

New Pipeline for Analysing Fruit Proteolytic Products Used as Digestive Health Nutraceuticals
 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Biochemical Properties
2.1.1. Total Soluble Protein
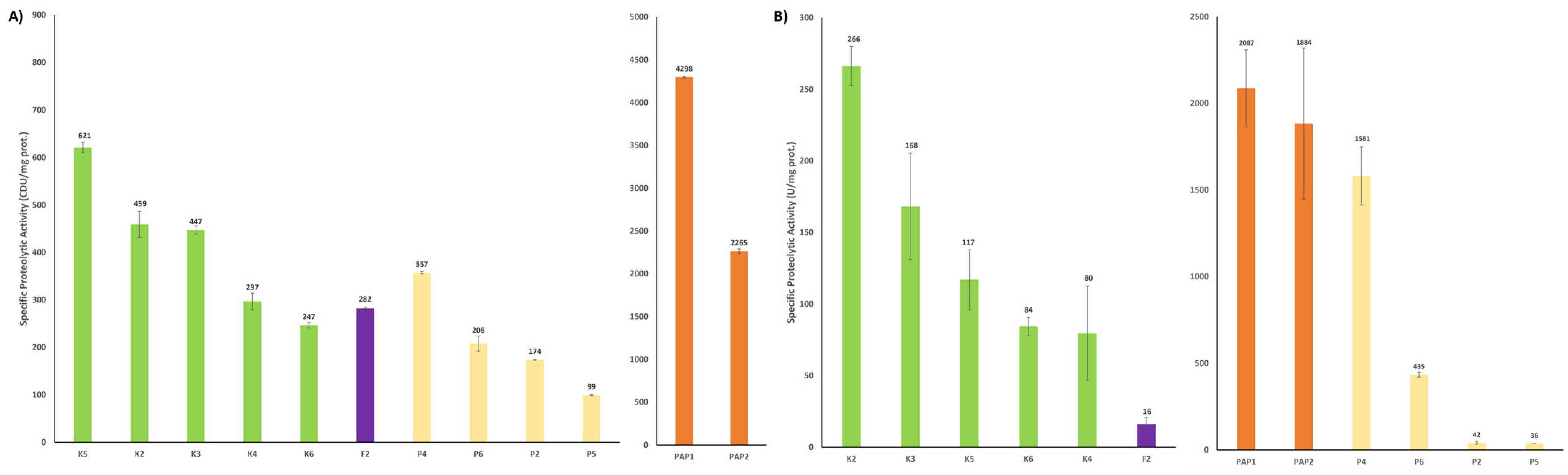
2.1.2. Proteolytic Activity
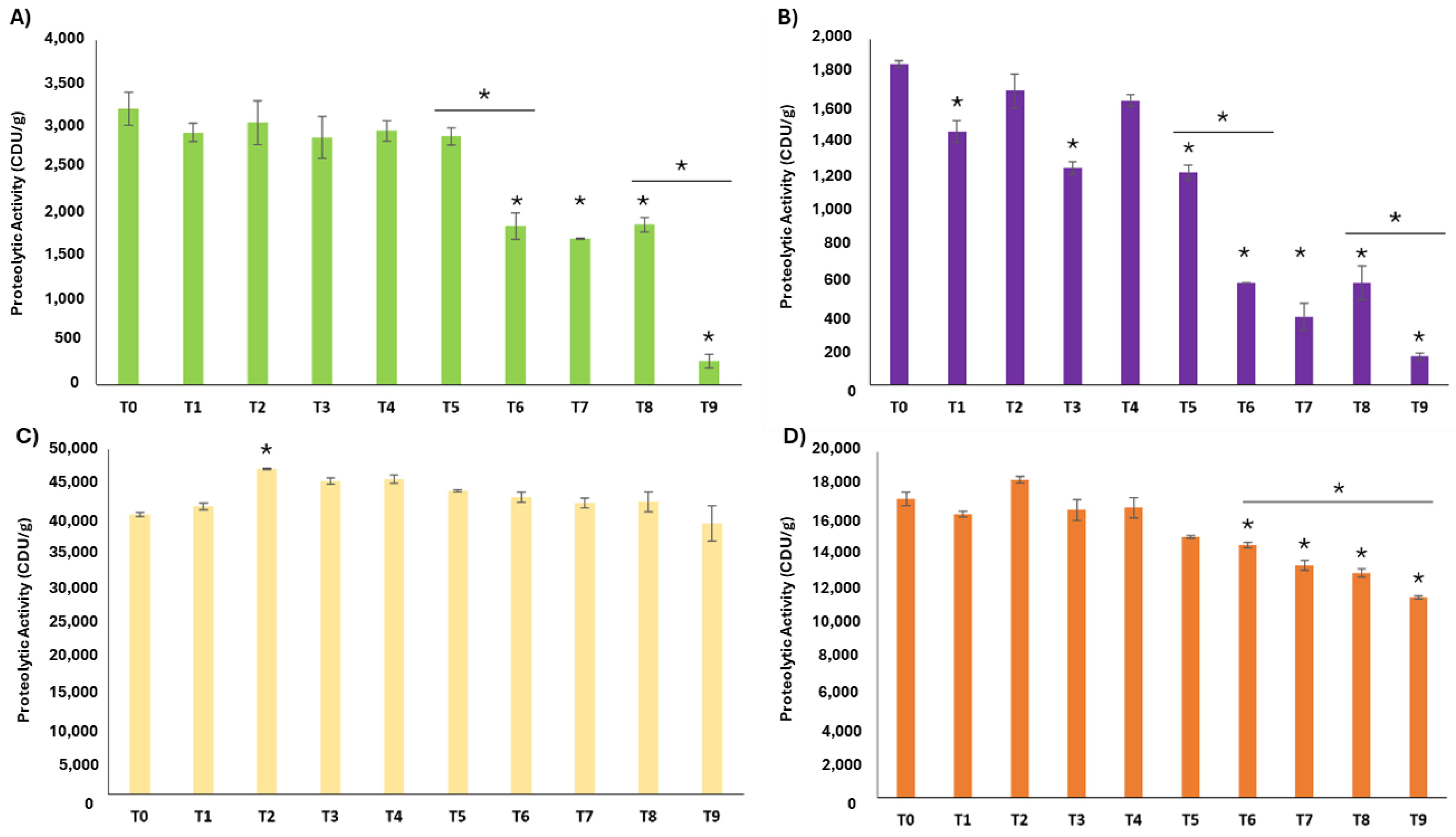
2.1.3. Enzyme Stability
2.2. Product Origin and Protein Profile Determination
2.2.1. SDS-PAGE
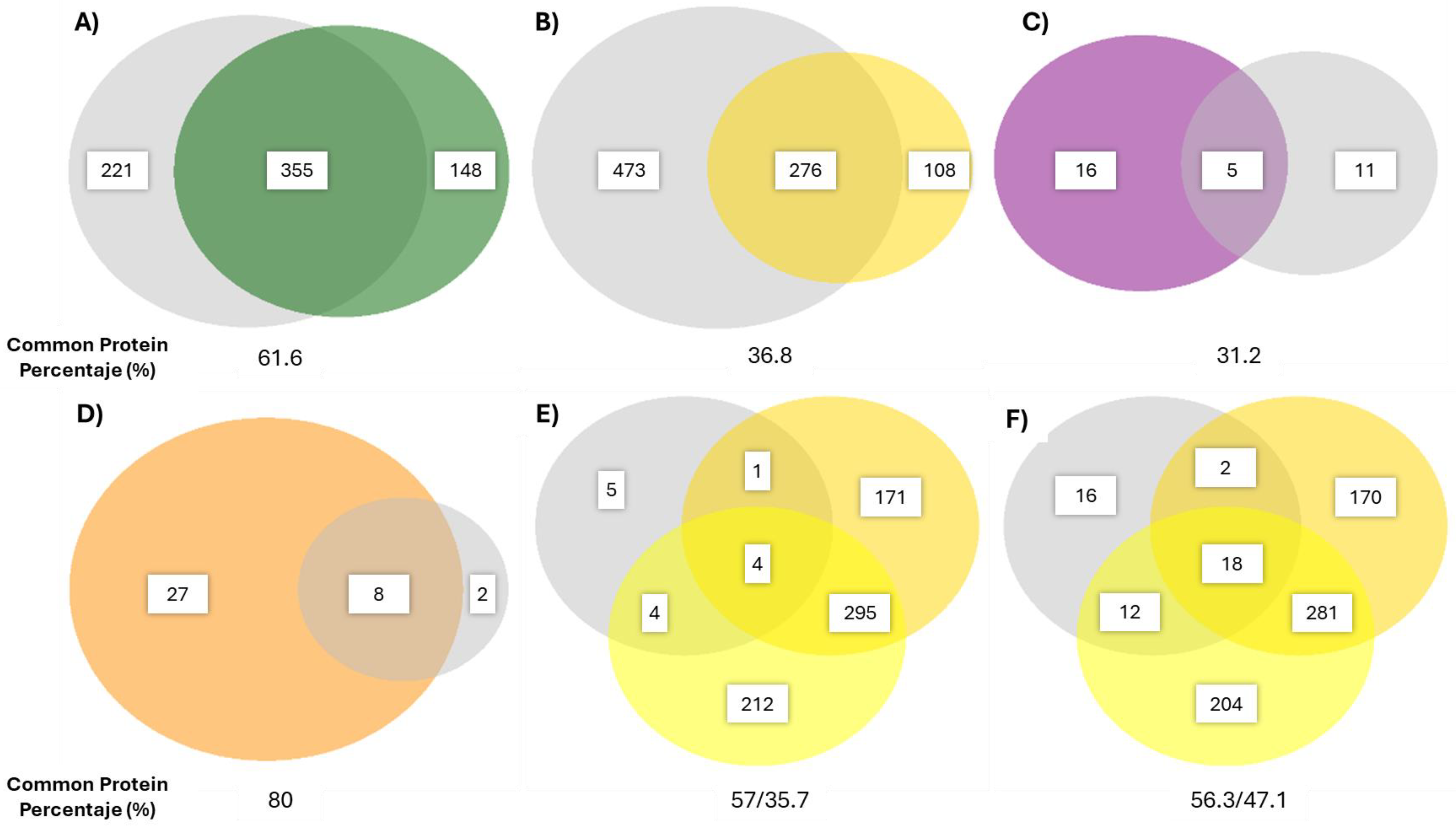
2.2.2. Protein Identification by LC-MS2-Based Proteomics
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Fruit In-House Extracts and Commercial Samples
5.2. Total Protein Quantification
5.3. Proteolytic Assays
5.3.1. Casein Assay
5.3.2. Lys-oNP Assay
5.4. Stability Assay
5.5. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
5.6. Proteomics
5.6.1. Protein Digestion
5.6.2. Reversed-Phase Liquid Chromatography–Mass Spectrometry (RP-LC–MS/MS) Analysis (Dynamic Exclusion Mode)
5.6.3. Data Processing
5.7. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cho, M.-R.; Lee, S.; Song, S.-K. A Review of Sarcopenia Pathophysiology, Diagnosis, Treatment and Future Direction. J. Korean Med. Sci. 2022, 37, e146. [Google Scholar] [CrossRef]
- Dardevet, D.; Rémond, D.; Peyron, M.-A.; Papet, I.; Savary-Auzeloux, I.; Mosoni, L. Muscle Wasting and Resistance of Muscle Anabolism: The “Anabolic Threshold Concept” for Adapted Nutritional Strategies during Sarcopenia. Sci. World J. 2012, 2012, 1–6. [Google Scholar] [CrossRef]
- Prokopidis, K.; Giannos, P.; Kirwan, R.; Ispoglou, T.; Galli, F.; Witard, O.C.; Triantafyllidis, K.K.; Kechagias, K.S.; Morwani-Mangnani, J.; Ticinesi, A.; et al. Impact of probiotics on muscle mass, muscle strength and lean mass: A systematic review and meta-analysis of randomized controlled trials. J. Cachex-Sarcopenia Muscle 2022, 14, 30–44. [Google Scholar] [CrossRef]
- Oben, J.; Kothari, S.C.; Anderson, M.L. An open label study to determine the effects of an oral proteolytic enzyme system on whey protein concentrate metabolism in healthy males. J. Int. Soc. Sports Nutr. 2008, 5, 10. [Google Scholar] [CrossRef]
- Jadhav, S.B.; Gaonkar, T.; Rathi, A. In vitro gastrointestinal digestion of proteins in the presence of enzyme supplements: Details of antioxidant and antidiabetic properties. LWT 2021, 147, 111650. [Google Scholar] [CrossRef]
- Miller, P.C.; Bailey, S.P.; Barnes, M.E.; Derr, S.J.; Hall, E.E. The effects of protease supplementation on skeletal muscle function and DOMS following downhill running. J. Sports Sci. 2004, 22, 365–372. [Google Scholar] [CrossRef]
- Matter, I.R.; Al-Omari, A.W.; Mohammed, N. Industrial Applications of Microbial Protease: A Review. Acad. Sci. J. 2023, 1, 141–160. [Google Scholar] [CrossRef]
