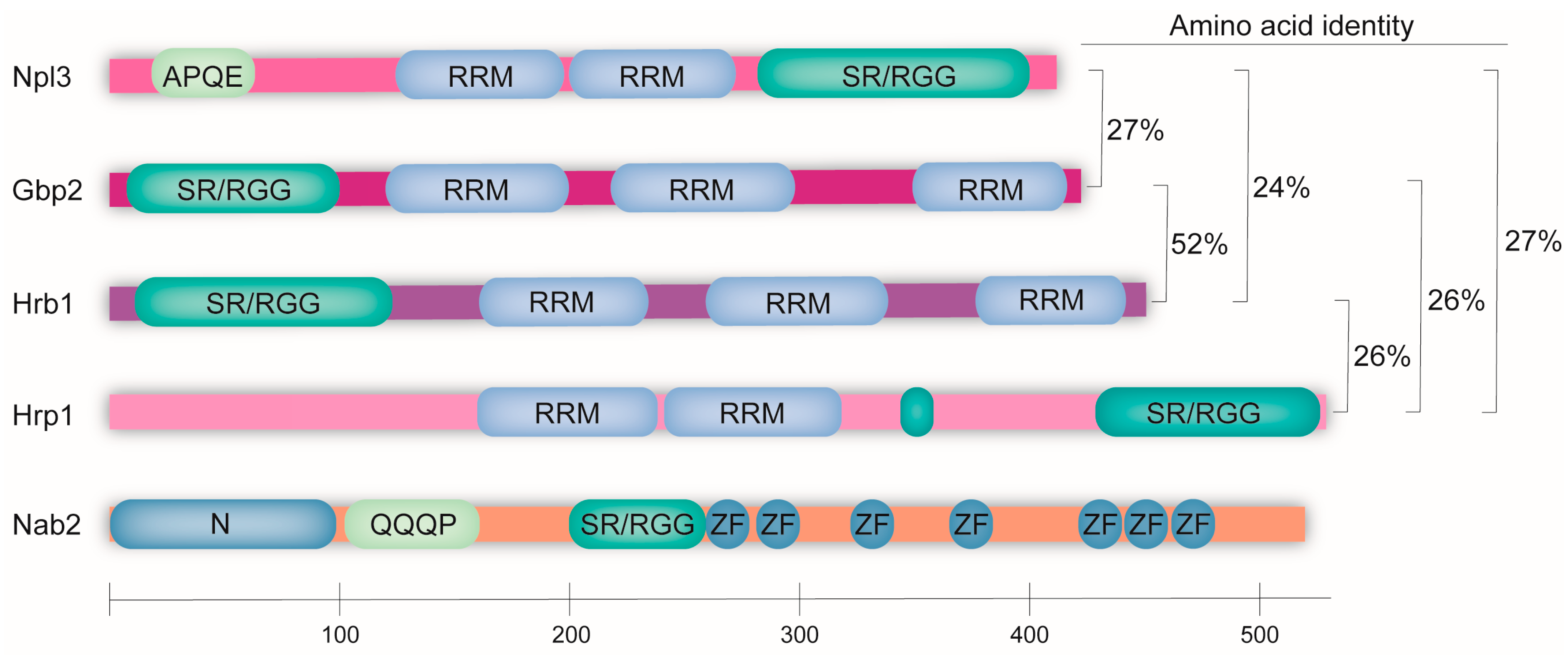
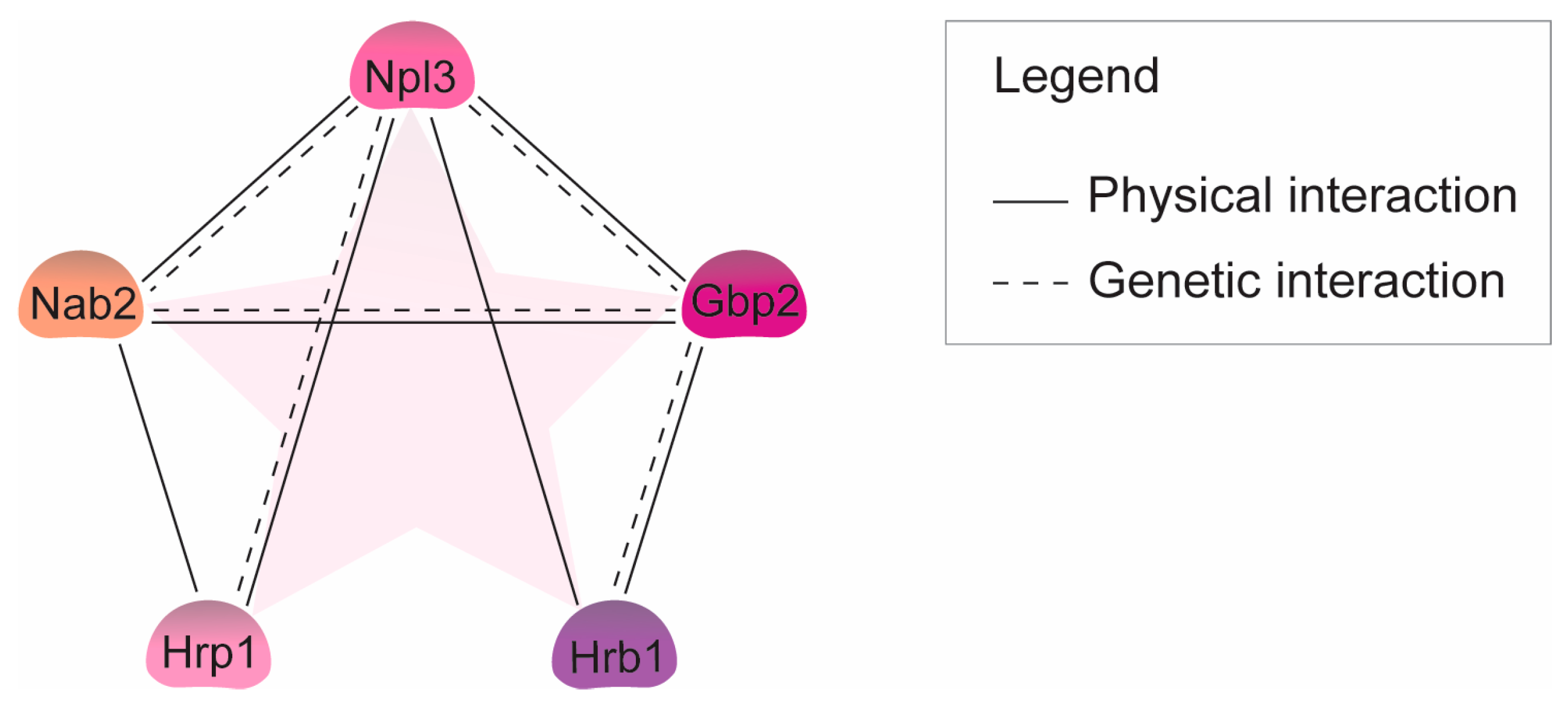
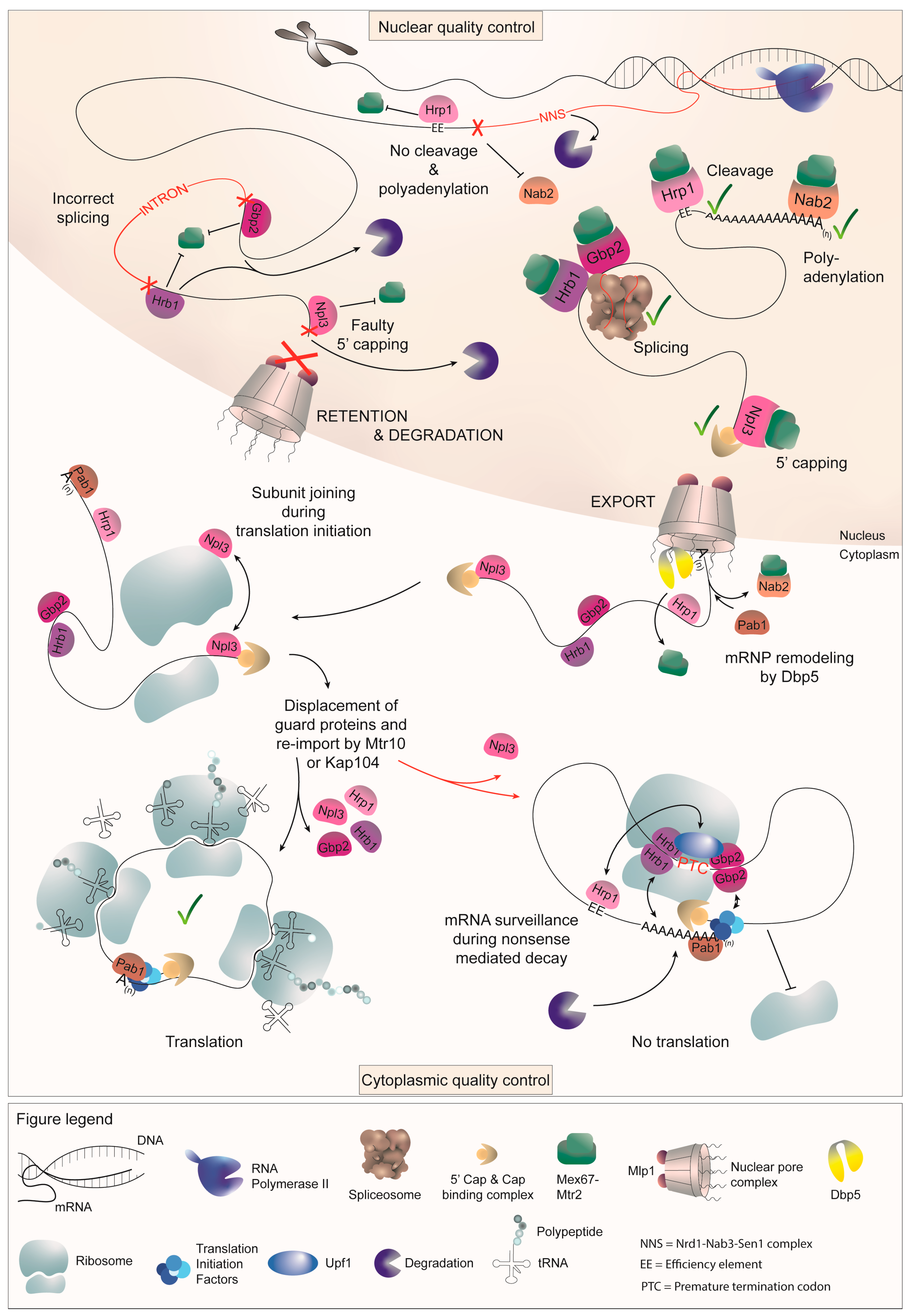
:1. Introduction
2. Maturation of Pre-mRNAs
2.1. Guard Protein-Mediated 5′ Capping Requires Npl3
2.2. Intron Splicing Is Surveilled through the Guard Proteins Gbp2 and Hrb1
2.3. Proper 3′ Cleavage Is Monitored by Hrp1
2.4. Nab2 Surveils the Proper Attachment of a Poly(A) Tail
2.5. Npl3 Participates in 3′ End Quality Control
3. The Guard Proteins Are Subjected to Post-Translational Modification
4. Human Guard Proteins
5. Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ghosh, A.; Lima, C.D. Enzymology of RNA cap synthesis. Wiley Interdiscip. Rev. RNA 2010, 1, 152–172. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rucobo, F.W.; Kohler, R.; van de Waterbeemd, M.; Heck, A.J.; Hemann, M.; Herzog, F.; Stark, H.; Cramer, P. Molecular Basis of Transcription-Coupled Pre-mRNA Capping. Mol. Cell 2015, 58, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Will, C.L.; Luhrmann, R. Spliceosomal UsnRNP biogenesis, structure and function. Curr. Opin. Cell Biol. 2001, 13, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Gonatopoulos-Pournatzis, T.; Cowling, V.H. Cap-binding complex (CBC). Biochem. J. 2014, 457, 231–242. [Google Scholar] [CrossRef]
- Tian, B.; Graber, J.H. Signals for pre-mRNA cleavage and polyadenylation. Wiley Interdiscip. Rev. RNA 2012, 3, 385–396. [Google Scholar] [CrossRef]
- Rondón, A.G.; Mischo, H.E.; Kawauchi, J.; Proudfoot, N.J. Fail-safe transcriptional termination for protein-coding genes in S. cerevisiae. Mol. Cell 2009, 36, 88–98. [Google Scholar] [CrossRef]
- Singh, P.; Chaudhuri, A.; Banerjea, M.; Marathe, N.; Das, B. Nrd1p identifies aberrant and natural exosomal target messages during the nuclear mRNA surveillance in Saccharomyces cerevisiae. Nucleic Acids Res. 2021, 49, 11512–11536. [Google Scholar] [CrossRef]
- Porrua, O.; Libri, D. Transcription termination and the control of the transcriptome: Why, where and how to stop. Nat. Rev. Mol. Cell Biol. 2015, 16, 190–202. [Google Scholar] [CrossRef]
- Mandel, C.R.; Bai, Y.; Tong, L. Protein factors in pre-mRNA 3′-end processing. Cell Mol. Life Sci. 2008, 65, 1099–1122. [Google Scholar] [CrossRef]
- Singleton, D.R.; Chen, S.; Hitomi, M.; Kumagai, C.; Tartakoff, A.M. A yeast protein that bidirectionally affects nucleocytoplasmic transport. J. Cell Sci. 1995, 108 Pt 1, 265–272. [Google Scholar] [CrossRef]
- Deka, P.; Bucheli, M.E.; Moore, C.; Buratowski, S.; Varani, G. Structure of the yeast SR protein Npl3 and Interaction with mRNA 3′-end processing signals. J. Mol. Biol. 2008, 375, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lumbreras, S.; Taverniti, V.; Zorrilla, S.; Séraphin, B.; Pérez-Cañadillas, J.M. Gbp2 interacts with THO/TREX through a novel type of RRM domain. Nucleic Acids Res. 2016, 44, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Marfatia, K.A.; Crafton, E.B.; Green, D.M.; Corbett, A.H. Domain analysis of the Saccharomyces cerevisiae heterogeneous nuclear ribonucleoprotein, Nab2p. Dissecting the requirements for Nab2p-facilitated poly(A) RNA export. J. Biol. Chem. 2003, 278, 6731–6740. [Google Scholar] [CrossRef] [PubMed]
- Busch, A.; Hertel, K.J. Evolution of SR protein and hnRNP splicing regulatory factors. Wiley Interdiscip. Rev. RNA 2012, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Klama, S.; Hirsch, A.G.; Schneider, U.M.; Zander, G.; Seel, A.; Krebber, H. A guard protein mediated quality control mechanism monitors 5′-capping of pre-mRNAs. Nucleic Acids Res. 2022, 50, 11301–11314. [Google Scholar] [CrossRef]
- Zander, G.; Hackmann, A.; Bender, L.; Becker, D.; Lingner, T.; Salinas, G.; Krebber, H. mRNA quality control is bypassed for immediate export of stress-responsive transcripts. Nature 2016, 540, 593–596. [Google Scholar] [CrossRef]
- Zander, G.; Krebber, H. Quick or quality? How mRNA escapes nuclear quality control during stress. RNA Biol. 2017, 14, 1642–1648. [Google Scholar] [CrossRef]
