
Molecular Markers and Marker-Assisted Selection Provide Genetic Insights for Identifying Key Quantitative Trait Locus for Watermelon Rind Thickness

Abstract
:1. Introduction
2. Results
2.1. Genetic and Phenotypic Characteristics of Watermelon RTs
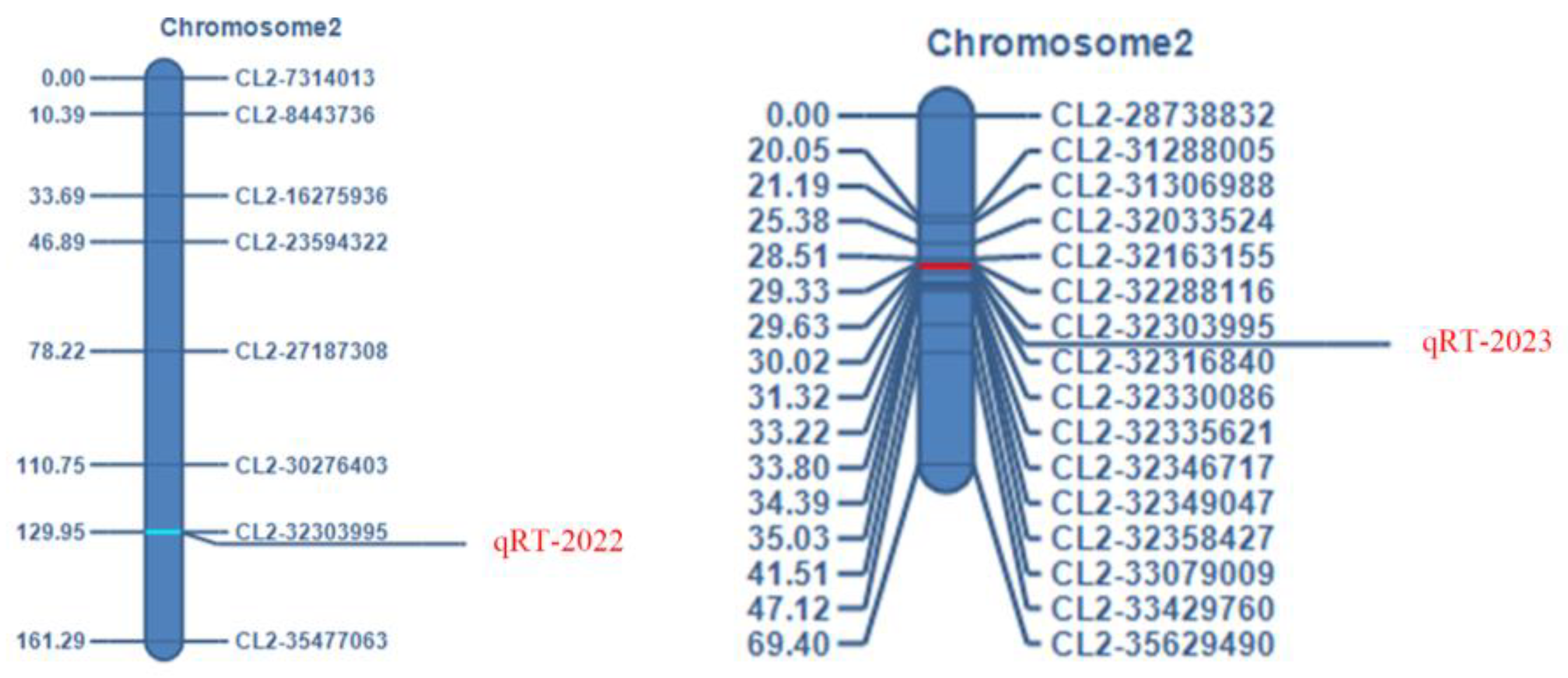
2.2. Genetic Linkage Analysis Placed the RT Locus into a 12.845 kb Region on Chromosome 2
2.3. Analysis of the Candidate Gene Cla97C02G044120
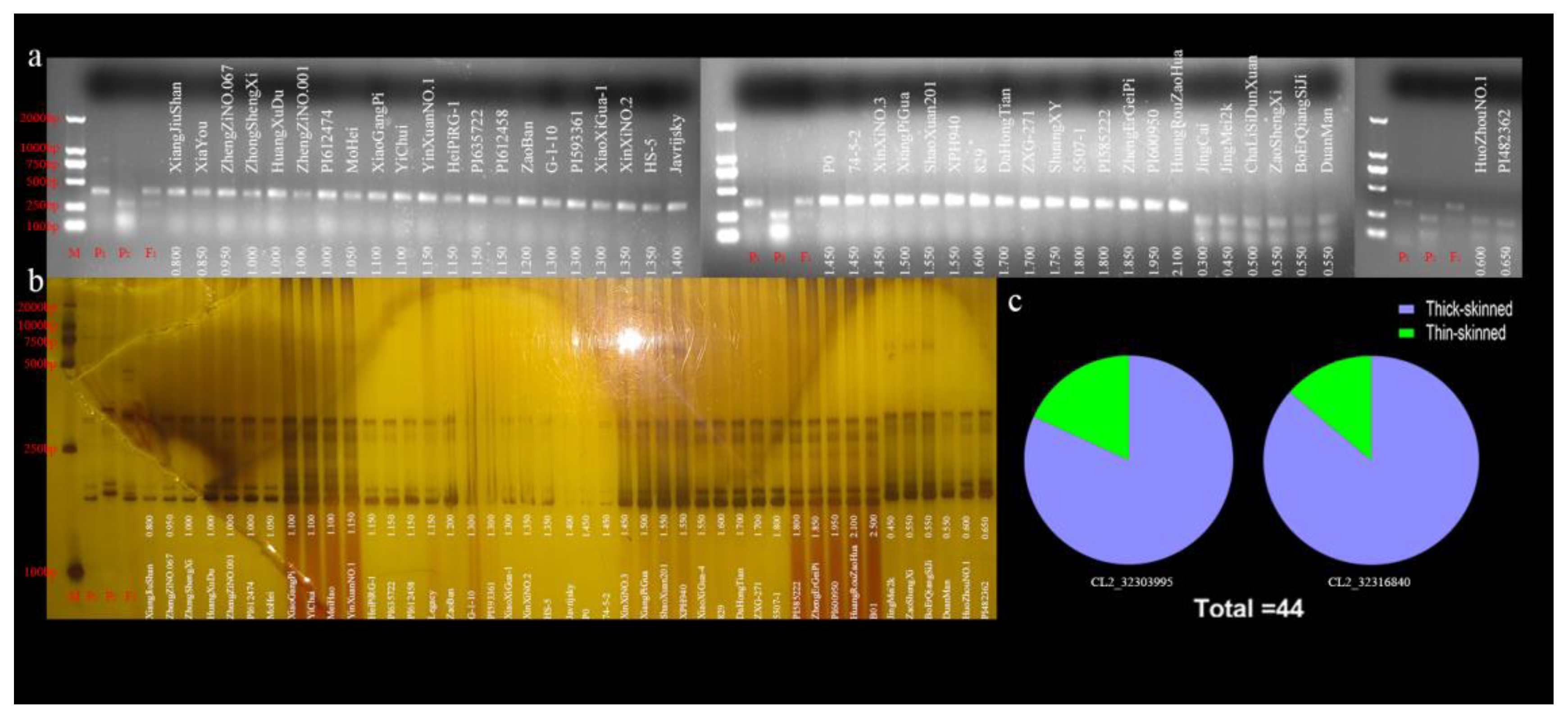
2.4. Marker-Assisted Selection (MAS) of the RT Trait in a Natural Watermelon Panel
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Genetic Mapping Population
4.2. DNA and RNA Extraction
4.3. Molecular Marker Development and Genetic Mapping
4.4. Candidate Gene Prediction and Analysis with MAS System Verification
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Liu, Y.; Chen, K.; Zhao, Z.; Sun, L.; Gao, Z.; Zhu, Z.; Han, W.; Luan, F. Mixed major gene and polygene inheritance analysis of single fruit weight in watermelon. J. Northeast. Agric. Univ. 2022, 53, 9–15. [Google Scholar] [CrossRef]
- Sun, H.; Zeng, J.; Ou, L.; Zou, X. Research Progress of Pericarp. Chin. Agric. Sci. Bull. 2021, 37, 49–53. [Google Scholar] [CrossRef]
- Yang, T. QTL Analysis of Important Characters related to Watermelon Rind Hardness. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2021. [Google Scholar] [CrossRef]
- Gong, C.; Wang, S.; Liu, J.; Pan, B.; Guo, G.; Diao, W. Research Progress on the Development and Regulation of Rind Thickness in Cucurbit Vegetables. J. China Capsicum 2022, 20, 7–13. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.; Chen, K.; Lu, B.; Zhang, Y.; Wu, P.; Luan, F. Mixed major gene and polygene inheritance analysis of fruit hardness in Citrullus lanatus. J. Northeast. Agric. Univ. 2016, 47, 24–32. [Google Scholar] [CrossRef]
- Gong, C.; Lu, X.; Zhu, H.; Anees, M.; He, N.; Liu, W.G. Genome-wide association study provides genetic insights into natural variation in watermelon rind thickness and single fruit weight. Front. Plant Sci. 2022, 13, 1074145. [Google Scholar] [CrossRef]