- Akram, F.; Haq, I.U.; Jabbar, Z. Production and characterization of a novel thermo- and detergent stable keratinase from Bacillus sp. NKSP-7 with perceptible applications in leather processing and laundry industries. Int. J. Biol. Macromol. 2020, 164, 371–383. [Google Scholar] [CrossRef]
- Song, P.; Zhang, X.; Wang, S.; Xu, W.; Wang, F.; Fu, R.; Wei, F. Microbial proteases and their applications. Front. Microbiol. 2023, 14, 1236368. [Google Scholar] [CrossRef]
- Adrio, J.L.; Demain, A.L. Microbial Enzymes: Tools for Biotechnological Processes. Biomolecules 2014, 4, 117–139. [Google Scholar] [CrossRef]
- Tavano, O.L.; Berenguer-Murcia, A.; Secundo, F.; Fernandez-Lafuente, R. Biotechnological Applications of Proteases in Food Technology. Compr. Rev. Food Sci. Food Saf. 2018, 17, 412–436. [Google Scholar] [CrossRef]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and Biotechnological Aspects of Microbial Proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar] [CrossRef]
- Bawa, A.S.; Anilakumar, K.R. Genetically modified foods: Safety, risks and public concerns—A review. J. Food Sci. Technol. 2012, 50, 1035–1046. [Google Scholar] [CrossRef]
- Stanton, J.; Rezai, G.; Baglione, S. The effect of persuasive/possessing information regarding GMOs on consumer attitudes. Futur. Foods 2021, 4, 100076. [Google Scholar] [CrossRef]
- Zhang, Y.; Geary, T.; Simpson, B.K. Genetically modified food enzymes: A review. Curr. Opin. Food Sci. 2019, 25, 14–18. [Google Scholar] [CrossRef]
- Troncoso, F.D.; Sánchez, D.A.; Ferreira, M.L. Production of Plant Proteases and New Biotechnological Applications: An Updated Review. ChemistryOpen 2022, 11, e202200017. [Google Scholar] [CrossRef]
- Matkawala, F.; Nighojkar, S.; Nighojkar, A. Next-generation nutraceuticals: Bioactive peptides from plant proteases. BioTechnologia 2022, 103, 397–408. [Google Scholar] [CrossRef]
- Grzonka, Z.; Kasprzykowski, F.; Wiczk, W. Cysteine Proteases. In Industrial Enzymes; Springer: Dordrecht, The Netherlands, 2007; pp. 181–195. [Google Scholar]
- Yang, N.; Matthew, M.A.; Yao, C. Roles of Cysteine Proteases in Biology and Pathogenesis of Parasites. Microorganisms 2023, 11, 1397. [Google Scholar] [CrossRef]
- Verma, S.; Dixit, R.; Pandey, K.C. Cysteine Proteases: Modes of Activation and Future Prospects as Pharmacological Targets. Front. Pharmacol. 2016, 7, 107. [Google Scholar] [CrossRef]
- Annaházi, A.; Schröder, A.; Schemann, M. Region-specific effects of the cysteine protease papain on gastric motility. Neurogastroenterol. Motil. 2021, 33, e14105. [Google Scholar] [CrossRef]