- Hackmann, A.; Wu, H.; Schneider, U.M.; Meyer, K.; Jung, K.; Krebber, H. Quality control of spliced mRNAs requires the shuttling SR proteins Gbp2 and Hrb1. Nat. Commun. 2014, 5, 3123. [Google Scholar] [CrossRef] [PubMed]
- Soheilypour, M.; Mofrad, M.R.K. Regulation of the affinity between RNA-binding proteins and the export receptor enables nuclear basket proteins to distinguish and retain aberrant mRNAs. Sci. Rep. 2016, 6, 35380. [Google Scholar] [CrossRef]
- Grosse, S.; Lu, Y.Y.; Coban, I.; Neumann, B.; Krebber, H. Nuclear SR-protein mediated mRNA quality control is continued in cytoplasmic nonsense-mediated decay. RNA Biol. 2021, 18, 1390–1407. [Google Scholar] [CrossRef]
- Baierlein, C.; Hackmann, A.; Gross, T.; Henker, L.; Hinz, F.; Krebber, H. Monosome formation during translation initiation requires the serine/arginine-rich protein Npl3. Mol. Cell Biol. 2013, 33, 4811–4823. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C.I.; Ruiz-Echevarria, M.J.; Vasudevan, S.; Henry, M.F.; Peltz, S.W. The yeast hnRNP-like protein Hrp1/Nab4 marks a transcript for nonsense-mediated mRNA decay. Mol. Cell 2000, 5, 489–499. [Google Scholar] [CrossRef]
- Senger, B.; Simos, G.; Bischoff, F.R.; Podtelejnikov, A.; Mann, M.; Hurt, E. Mtr10p functions as a nuclear import receptor for the mRNA-binding protein Npl3p. Embo J. 1998, 17, 2196–2207. [Google Scholar] [CrossRef]
- Hacker, S.; Krebber, H. Differential Export Requirements for Shuttling Serine/Arginine-type mRNA-binding Proteins. J. Biol. Chem. 2004, 279, 5049–5052. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.C.; Aitchison, J.D. Kap104p-mediated nuclear import. Nuclear localization signals in mRNA- binding proteins and the role of Ran and Rna. J. Biol. Chem. 1999, 274, 29031–29037. [Google Scholar] [CrossRef]
- Iglesias, N.; Stutz, F. Regulation of mRNP dynamics along the export pathway. FEBS Lett. 2008, 582, 1987–1996. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Querl, L.; Coban, I.; Salinas, G.; Krebber, H. Surveillance of 3′ mRNA cleavage during transcription termination requires CF IB/Hrp1. Nucleic Acids Res. 2023, 51, 8758–8773. [Google Scholar] [CrossRef]
- Wilmes, G.M.; Bergkessel, M.; Bandyopadhyay, S.; Shales, M.; Braberg, H.; Cagney, G.; Collins, S.R.; Whitworth, G.B.; Kress, T.L.; Weissman, J.S.; et al. A genetic interaction map of RNA-processing factors reveals links between Sem1/Dss1-containing complexes and mRNA export and splicing. Mol. Cell 2008, 32, 735–746. [Google Scholar] [CrossRef]
- Vinciguerra, P.; Iglesias, N.; Camblong, J.; Zenklusen, D.; Stutz, F. Perinuclear Mlp proteins downregulate gene expression in response to a defect in mRNA export. Embo J. 2005, 24, 813–823. [Google Scholar] [CrossRef]
- Green, D.M.; Marfatia, K.A.; Crafton, E.B.; Zhang, X.; Cheng, X.; Corbett, A.H. Nab2p Is Required for Poly(A) RNA Export in Saccharomyces cerevisiae and Is Regulated by Arginine Methylation via Hmt1p. J. Biol. Chem. 2002, 277, 7752–7760. [Google Scholar] [CrossRef]
- Furuichi, Y.; LaFiandra, A.; Shatkin, A.J. 5′-Terminal structure and mRNA stability. Nature 1977, 266, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Krogan, N.J.; Vasiljeva, L.; Rando, O.J.; Nedea, E.; Greenblatt, J.F.; Buratowski, S. The yeast Rat1 exonuclease promotes transcription termination by RNA polymerase II. Nature 2004, 432, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Schwer, B.; Saha, N.; Mao, X.; Chen, H.W.; Shuman, S. Structure-function analysis of yeast mRNA cap methyltransferase and high-copy suppression of conditional mutants by AdoMet synthase and the ubiquitin conjugating enzyme Cdc34p. Genetics 2000, 155, 1561–1576. [Google Scholar] [CrossRef]
- Shatkin, A.J. Capping of eucaryotic mRNAs. Cell 1976, 9, 645–653. [Google Scholar] [CrossRef]
- Takagi, T.; Cho, E.J.; Janoo, R.T.; Polodny, V.; Takase, Y.; Keogh, M.C.; Woo, S.A.; Fresco-Cohen, L.D.; Hoffman, C.S.; Buratowski, S. Divergent subunit interactions among fungal mRNA 5′-capping machineries. Eukaryot. Cell 2002, 1, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Ramanathan, A.; Robb, G.B.; Chan, S.H. mRNA capping: Biological functions and applications. Nucleic Acids Res. 2016, 44, 7511–7526. [Google Scholar] [CrossRef]