- Fan, M.; Xu, Y.; Zhang, H.; Ren, H.; Kang, G.; Wang, Y.; Chen, H. Identification of Quantitative Trait Loci Associated with Fruit Traits in Watermelon [Citullus lanantus (Thanb) Mansf] and Analysis of Their Genetic Effects. Acta genetica Sinica. 2000, 10, 902–910. [Google Scholar]
- Cheng, Y. Construction of Genetic Linkage Map and QTL Analysis for Fruit-Related Traits in Watermelon. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2015. [Google Scholar]
- Sandlin, K.; Prothro, J.; Heesacker, A.; Khalilian, N.; Okashah, R.; Xiang, W.; Bachlava, E.; Caldwell, D.G.; Taylor, C.A.; Seymour, D.K.; et al. Comparative mapping in watermelon [Citrullus lanatus (Thunb.) Matsum. et Nakai]. Theor. Appl. Genet. 2012, 125, 1603–1618. [Google Scholar] [CrossRef]
- Ren, Y.; McGregor, C.; Zhang, Y.; Gong, G.; Zhang, H.; Guo, S.; Sun, H.; Cai, W.; Zhang, J.; Xu, Y. An integrated genetic map based on four mapping populations and quantitative trait loci associated with economically important traits in watermelon (Citrullus lanatus). BMC Plant Biol. 2014, 14, 33. [Google Scholar] [CrossRef]
- Kim, M.; Nguyen, T.T.P.; Ahn, J.H.; Kim, G.J.; Sim, S.C. Genome-wide association study identifies QTL for eight fruit traits in cultivated tomato (Solanum lycopersicum L.). Hortic. Res. 2021, 8, 203. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhou, Y.; Li, J.; Yu, T.; Wu, T.; Luo, J.; Luo, S.; Huang, H. A high-density linkage map and QTL mapping of fruit-related traits in pumpkin (Cucurbita moschata Duch.). Sci. Rep. 2017, 7, 12785. [Google Scholar] [CrossRef]
- Xie, D.; Xu, Y.; Wang, J.; Liu, W.; Zhou, Q.; Luo, S.; Huang, W.; He, X.; Li, Q.; Peng, Q.; et al. The wax gourd genomes offer insights into the genetic diversity and ancestral cucurbit karyotype. Nat. Commun. 2019, 10, 51–58. [Google Scholar] [CrossRef]