- Kang, Y.-M.; Kang, H.-A.; Cominguez, D.C.; Kim, S.-H.; An, H.-J. Papain Ameliorates Lipid Accumulation and Inflammation in High-Fat Diet-Induced Obesity Mice and 3T3-L1 Adipocytes via AMPK Activation. Int. J. Mol. Sci. 2021, 22, 9885. [Google Scholar] [CrossRef]
- Varilla, C.; Marcone, M.; Paiva, L.; Baptista, J. Bromelain, a Group of Pineapple Proteolytic Complex Enzymes (Ananas comosus) and Their Possible Therapeutic and Clinical Effects. A Summary. Foods 2021, 10, 2249. [Google Scholar] [CrossRef]
- Maurer, H.R. Bromelain: Biochemistry, pharmacology and medical use. Cell. Mol. Life Sci. CMLS 2001, 58, 1234–1245. [Google Scholar] [CrossRef]
- Azmi, S.I.M.; Kumar, P.; Sharma, N.; Sazili, A.Q.; Lee, S.-J.; Ismail-Fitry, M.R. Application of Plant Proteases in Meat Tenderization: Recent Trends and Future Prospects. Foods 2023, 12, 1336. [Google Scholar] [CrossRef]
- Madhusankha, G.; Thilakarathna, R. Meat tenderization mechanism and the impact of plant exogenous proteases: A review. Arab. J. Chem. 2020, 14, 102967. [Google Scholar] [CrossRef]
- Anaduaka, E.G.; Chibuogwu, C.C.; Ezugwu, A.L.; Ezeorba, T.P.C. Nature-derived ingredients as sustainable alternatives for tenderizing meat and meat products: An updated review. Food Biotechnol. 2023, 37, 136–165. [Google Scholar] [CrossRef]
- Ansell, J.; Butts, C.A.; Paturi, G.; Eady, S.L.; Wallace, A.J.; Hedderley, D.; Gearry, R.B. Kiwifruit-derived supplements increase stool frequency in healthy adults: A randomized, double-blind, placebo-controlled study. Nutr. Res. 2015, 35, 401–408. [Google Scholar] [CrossRef]
- Kaur, L.; Rutherfurd, S.M.; Moughan, P.J.; Drummond, L.; Boland, M.J. Actinidin Enhances Gastric Protein Digestion As Assessed Using an in Vitro Gastric Digestion Model. J. Agric. Food Chem. 2010, 58, 5068–5073. [Google Scholar] [CrossRef]
- Montoya, C.A.; Rutherfurd, S.M.; Olson, T.D.; Purba, A.S.; Drummond, L.N.; Boland, M.J.; Moughan, P.J. Actinidin from kiwifruit (Actinidia deliciosa cv. Hayward) increases the digestion and rate of gastric emptying of meat proteins in the growing pig. Br. J. Nutr. 2013, 111, 957–967. [Google Scholar] [CrossRef]
- Christensen, M.; Tørngren, M.A.; Gunvig, A.; Rozlosnik, N.; Lametsch, R.; Karlsson, A.H.; Ertbjerg, P. Injection of marinade with actinidin increases tenderness of porcine M. biceps femoris and affects myofibrils and connective tissue. J. Sci. Food Agric. 2009, 89, 1607–1614. [Google Scholar] [CrossRef]
- Katsaros, G.I.; Tavantzis, G.; Taoukis, P.S. Production of novel dairy products using actinidin and high pressure as enzyme activity regulator. Innov. Food Sci. Emerg. Technol. 2010, 11, 47–51. [Google Scholar] [CrossRef]
- Katsaros, G.; Katapodis, P.; Taoukis, P. Modeling the effect of temperature and high hydrostatic pressure on the proteolytic activity of kiwi fruit juice. J. Food Eng. 2009, 94, 40–45. [Google Scholar] [CrossRef]