- Becker, D.; Hirsch, A.G.; Bender, L.; Lingner, T.; Salinas, G.; Krebber, H. Nuclear Pre-snRNA Export Is an Essential Quality Assurance Mechanism for Functional Spliceosomes. Cell Rep. 2019, 27, 3199–3214 e3. [Google Scholar] [CrossRef]
- Hsin, J.-P.; Manley, J.L. The RNA polymerase II CTD coordinates transcription and RNA processing. Genes Dev. 2012, 26, 2119–2137. [Google Scholar] [CrossRef]
- Carrillo Oesterreich, F.; Bieberstein, N.; Neugebauer, K.M. Pause locally, splice globally. Trends Cell Biol. 2011, 21, 328–335. [Google Scholar] [CrossRef]
- Brody, E.; Abelson, J. The “spliceosome”: Yeast pre-messenger RNA associates with a 40S complex in a splicing-dependent reaction. Science 1985, 228, 963–967. [Google Scholar] [CrossRef]
- Plaschka, C.; Newman, A.J.; Nagai, K. Structural Basis of Nuclear pre-mRNA Splicing: Lessons from Yeast. Cold Spring Harb. Perspect. Biol. 2019, 11, a032391. [Google Scholar] [CrossRef] [PubMed]
- Tutucci, E.; Stutz, F. Keeping mRNPs in check during assembly and nuclear export. Nat. Rev. Mol. Cell Biol. 2011, 12, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Nino, C.A.; Herissant, L.; Babour, A.; Dargemont, C. mRNA Nuclear Export in Yeast. Chem. Rev. 2013, 113, 8523–8545. [Google Scholar] [CrossRef] [PubMed]
- Lei, E.P.; Krebber, H.; Silver, P.A. Messenger RNAs are recruited for nuclear export during transcription. Genes Dev. 2001, 15, 1771–1782. [Google Scholar] [CrossRef]
- Hackmann, A.; Gross, T.; Baierlein, C.; Krebber, H. The mRNA export factor Npl3 mediates the nuclear export of large ribosomal subunits. EMBO Rep. 2011, 12, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Hellen, C.U.T. Translation Termination and Ribosome Recycling in Eukaryotes. Cold Spring Harb. Perspect. Biol. 2018, 10, a032656. [Google Scholar] [CrossRef]
- Sherlock, M.E.; Baquero Galvis, L.; Vicens, Q.; Kieft, J.S.; Jagannathan, S. Principles, mechanisms, and biological implications of translation termination-reinitiation. RNA 2023, 29, 865–884. [Google Scholar] [CrossRef]
- Hurt, E.; Luo, M.-J.; Röther, S.; Reed, R.; Strässer, K. Cotranscriptional recruitment of the serine-arginine-rich (SR)-like proteins Gbp2 and Hrb1 to nascent mRNA via the TREX complex. Proc. Natl. Acad. Sci. USA 2004, 101, 1858–1862. [Google Scholar] [CrossRef]
- Windgassen, M.; Krebber, H. Identification of Gbp2 as a novel poly(A)+ RNA-binding protein involved in the cytoplasmic delivery of messenger RNAs in yeast. EMBO Rep. 2003, 4, 278–283. [Google Scholar] [CrossRef]
- Windgassen, M.; Sturm, D.; Cajigas, I.J.; Gonzalez, C.I.; Seedorf, M.; Bastians, H.; Krebber, H. Yeast shuttling SR proteins Npl3p, Gbp2p, and Hrb1p are part of the translating mRNPs, and Npl3p can function as a translational repressor. Mol. Cell Biol. 2004, 24, 10479–10491. [Google Scholar] [CrossRef]
- Houseley, J.; Tollervey, D. The Many Pathways of RNA Degradation. Cell 2009, 136, 763–776. [Google Scholar] [CrossRef] [PubMed]
- LaCava, J.; Houseley, J.; Saveanu, C.; Petfalski, E.; Thompson, E.; Jacquier, A.; Tollervey, D. RNA degradation by the exosome is promoted by a nuclear polyadenylation complex. Cell 2005, 121, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Baejen, C.; Torkler, P.; Gressel, S.; Essig, K.; Söding, J.; Cramer, P. Transcriptome Maps of mRNP Biogenesis Factors Define Pre-mRNA Recognition. Mol. Cell 2014, 55, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Tuck, A.C.; Tollervey, D. A Transcriptome-wide Atlas of RNP Composition Reveals Diverse Classes of mRNAs and lncRNAs. Cell 2013, 154, 996–1009. [Google Scholar] [CrossRef]
- Muhlrad, D.; Parker, R. Premature translational termination triggers mRNA decapping. Nature 1994, 370, 578–581. [Google Scholar] [CrossRef]
- He, F.; Jacobson, A. Nonsense-Mediated mRNA Decay: Degradation of Defective Transcripts Is Only Part of the Story. Annu. Rev. Genet. 2015, 49, 339–366. [Google Scholar] [CrossRef]
- Henry, M.; Borland, C.Z.; Bossie, M.; Silver, P.A. Potential RNA binding proteins in Saccharomyces cerevisiae identified as suppressors of temperature-sensitive mutations in NPL3. Genetics 1996, 142, 103–115. [Google Scholar] [CrossRef]