- Lee, H.Y.; Ro, N.Y.; Patil, A.; Lee, J.H.; Kwon, J.K.; Kang, B.C. Uncovering Candidate Genes Controlling Major Fruit-Related Traits in Pepper via Genotype-by-Sequencing Based QTL Mapping and Genome-Wide Association Study. Front. Plant Sci. 2020, 11, 1100. [Google Scholar] [CrossRef]
- Yuan, X.; Fang, R.; Zhou, K.; Lei, G.; Huang, Y.; Chen, X. Association Analysis of Important Agronomic Traits in Pepper (Capsicum annuum L.) and Mining of Elite Alleles. J. Nucl. Agric. Sci. 2020, 34, 2658–2672. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, X.; Qiu, L.; Zhai, C.; Song, X. Research Progress on Pericarp Thickness of Waxy Corn. Heilongjiang Agric. Sci. 2021, 8, 108–111. [Google Scholar]
- Jiang, F.; Liang, R.; Yan, Y.; Liang, Z.; Huang, Z.; Liu, P. Genetic Analysis and QTL Mapping of Pericarp Thickness in Sweet Corn. Guangdong Agric. Sci. 2022, 49, 21–28. [Google Scholar]
- Czerednik, A.; Busscher, M.; Angenent, G.C.; de Maagd, R.A. The cell size distribution of tomato fruit can be changed by overexpression of CDKA1. Plant Biotechnol. J. 2015, 13, 259–268. [Google Scholar] [CrossRef]
- Swinnen, G.; Mauxion, J.P.; Baekelandt, A.; De Clercq, R.; Van Doorsselaere, J.; Inzé, D.; Gonzalez, N.; Goossens, A.; Pauwels, L. SlKIX8 and SlKIX9 are negative regulators of leaf and fruit growth in tomato. Plant Physiol. 2022, 188, 382–396. [Google Scholar] [CrossRef]
- De Jong, M.; Wolters-Arts, M.; Feron, R.; Mariani, C.; Vriezen, W.H. The Solanum lycopersicum auxin response factor 7 (SlARF7) regulates auxin signaling during tomato fruit set and development. Plant J. Cell Mol. Biol. 2009, 57, 160–170. [Google Scholar] [CrossRef]
- Su, L.Y.; Bassa, C.; Audran, C.; Mila, I.; Cheniclet, C.; Chevalier, C.; Bouzayen, M.; Roustan, J.P.; Chervin, C. The auxin Sl-IAA17 transcriptional repressor controls fruit size via the regulation of endoreduplication-related cell expansion. Plant Cell Physiol. 2014, 55, 1969–1976. [Google Scholar] [CrossRef]