- Paul, W.; Amiss, J.; Try, R.; Praekelt, U.; Scott, R.; Smith, H. Correct Processing of the Kiwifruit Protease Actinidin in Transgenic Tobacco Requires the Presence of the C-Terminal Propeptide. Plant Physiol. 1995, 108, 261–268. [Google Scholar] [CrossRef]
- Tello-Solís, S.R.; Valle-Guadarrama, M.E.; Hernández-Arana, A. Purification and circular dichroism studies of multiple forms of actinidin from Actinidia chinensis (kiwifruit). Plant Sci. 1995, 106, 227–232. [Google Scholar] [CrossRef]
- Amid, A.; Ismail, N.A.; Yusof, F.; Salleh, H.M. Expression, purification, and characterization of a recombinant stem bromelain from Ananas comosus. Process. Biochem. 2011, 46, 2232–2239. [Google Scholar] [CrossRef]
- Ha, M.; Bekhit, A.E.-D.A.; Carne, A.; Hopkins, D.L. Characterisation of commercial papain, bromelain, actinidin and zingibain protease preparations and their activities toward meat proteins. Food Chem. 2012, 134, 95–105. [Google Scholar] [CrossRef]
- Fissore, A.; Marengo, M.; Santoro, V.; Grillo, G.; Oliaro-Bosso, S.; Cravotto, G.; Piaz, F.D.; Adinolfi, S. Extraction and Characterization of Bromelain from Pineapple Core: A Strategy for Pineapple Waste Valorization. Processes 2023, 11, 2064. [Google Scholar] [CrossRef]
- Milošević, J.; Janković, B.; Prodanović, R.; Polović, N. Comparative stability of ficin and papain in acidic conditions and the presence of ethanol. Amino Acids 2019, 51, 829–838. [Google Scholar] [CrossRef]
- Baeyens-Volant, D.; Matagne, A.; El Mahyaoui, R.; Wattiez, R.; Azarkan, M. A novel form of ficin from Ficus carica latex: Purification and characterization. Phytochemistry 2015, 117, 154–167. [Google Scholar] [CrossRef]
- Muharram, M.M.; Abdel-Kader, M.S. Utilization of gel electrophoreses for the quantitative estimation of digestive enzyme papain. Saudi Pharm. J. 2017, 25, 359–364. [Google Scholar] [CrossRef]
- Gong, X.; Morton, J.D.; Bhat, Z.F.; Mason, S.L.; Bekhit, A.E.A. Comparative efficacy of actinidin from green and gold kiwi fruit extract on in vitro simulated protein digestion of beef Semitendinosus and its myofibrillar protein fraction. Int. J. Food Sci. Technol. 2019, 55, 742–750. [Google Scholar] [CrossRef]
- Pezzani, R.; Jiménez-Garcia, M.; Capó, X.; Gürer, E.S.; Sharopov, F.; Rachel, T.Y.L.; Woutouoba, D.N.; Rescigno, A.; Peddio, S.; Zucca, P.; et al. Anticancer properties of bromelain: State-of-the-art and recent trends. Front. Oncol. 2023, 12, 1068778. [Google Scholar] [CrossRef]