- Kessler, M.M.; Henry, M.F.; Shen, E.; Zhao, J.; Gross, S.; Silver, P.A.; Moore, C.L. Hrp1, a sequence-specific RNA-binding protein that shuttles between the nucleus and the cytoplasm, is required for mRNA 3′-end formation in yeast. Genes Dev. 1997, 11, 2545–2556. [Google Scholar] [CrossRef]
- Henry, M.F.; Silver, P.A. A novel methyltransferase (Hmt1p) modifies poly(A)+-RNA-binding proteins. Mol. Cell Biol. 1996, 16, 3668–3678. [Google Scholar] [CrossRef]
- Amrani, N.; Minet, M.; Le Gouar, M.; Lacroute, F.; Wyers, F. Yeast Pab1 interacts with Rna15 and participates in the control of the poly(A) tail length in vitro. Mol. Cell Biol. 1997, 17, 3694–3701. [Google Scholar] [CrossRef]
- Hammell, C.M.; Gross, S.; Zenklusen, D.; Heath, C.V.; Stutz, F.; Moore, C.; Cole, C.N. Coupling of termination, 3′ processing, and mRNA export. Mol. Cell Biol. 2002, 22, 6441–6457. [Google Scholar] [CrossRef]
- Carroll, K.L.; Pradhan, D.A.; Granek, J.A.; Clarke, N.D.; Corden, J.L. Identification of cis elements directing termination of yeast nonpolyadenylated snoRNA transcripts. Mol. Cell Biol. 2004, 24, 6241–6252. [Google Scholar] [CrossRef]
- Creamer, T.J.; Darby, M.M.; Jamonnak, N.; Schaughency, P.; Hao, H.; Wheelan, S.J.; Corden, J.L. Transcriptome-wide binding sites for components of the Saccharomyces cerevisiae non-poly(A) termination pathway: Nrd1, Nab3, and Sen1. PLoS Genet. 2011, 7, e1002329. [Google Scholar] [CrossRef]
- Nedea, E.; Nalbant, D.; Xia, D.; Theoharis, N.T.; Suter, B.; Richardson, C.J.; Tatchell, K.; Kislinger, T.; Greenblatt, J.F.; Nagy, P.L. The Glc7 phosphatase subunit of the cleavage and polyadenylation factor is essential for transcription termination on snoRNA genes. Mol. Cell 2008, 29, 577–587. [Google Scholar] [CrossRef]
- Vasiljeva, L.; Buratowski, S. Nrd1 interacts with the nuclear exosome for 3′ processing of RNA polymerase II transcripts. Mol. Cell 2006, 21, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Lemay, J.F.; Bachand, F. Fail-safe transcription termination: Because one is never enough. RNA Biol. 2015, 12, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Hector, R.E.; Nykamp, K.R.; Dheur, S.; Anderson, J.T.; Non, P.J.; Urbinati, C.R.; Wilson, S.M.; Minvielle-Sebastia, L.; Swanson, M.S. Dual requirement for yeast hnRNP Nab2p in mRNA poly(A) tail length control and nuclear export. Embo J. 2002, 21, 1800–1810. [Google Scholar] [CrossRef]
- Green, D.M.; Johnson, C.P.; Hagan, H.; Corbett, A.H. The C-terminal domain of myosin-like protein 1 (Mlp1p) is a docking site for heterogeneous nuclear ribonucleoproteins that are required for mRNA export. Proc. Natl. Acad. Sci. USA 2003, 100, 1010–1015. [Google Scholar] [CrossRef]
- Tran, E.J.; Zhou, Y.; Corbett, A.H.; Wente, S.R. The DEAD-box protein Dbp5 controls mRNA export by triggering specific RNA:protein remodeling events. Mol. Cell 2007, 28, 850–859. [Google Scholar] [CrossRef]
- Kessler, S.H.; Sachs, A.B. RNA recognition motif 2 of yeast Pab1p is required for its functional interaction with eukaryotic translation initiation factor 4G. Mol. Cell Biol. 1998, 18, 51–57. [Google Scholar] [CrossRef]
- Rajkowitsch, L.; Vilela, C.; Berthelot, K.; Ramirez, C.V.; McCarthy, J.E. Reinitiation and recycling are distinct processes occurring downstream of translation termination in yeast. J. Mol. Biol. 2004, 335, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Batisse, J.; Batisse, C.; Budd, A.; Bottcher, B.; Hurt, E. Purification of nuclear poly(A)-binding protein Nab2 reveals association with the yeast transcriptome and a messenger ribonucleoprotein core structure. J. Biol. Chem. 2009, 284, 34911–34917. [Google Scholar] [CrossRef]
- Aibara, S.; Gordon, J.M.; Riesterer, A.S.; McLaughlin, S.H.; Stewart, M. Structural basis for the dimerization of Nab2 generated by RNA binding provides insight into its contribution to both poly(A) tail length determination and transcript compaction in Saccharomyces cerevisiae. Nucleic Acids Res. 2017, 45, 1529–1538. [Google Scholar] [CrossRef]