- Zhu, Z.; Liang, H.; Chen, G.; Li, F.; Wang, Y.; Liao, C.; Hu, Z. The bHLH transcription factor SlPRE2 regulates tomato fruit development and modulates plant response to gibberellin. Plant Cell Rep. 2019, 38, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Musseau, C.; Jorly, J.; Gadin, S.; Sørensen, I.; Deborde, C.; Bernillon, S.; Mauxion, J.P.; Atienza, I.; Moing, A.; Lemaire-Chamley, M.; et al. The Tomato Guanylate-Binding Protein SlGBP1 Enables Fruit Tissue Differentiation by Maintaining Endopolyploid Cells in a Non-Proliferative State. Plant Cell 2020, 32, 3188–3205. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, J.; Zhang, H.; Shi, S.; Liu, L.; Shu, B.; Liang, Q.; Xie, J.; Wei, Y. De novo assembly and characterization of pericarp transcriptome and identification of candidate genes mediating fruit cracking in Litchi chinensis Sonn. Int. J. Mol. Sci. 2014, 15, 17667–17685. [Google Scholar] [CrossRef]
- Landoni, M.; Puglisi, D.; Cassani, E.; Borlini, G.; Brunoldi, G.; Comaschi, C.; Pilu, R. Phlobaphenes modify pericarp thickness in maize and accumulation of the fumonisin mycotoxins. Sci. Rep. 2020, 10, 1417. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bai, L.; Du, R.; Guo, H.; Yang, Q.; Wang, P.; Xu, G. Analysis of regression and path of main yield traits of melon. J. North. Agric. 2016, 44, 37–39. [Google Scholar]
- Yang, T.; Tang, Y.; Zhang, G.; Li, X.; Patiguli; Wang, B.; Li, N.; Wang, J.; Yu, Q.; Yang, S. Correlation Analysis of the Processing Tomato Based on the Main Characteristics of the Fruit and Pulp Tissue. Xinjiang Agric. Sci. 2019, 56, 797–807. [Google Scholar]
- Yang, T.; Amanullah, S.; Li, S.; Cheng, R.; Zhang, C.; Zhao, Z.; Liu, H.; Luan, F.; Wang, X. Molecular Mapping of Putative Genomic Regions Controlling Fruit and Seed Morphology of Watermelon. Int. J. Mol. Sci. 2023, 24, 15755. [Google Scholar] [CrossRef]
- Zhan, Y.; He, H.; Xie, C.; Dang, X. Study on Fruit Mechanical Properties and Peel Structure during Development of Watermelon with Different Crack Tolerance. Guangdong Agric. Sci. 2020, 47, 18–24. [Google Scholar] [CrossRef]