- Tuppo, L.; Giangrieco, I.; Palazzo, P.; Bernardi, M.L.; Scala, E.; Carratore, V.; Tamburrini, M.; Mari, A.; Ciardiello, M.A. Kiwellin, a Modular Protein from Green and Gold Kiwi Fruits: Evidence of in Vivo and in Vitro Processing and IgE Binding. J. Agric. Food Chem. 2008, 56, 3812–3817. [Google Scholar] [CrossRef]
- Napper, A.D.; Bennett, S.P.; Borowski, M.; Holdridge, M.B.; Leonard, M.J.; E Rogers, E.; Duan, Y.; A Laursen, R.; Reinhold, B.; Shames, S.L. Purification and characterization of multiple forms of the pineapple-stem-derived cysteine proteinases ananain and comosain. Biochem. J. 1994, 301, 727–735. [Google Scholar] [CrossRef]
- Sperber, A.D.; Bangdiwala, S.I.; Drossman, D.A.; Ghoshal, U.C.; Simren, M.; Tack, J.; Whitehead, W.E.; Dumitrascu, D.L.; Fang, X.; Fukudo, S.; et al. Worldwide Prevalence and Burden of Functional Gastrointestinal Disorders, Results of Rome Foundation Global Study. Gastroenterology 2021, 160, 99–114. [Google Scholar] [CrossRef]
- Benito-Vázquez, I.; Garrido-Romero, M.; Díez-Municio, M.; Moreno, F.J. Cysteine Proteases from Plants: Uti-lization in Foods and Nutraceuticals and Characterization Approaches for Quality Management. Appl. Food Res. 2024, 4, 100491. [Google Scholar] [CrossRef]
- Corzo, C.A.; Waliszewski, K.N.; Welti-Chanes, J. Pineapple fruit bromelain affinity to different protein substrates. Food Chem. 2011, 133, 631–635. [Google Scholar] [CrossRef]
- Coêlho, D.F.; Saturnino, T.P.; Fernandes, F.F.; Mazzola, P.G.; Silveira, E.; Tambourgi, E.B. Azocasein Substrate for Determination of Proteolytic Activity: Reexamining a Traditional Method Using Bromelain Samples. BioMed Res. Int. 2016, 2016, 1–6. [Google Scholar] [CrossRef]
- Vallés, D.; Cantera, A.M. Antiacanthain A: New proteases isolated from Bromelia antiacantha Bertol. (Bromeliaceae). Int. J. Biol. Macromol. 2018, 113, 916–923. [Google Scholar] [CrossRef]
- Charney, J.; Tomarelli, R.M. A Colorimetric Method for the Determination of the Proteolytic Activity of Duodenal Juice. J. Biol. Chem. 1947, 171, 501–505. [Google Scholar] [CrossRef]
- Devakate, R.; Patil, V.; Waje, S.; Thorat, B. Purification and drying of bromelain. Sep. Purif. Technol. 2008, 64, 259–264. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, B.; Yan, Q.-J.; Jiang, Z.-Q. Comparative analysis on the distribution of protease activities among fruits and vegetable resources. Food Chem. 2016, 213, 708–713. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Yamashita, Y.; Takeda, I.; Kiso, H. Proteolytic Enzymes in Green Asparagus, Kiwi Fruit and Miut: Occurrence and Partial Characterization. Agric. Biol. Chem. 1982, 46, 1983–1986. [Google Scholar] [CrossRef]
- Campos, D.A.; Coscueta, E.R.; Valetti, N.W.; Pastrana-Castro, L.M.; Teixeira, J.A.; Picó, G.A.; Pintado, M.M. Optimization of bromelain isolation from pineapple byproducts by polysaccharide complex formation. Food Hydrocoll. 2018, 87, 792–804. [Google Scholar] [CrossRef]