- Brockmann, C.; Soucek, S.; Kuhlmann, S.I.; Mills-Lujan, K.; Kelly, S.M.; Yang, J.C.; Iglesias, N.; Stutz, F.; Corbett, A.H.; Neuhaus, D.; et al. Structural basis for polyadenosine-RNA binding by Nab2 Zn fingers and its function in mRNA nuclear export. Structure 2012, 20, 1007–1018. [Google Scholar] [CrossRef]
- Dermody, J.L.; Dreyfuss, J.M.; Villen, J.; Ogundipe, B.; Gygi, S.P.; Park, P.J.; Ponticelli, A.S.; Moore, C.L.; Buratowski, S.; Bucheli, M.E. Unphosphorylated SR-like protein Npl3 stimulates RNA polymerase II elongation. PLoS ONE 2008, 3, e3273. [Google Scholar] [CrossRef]
- Bucheli, M.E.; Buratowski, S. Npl3 is an antagonist of mRNA 3′ end formation by RNA polymerase II. Embo J. 2005, 24, 2150–2160. [Google Scholar] [CrossRef]
- Bernstein, J.; Toth, E.A. Yeast nuclear RNA processing. World J. Biol. Chem. 2012, 3, 7–26. [Google Scholar] [CrossRef]
- Holmes, R.K.; Tuck, A.C.; Zhu, C.; Dunn-Davies, H.R.; Kudla, G.; Clauder-Munster, S.; Granneman, S.; Steinmetz, L.M.; Guthrie, C.; Tollervey, D. Loss of the Yeast SR Protein Npl3 Alters Gene Expression Due to Transcription Readthrough. PLoS Genet. 2015, 11, e1005735. [Google Scholar] [CrossRef]
- Gilbert, W.; Guthrie, C. The glc7p nuclear phosphatase promotes mRNA export by facilitating association of mex67p with mRNA. Mol. Cell 2004, 13, 201–212. [Google Scholar] [CrossRef]
- Weiss, V.H.; McBride, A.E.; Soriano, M.A.; Filman, D.J.; Silver, P.A.; Hogle, J.M. The structure and oligomerization of the yeast arginine methyltransferase, Hmt1. Nat. Struct. Biol. 2000, 7, 1165–1171. [Google Scholar] [CrossRef]
- Gilbert, W.; Siebel, C.W.; Guthrie, C. Phosphorylation by Sky1p promotes Npl3p shuttling and mRNA dissociation. RNA 2001, 7, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Mizuno, T.; Wada, K.; Hagiwara, M. Novel RING finger proteins, Air1p and Air2p, interact with Hmt1p and inhibit the arginine methylation of Npl3p. J. Biol. Chem. 2000, 275, 32793–32799. [Google Scholar] [CrossRef] [PubMed]
- Hamey, J.J.; Nguyen, A.; Wilkins, M.R. Discovery of Arginine Methylation, Phosphorylation, and Their Co-occurrence in Condensate-Associated Proteins in Saccharomyces cerevisiae. J. Proteome Res. 2021, 20, 2420–2434. [Google Scholar] [CrossRef]
- Valentini, S.R.; Weiss, V.H.; Silver, P.A. Arginine methylation and binding of Hrp1p to the efficiency element for mRNA 3′-end formation. RNA 1999, 5, 272–280. [Google Scholar] [CrossRef]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Véronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; André, B.; et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.C.; Bachand, F.; McBride, A.E.; Komili, S.; Casolari, J.M.; Silver, P.A. Arginine methyltransferase affects interactions and recruitment of mRNA processing and export factors. Genes Dev. 2004, 18, 2024–2035. [Google Scholar] [CrossRef]
- Shen, E.C.; Henry, M.F.; Weiss, V.H.; Valentini, S.R.; Silver, P.A.; Lee, M.S. Arginine methylation facilitates the nuclear export of hnRNP proteins. Genes Dev. 1998, 12, 679–691. [Google Scholar] [CrossRef]
- Xu, C.; Henry, M.F. Nuclear export of hnRNP Hrp1p and nuclear export of hnRNP Npl3p are linked and influenced by the methylation state of Npl3p. Mol. Cell Biol. 2004, 24, 10742–10756. [Google Scholar] [CrossRef]
- Swaney, D.L.; Beltrao, P.; Starita, L.; Guo, A.; Rush, J.; Fields, S.; Krogan, N.J.; Villén, J. Global analysis of phosphorylation and ubiquitylation cross-talk in protein degradation. Nat. Methods 2013, 10, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Lanz, M.C.; Yugandhar, K.; Gupta, S.; Sanford, E.J.; Faça, V.M.; Vega, S.; Joiner, A.M.N.; Fromme, J.C.; Yu, H.; Smolka, M.B. In-depth and 3-dimensional exploration of the budding yeast phosphoproteome. EMBO Rep. 2021, 22, e51121. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, N.R.; Owens, S.N.; Ito, M.; Boinapalli, J.V.; Poa, P.; Ditzel, A.; Kopparapu, S.; Mahalawat, M.; Davies, O.R.; Collins, S.R.; et al. SUMO is a pervasive regulator of meiosis. Elife 2021, 10, e57720. [Google Scholar] [CrossRef] [PubMed]
- Weinert, B.T.; Schölz, C.; Wagner, S.A.; Iesmantavicius, V.; Su, D.; Daniel, J.A.; Choudhary, C. Lysine succinylation is a frequently occurring modification in prokaryotes and eukaryotes and extensively overlaps with acetylation. Cell Rep. 2013, 4, 842–851. [Google Scholar] [CrossRef]
- Henriksen, P.; Wagner, S.A.; Weinert, B.T.; Sharma, S.; Bacinskaja, G.; Rehman, M.; Juffer, A.H.; Walther, T.C.; Lisby, M.; Choudhary, C. Proteome-wide analysis of lysine acetylation suggests its broad regulatory scope in Saccharomyces cerevisiae. Mol. Cell Proteom. 2012, 11, 1510–1522. [Google Scholar] [CrossRef] [PubMed]
- Erce, M.A.; Abeygunawardena, D.; Low, J.K.; Hart-Smith, G.; Wilkins, M.R. Interactions affected by arginine methylation in the yeast protein-protein interaction network. Mol. Cell Proteom. 2013, 12, 3184–3198. [Google Scholar] [CrossRef]
- Geuens, T.; Bouhy, D.; Timmerman, V. The hnRNP family: Insights into their role in health and disease. Hum. Genet. 2016, 135, 851–867. [Google Scholar] [CrossRef] [PubMed]
- Thandapani, P.; O’Connor, T.R.; Bailey, T.L.; Richard, S. Defining the RGG/RG motif. Mol. Cell 2013, 50, 613–623. [Google Scholar] [CrossRef]
- Ozdilek, B.A.; Thompson, V.F.; Ahmed, N.S.; White, C.I.; Batey, R.T.; Schwartz, J.C. Intrinsically disordered RGG/RG domains mediate degenerate specificity in RNA binding. Nucleic Acids Res. 2017, 45, 7984–7996. [Google Scholar] [CrossRef]
- Slišković, I.; Eich, H.; Müller-McNicoll, M. Exploring the multifunctionality of SR proteins. Biochem. Soc. Trans. 2022, 50, 187–198. [Google Scholar] [CrossRef]
- Jeong, S. SR Proteins: Binders, Regulators, and Connectors of RNA. Mol. Cells 2017, 40, 1–9. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Krebber, H. Nuclear mRNA Quality Control and Cytoplasmic NMD Are Linked by the Guard Proteins Gbp2 and Hrb1. Int. J. Mol. Sci. 2021, 22, 11275. [Google Scholar] [CrossRef]
- Huang, Y.; Gattoni, R.; Stevenin, J.; Steitz, J.A. SR splicing factors serve as adapter proteins for TAP-dependent mRNA export. Mol. Cell 2003, 11, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Zahler, A.M.; Lane, W.S.; Stolk, J.A.; Roth, M.B. SR proteins: A conserved family of pre-mRNA splicing factors. Genes Dev. 1992, 6, 837–847. [Google Scholar] [CrossRef] [PubMed]
- Manley, J.L.; Krainer, A.R. A rational nomenclature for serine/arginine-rich protein splicing factors (SR proteins). Genes Dev. 2010, 24, 1073–1074. [Google Scholar] [CrossRef] [PubMed]
- Cáceres, J.F.; Misteli, T.; Screaton, G.R.; Spector, D.L.; Krainer, A.R. Role of the modular domains of SR proteins in subnuclear localization and alternative splicing specificity. J. Cell Biol. 1997, 138, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Caceres, J.F.; Screaton, G.R.; Krainer, A.R. A specific subset of SR proteins shuttles continuously between the nucleus and the cytoplasm. Genes Dev. 1998, 12, 55–66. [Google Scholar] [CrossRef]
- Cazalla, D.; Zhu, J.; Manche, L.; Huber, E.; Krainer, A.R.; Cáceres, J.F. Nuclear export and retention signals in the RS domain of SR proteins. Mol. Cell Biol. 2002, 22, 6871–6882. [Google Scholar] [CrossRef]
- Sapra, A.K.; Ankö, M.L.; Grishina, I.; Lorenz, M.; Pabis, M.; Poser, I.; Rollins, J.; Weiland, E.M.; Neugebauer, K.M. SR protein family members display diverse activities in the formation of nascent and mature mRNPs in vivo. Mol. Cell 2009, 34, 179–190. [Google Scholar] [CrossRef]
- Botti, V.; McNicoll, F.; Steiner, M.C.; Richter, F.M.; Solovyeva, A.; Wegener, M.; Schwich, O.D.; Poser, I.; Zarnack, K.; Wittig, I.; et al. Cellular differentiation state modulates the mRNA export activity of SR proteins. J. Cell Biol. 2017, 216, 1993–2009. [Google Scholar] [CrossRef]
- Li, D.; Yu, W.; Lai, M. Towards understandings of serine/arginine-rich splicing factors. Acta Pharm. Sin. B 2023, 13, 3181–3207. [Google Scholar] [CrossRef]
- Sanford, J.R.; Ellis, J.; Cáceres, J.F. Multiple roles of arginine/serine-rich splicing factors in RNA processing. Biochem. Soc. Trans. 2005, 33 Pt 3, 443–446. [Google Scholar] [CrossRef]