- Chalabi, M.; Khademi, F.; Yarani, R.; Mostafaie, A. Proteolytic Activities of Kiwifruit Actinidin (Actinidia deliciosa cv. Hayward) on Different Fibrous and Globular Proteins: A Comparative Study of Actinidin with Papain. Appl. Biochem. Biotechnol. 2014, 172, 4025–4037. [Google Scholar] [CrossRef]
- Yow, A.G.; Bostan, H.; Young, R.; Valacchi, G.; Gillitt, N.; Perkins-Veazie, P.; Xiang, Q.-Y.; Iorizzo, M. Identification of bromelain subfamily proteases encoded in the pineapple genome. Sci. Rep. 2023, 13, 11605. [Google Scholar] [CrossRef]
- Calişkan, G.; Ergün, K.; Dirim, N. Freeze Drying of Kiwi (Actinidia Deliciosa) Puree and the Powder Proper-ties. Ital. J. Food Sci. 2015, 27, 385–396. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kunitz, M. Crystalline Soybean Trypsin Inhibitor. J. Gen. Physiol. 1947, 30, 291–310. [Google Scholar] [CrossRef]
- Dhiman, V.K.; Chauhan, V.; Kanwar, S.S.; Singh, D.; Pandey, H. Purification and characterization of actinidin from Actinidia deliciosa and its utilization in inactivation of α-amylase. Bull. Natl. Res. Cent. 2021, 45, 213. [Google Scholar] [CrossRef]
- Baker, E.N.; Boland, M.J.; Calder, P.C.; Hardman, M.J. The specificity of actinidin and its relationship to the structure of the enzyme. Biochim. Et Biophys. Acta BBA-Enzym. 1980, 616, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Boland, M.; Hardman, M. Kinetic studies on the thiol protease from Actinidia chinensis. FEBS Lett. 1972, 27, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Pisa, D.; Marina, A.I.; Morato, E.; Rábano, A.; Rodal, I.; Carrasco, L. Evidence for Fungal Infection in Cerebrospinal Fluid and Brain Tissue from Patients with Amyotrophic Lateral Sclerosis. Int. J. Biol. Sci. 2015, 11, 546–558. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Sample | Total Identified Proteins ** | Proteolytic Enzymes *** | Percentage Ratio of Proteolytic Enzymes vs Total Identified Proteins |
|---|---|---|---|
| Kiwi Green * | 503 | 11 | 2.2 |
| K2 | 576 | 11 | 2 |
| Kiwi Gold * | 384 | 10 | 2.6 |
| K5 | 749 | 10 | 1.3 |
| Pineapple core * | 471 | 10 | 2.1 |
| Pineapple pulp * | 515 | 14 | 2.7 |
| P5 | 48 | 13 | 27 |
| P4 | 14 | 6 | 42.9 |
| Papaya * | 35 | 6 | 17.1 |
| PAP2 | 10 | 3 | 70 |
| Fig * | 21 | 8 | 38 |
| F2 | 16 | 1 | 6.25 |
| Fruit | Database Entry (Uniprot) | −10log(P) a | Coverage (%) b | Protein Identification | Theoretical Mw (kDa) Based on Amino Acid or Nucleotide Sequence | Mr (kDa) SDS-PAGE |
|---|---|---|---|---|---|---|
| Kiwi Green/K2 | P00785|ACTN_ACTCC ** | 455.00485/471.76282 | 42.4/53.4 | Actinidin | 27.5 | 26.5• |