- Zhong, X.Y.; Wang, P.; Han, J.; Rosenfeld, M.G.; Fu, X.D. SR proteins in vertical integration of gene expression from transcription to RNA processing to translation. Mol. Cell 2009, 35, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.; Allemand, E.; Zhang, Z.; Karni, R.; Myers, M.P.; Krainer, A.R. Arginine methylation controls the subcellular localization and functions of the oncoprotein splicing factor SF2/ASF. Mol. Cell Biol. 2010, 30, 2762–2774. [Google Scholar] [CrossRef] [PubMed]
- Mure, F.; Corbin, A.; Benbahouche, N.E.H.; Bertrand, E.; Manet, E.; Gruffat, H. The splicing factor SRSF3 is functionally connected to the nuclear RNA exosome for intronless mRNA decay. Sci. Rep. 2018, 8, 12901. [Google Scholar] [CrossRef] [PubMed]
- Corbo, C.; Orrù, S.; Salvatore, F. SRp20: An overview of its role in human diseases. Biochem. Biophys. Res. Commun. 2013, 436, 1–5. [Google Scholar] [CrossRef]
- Das, S.; Krainer, A.R. Emerging functions of SRSF1, splicing factor and oncoprotein, in RNA metabolism and cancer. Mol. Cancer Res. 2014, 12, 1195–1204. [Google Scholar] [CrossRef]
- de Bruin, R.G.; Rabelink, T.J.; van Zonneveld, A.J.; van der Veer, E.P. Emerging roles for RNA-binding proteins as effectors and regulators of cardiovascular disease. Eur. Heart J. 2017, 38, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Fasken, M.B.; Corbett, A.H. Links between mRNA Splicing, mRNA Quality Control, and Intellectual Disability. RNA Dis. 2016, 3, e1448. [Google Scholar]
- Zheng, X.; Peng, Q.; Wang, L.; Zhang, X.; Huang, L.; Wang, J.; Qin, Z. Serine/arginine-rich splicing factors: The bridge linking alternative splicing and cancer. Int. J. Biol. Sci. 2020, 16, 2442–2453. [Google Scholar] [CrossRef]
- Liu, J.; Huang, B.; Xiao, Y.; Xiong, H.M.; Li, J.; Feng, D.Q.; Chen, X.M.; Zhang, H.B.; Wang, X.Z. Aberrant expression of splicing factors in newly diagnosed acute myeloid leukemia. Onkologie 2012, 35, 335–340. [Google Scholar] [CrossRef]
- Wong, J.; Garner, B.; Halliday, G.M.; Kwok, J.B. Srp20 regulates TrkB pre-mRNA splicing to generate TrkB-Shc transcripts with implications for Alzheimer’s disease. J. Neurochem. 2012, 123, 159–171. [Google Scholar] [CrossRef]
- Wagner, R.E.; Frye, M. Noncanonical functions of the serine-arginine-rich splicing factor (SR) family of proteins in development and disease. Bioessays 2021, 43, e2000242. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.M.; Leung, S.W.; Pak, C.; Banerjee, A.; Moberg, K.H.; Corbett, A.H. A conserved role for the zinc finger polyadenosine RNA binding protein, ZC3H14, in control of poly(A) tail length. RNA 2014, 20, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Corgiat, E.B.; List, S.M.; Rounds, J.C.; Corbett, A.H.; Moberg, K.H. The RNA-binding protein Nab2 regulates the proteome of the developing Drosophila brain. J. Biol. Chem. 2021, 297, 100877. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Zhang, S.; Kim, B.; Hull, V.L.; Xu, J.; Prabhu, P.; Gregory, M.; Martinez-Cerdeno, V.; Zhan, X.; et al. PARP1-mediated PARylation activity is essential for oligodendroglial differentiation and CNS myelination. Cell Rep. 2021, 37, 109695. [Google Scholar] [CrossRef] [PubMed]
- Llères, D.; Denegri, M.; Biggiogera, M.; Ajuh, P.; Lamond, A.I. Direct interaction between hnRNP-M and CDC5L/PLRG1 proteins affects alternative splice site choice. EMBO Rep. 2010, 11, 445–451. [Google Scholar] [CrossRef]
- Sauliere, J.; Murigneux, V.; Wang, Z.; Marquenet, E.; Barbosa, I.; Le Tonqueze, O.; Audic, Y.; Paillard, L.; Roest Crollius, H.; Le Hir, H. CLIP-seq of eIF4AIII reveals transcriptome-wide mapping of the human exon junction complex. Nat. Struct. Mol. Biol. 2012, 19, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Boehm, V.; Gehring, N.H. Exon Junction Complexes: Supervising the Gene Expression Assembly Line. Trends Genet. 2016, 32, 724–735. [Google Scholar] [CrossRef]
- Lejeune, F.; Ishigaki, Y.; Li, X.; Maquat, L.E. The exon junction complex is detected on CBP80-bound but not eIF4E-bound mRNA in mammalian cells: Dynamics of mRNP remodeling. Embo J. 2002, 21, 3536–3545. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Karousis, E.D.; Bourquin, J.; Bruggmann, R.; Mühlemann, O. Transcriptome-wide identification of NMD-targeted human mRNAs reveals extensive redundancy between SMG6- and SMG7-mediated degradation pathways. RNA 2017, 23, 189–201. [Google Scholar] [CrossRef]