| A0A2R6QY36|A0A2R6QY36_ACTCC ** | 384.40567/420.12158 | 40.2/52.4 | Actinidin-like | 23.3 | 23.3• | |
| P83958|TLP_ACTCC ** | 383.77435/378.04047 | 52.9/52.9 | Thaumatin-like protein | 21.7 | ||
| P85261|KIWEL_ACTCC ** | 452.24118/467.28903 | 77.9/74.2 | Kiwellin | 19.8 | ||
| A0A2R6PEY1|A0A2R6PEY1_ACTCC ** | 290.15417/304.33725 | 46.3/37.9 | KiTH-2 like | 15.5 | 17.4•/14.0• | |
| A0A2R6PAW0|A0A2R6PAW0_ACTCC ** | 390.88022/391.0907 | 70.7/85.3 | Kirola | 17.2 | ||
| Kiwi Gold/K5 | P00785|ACTN_ACTCC ** | 455.00485/399.8263 | 42.4/32.4 | Actinidin | 27.5 | |
| A0A2R6QY36|A0A2R6QY36_ACTCC ** | 384.40567/494.2946 | 40.2/62.2 | Actinidin-like | 23.3 | 22.8• | |
| P83958|TLP_ACTCC ** | 383.77435/268.69028 | 52.9/32.4 | Thaumatin-like protein | 21.7 | 20.3• | |
| P85261|KIWEL_ACTCC ** | 452.24118/504.32312 | 77.9/79.3 | Kiwellin | 19.8 | ||
| A0A2R6PEY1|A0A2R6PEY1_ACTCC ** | 290.15417/289.5369 | 46.2/38.7 | KiTH-2 like | 15.5 | 13.4• | |
| A0A2R6PAW0|A0A2R6PAW0_ACTCC ** | 390.88022/432.87717 | 70.7/84 | Kirola | 17.2 | ||
| Fig/F2 | A0A2Z6DRL4|A0A2Z6DRL4_FICCA | 457.78674 | 59 | Ficin 1a | 23.6 | 22.9 †/25.8 ‡ |
| A0A2Z6DRT1|A0A2Z6DRT1_FICCA | 419.7921 | 57.6 | Ficin 1b | 23.6 | ||
| A0A2Z6DRN1|A0A2Z6DRN1_FICCA | 391.46454 | 36.1 | Ficin 3 | 23.7 | ||
| A0A2Z6DRM5|A0A2Z6DRM5_FICCA | 245.10976 | 15.1 | Ficin 2c | 23.3 | ||
| A0A2Z6DRL9|A0A2Z6DRL9_FICCA | 245.10976 | 15.1 | Ficin 2a | 23.3 | ||
| A0A2Z6DRP9|A0A2Z6DRP9_FICCA | 245.10976 | 15.1 | Ficin 2b | 23.3 | ||
| A0A182DW11|A0A182DW11_FICCA | 239.45866 | 22.3 | Ficin isoform D | 23.4 | ||
| A0A2Z6DRL5|A0A2Z6DRL5_FICCA** | 448.26428/292.24368 | 51.1/34.7 | Ficin 1c | 23.6 | ||
| Papaya/PAP2 | P14080|PAPA2_CARPA | 367.02426 | 48 | Chymopapain | 23.7 | 23.8 ‡ |
| P05994|PAPA4_CARPA ** | 331.4425/ 363.13953 | 46.8/49.4 | Papaya proteinase 4 | 23.3 | ||
| P10056|PAPA3_CARPA ** | 271.31442/341.473 | 37.6/45.4 | Caricain | 23.4 | ||
| H6USN0|H6USN0_CARPA | 264.59772 | 25 | Cysteine protease | ND | ||
| P00784|PAPA1_CARPA ** | 216.544/298.2124 | 28.4/43.2 | Papain | 23.4 | ||
| H6USN1|H6USN1_CARPA | 205.06474 | 16.3 | Cysteine protease | 23.4 | ||
| Pineapple core/pulp/P4/P5 | A0A199W8R4|A0A199W8R4_ANACO ** (P4) | 392.8472/125.80339/472.0772 | 18.9/5.9/29.7 | Fruit bromelain (core) | ND | 26.0•/23.0• (core and pulp, and P5)/21.8 ‡/29.7 ‡ (P4) |
| A0A199VSS3|A0A199VSS3_ANACO ** (P4 and P5) | 339.10794/457.8168/228.36128/526.3434 | 32.9/47.5/33.7/54.5 | Ananain (core and pulp) | 22.9 | ||
| A0A199UL32|A0A199UL32_ANACO | 330.18286/ 428.22952 | 21.8/24.8 | Ananain (core and pulp) | ND | ||
| A0A199W9F2|A0A199W9F2_ANACO ** (P4 and P5) | 294.8118/452.54492/186.53676/540.68054 | 7.2/19.2/8.3/24.5 | Fruit bromelain (core and pulp) | ND | ||