- Nagy, E.; Maquat, L.E. A rule for termination-codon position within intron-containing genes: When nonsense affects RNA abundance. Trends Biochem. Sci. 1998, 23, 198–199. [Google Scholar] [CrossRef]
- Kataoka, N.; Yong, J.; Kim, V.N.; Velazquez, F.; Perkinson, R.A.; Wang, F.; Dreyfuss, G. Pre-mRNA splicing imprints mRNA in the nucleus with a novel RNA-binding protein that persists in the cytoplasm. Mol. Cell 2000, 6, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, T.; Ishigaki, Y. C-terminal short arginine/serine repeat sequence-dependent regulation of Y14 (RBM8A) localization. Sci. Rep. 2018, 8, 612. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kucukural, A.; Cenik, C.; Leszyk, J.D.; Shaffer, S.A.; Weng, Z.; Moore, M.J. The cellular EJC interactome reveals higher-order mRNP structure and an EJC-SR protein nexus. Cell 2012, 151, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Neuveglise, C.; Marck, C.; Gaillardin, C. The intronome of budding yeasts. Comptes Rendus Biol. 2011, 334, 662–670. [Google Scholar] [CrossRef]
- Parenteau, J.; Durand, M.; Véronneau, S.; Lacombe, A.A.; Morin, G.; Guérin, V.; Cecez, B.; Gervais-Bird, J.; Koh, C.S.; Brunelle, D.; et al. Deletion of many yeast introns reveals a minority of genes that require splicing for function. Mol. Biol. Cell 2008, 19, 1932–1941. [Google Scholar] [CrossRef] [PubMed]
- Piovesan, A.; Antonaros, F.; Vitale, L.; Strippoli, P.; Pelleri, M.C.; Caracausi, M. Human protein-coding genes and gene feature statistics in 2019. BMC Res. Notes 2019, 12, 315. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, P.; Hanicinec, V.; Soucek, P. The position of the longest intron is related to biological functions in some human genes. Front. Genet. 2022, 13, 1085139. [Google Scholar] [CrossRef]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef]
- Coban, I.; Lamping, J.P.; Hirsch, A.G.; Wasilewski, S.; Shomroni, O.; Giesbrecht, O.; Salinas, G.; Krebber, H. dsRNA formation leads to preferential nuclear export and gene expression. Nature 2024, 631, 432–438. [Google Scholar] [CrossRef]
- Wu, H.; Becker, D.; Krebber, H. Telomerase RNA TLC1 Shuttling to the Cytoplasm Requires mRNA Export Factors and Is Important for Telomere Maintenance. Cell Rep. 2014, 8, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, A.G.; Becker, D.; Lamping, J.P.; Krebber, H. Unraveling the stepwise maturation of the yeast telomerase including a Cse1 and Mtr10 mediated quality control checkpoint. Sci. Rep. 2021, 11, 22174. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Npl3 | Gbp2 | Hrb1 | Hrp1 | Nab2 | |
|---|---|---|---|---|---|
| Alternative names | Nab1 | Rlf7 | Tom34 | Nab4 | - |
| Mtr13 | Nab5 | ||||
| Mts1 | CFIB | ||||
| Nop3 | |||||
| Length | 414 aa | 427 aa | 454 aa | 534 aa | 525 aa |
| Molecular weight | 45.4 kDa | 48.7 kDa | 52.1 kDa | 59.6 kDa | 58.3 kDa |
| Median abundance (mol./cell) | 34,936 ± 26,291 | 3171 ± 1870 | 5191 ± 1126 | 13,717 ± 8354 | 8716 ± 2946 |
| Export receptor | Mex67 | Mex67 | Mex67 | Mex67 | Mex67 |
| Import receptor | Mtr10 | Mtr10 | Mtr10 | Kap104 | Kap104 |
| Essential in yeast? | No | No | No | Yes | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Querl, L.; Krebber, H. Defenders of the Transcriptome: Guard Protein-Mediated mRNA Quality Control in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2024, 25, 10241. https://doi.org/10.3390/ijms251910241
Querl L, Krebber H. Defenders of the Transcriptome: Guard Protein-Mediated mRNA Quality Control in Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2024; 25(19):10241. https://doi.org/10.3390/ijms251910241
Chicago/Turabian StyleQuerl, Luisa, and Heike Krebber. 2024. "Defenders of the Transcriptome: Guard Protein-Mediated mRNA Quality Control in Saccharomyces cerevisiae" International Journal of Molecular Sciences 25, no. 19: 10241. https://doi.org/10.3390/ijms251910241