| O24641|O24641_ANACO | 267.19427/285.8756 | 11.1/14.2 | Bromelain (core and pulp) | 22.9 | ||
| A0A199W9E5|A0A199W9E5_ANACO ** (P4) | 267.19427/285.8756/325.2002 | 11.1/14.3/15.7 | Fruit bromelain (core and pulp) | 22.9 | ||
| A0A199UUY2|A0A199UUY2_ANACO ** (P4 and P5) | 435.98737/164.08255/536.8819 | 40.7/24.5/68.3 | Ananain (pulp) | ND | ||
| A0A199W8N4|A0A199W8N4_ANACO ** (P4 and P5) | 412.75708/177.17471/451.46368 | 30.2/20.4/34.1 | Ananain (pulp) | ND | ||
| A0A199UKV8|A0A199UKV8_ANACO ** (P4) | 361.48822/434.87213 | 24.4/38.2 | Ananain (pulp) | 23.2 | ||
| A0A199W3D2|A0A199W3D2_ANACO ** (P4 and P5) | 425.63855/199.6339/534.271 | 14.1/11.1/23.1 | Fruit bromelain (pulp) | ND | ||
| A0A199VJ20|A0A199VJ20_ANACO * (P5) | 325.2002 | 15.7 | Fruit bromelain | ND | ||
| A0A199UVV3|A0A199UVV3_ANACO * (P5) | 158.96414 | 3.7 | Fruit bromelain | 21.9 |
| Sample | Fruit | Code * |
|---|---|---|
| Kiwi Green commercial products | Actinidia deliciosa (Kiwi Green) | K2, K3, K4, and K6 |
| Kiwi Gold commercial products | Actinidia chinensis (Kiwi Gold) | K5 |
| Papaya commercial products | Carica papaya (Papaya) | PAP1 and PAP2 |
| Pineapple commercial products | Ananas comosus (Pineapple) | P2, P4, P5, and P6 |
| Fig commercial product | Ficus carica (Fig) | F2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benito-Vázquez, I.; Muñoz-Labrador, A.; Garrido-Romero, M.; Hontoria-Caballo, G.; García-García, C.; Diez-Municio, M.; Moreno, F.J. New Pipeline for Analysing Fruit Proteolytic Products Used as Digestive Health Nutraceuticals. Int. J. Mol. Sci. 2024, 25, 10315. https://doi.org/10.3390/ijms251910315
Benito-Vázquez I, Muñoz-Labrador A, Garrido-Romero M, Hontoria-Caballo G, García-García C, Diez-Municio M, Moreno FJ. New Pipeline for Analysing Fruit Proteolytic Products Used as Digestive Health Nutraceuticals. International Journal of Molecular Sciences. 2024; 25(19):10315. https://doi.org/10.3390/ijms251910315
Chicago/Turabian StyleBenito-Vázquez, Iván, Ana Muñoz-Labrador, Manuel Garrido-Romero, Gema Hontoria-Caballo, Carlos García-García, Marina Diez-Municio, and F. Javier Moreno. 2024. "New Pipeline for Analysing Fruit Proteolytic Products Used as Digestive Health Nutraceuticals" International Journal of Molecular Sciences 25, no. 19: 10315. https://doi.org/10.3390/ijms251910315
APA StyleBenito-Vázquez, I., Muñoz-Labrador, A., Garrido-Romero, M., Hontoria-Caballo, G., García-García, C., Diez-Municio, M., & Moreno, F. J. (2024). New Pipeline for Analysing Fruit Proteolytic Products Used as Digestive Health Nutraceuticals. International Journal of Molecular Sciences, 25(19), 10315. https://doi.org/10.3390/ijms251910315