- Du, Q.; Li, L.; Xu, Q.; Chen, X. Studies on the Relations between Fruit Crispness and the Structure of Pickling-type Cucumber (Cucumis sativus L.). Hubei Agric. Sci. 2012, 51, 940–942. [Google Scholar] [CrossRef]
- Gao, M.; Yuan, C.; Wang, Y. A comparative study of the microstructure of watermelon rind. Jiangsu Agric. Sci. 2013, 41, 133–135. [Google Scholar] [CrossRef]
- Liu, P.; Liu, W.; Zhao, S.; Yan, Z.; He, N.; Lu, X. Comparison on anatomic structure of fruit rind in different ploidy watermelons. J. Fruit Sci. 2012, 29, 296–301+320. [Google Scholar] [CrossRef]
- Guan, Z.; Cheng, Z. Effects of Foliage Application of CaCl2 and NaCl on Pericarp Microstructure of Processing Tomato. Acta Hortic. Sin. 2010, 37, 229–234. [Google Scholar] [CrossRef]
- Yan, S.; Yuan, H.; Lei, Y.; Zhao, L.; Luo, R.; Ni, D.; Wang, L. Micro-morphology analysis of liquor-making sorghums grain and the correlation of pericarp thickness and tannin content. China Brew. 2018, 37, 67–71. [Google Scholar]
- Wang, X.; Yang, T.; Liu, Z.; Sun, L.; Zhu, Z.; Gao, P.; Liu, S.; Luan, F. Analysis on hardness related characters of watermelon rind. J. Northeast. Agric. Univ. 2020, 51, 35–44. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, Z.; Pei, S.; Zhang, X.; Lin, S.; Luan, F. Correlation and Principal Component Analysis of Main Quantitative Characteristics in Watermelon. North. Hortic. 2017, 12, 35–39. [Google Scholar]
- Zhou, H. The Research of Phenotype Analysis and CAPS Marker for Fruit Traits in Melon. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2016. [Google Scholar]
- Li, Y.; Jin, G.; Chun, C.; Zhu, X.; Qiu, X. Developmental characteristics of citrus peel and the effect of gibberellic acid on fruit cracking. J. Fruit Sci. 2021, 38, 1092–1101. [Google Scholar] [CrossRef]
- Guo, Q.; Yu, L.; Zhao, G. Effects of Different Harvest Date on Grain Quality of Waxy Maize. Crops 2017, 2, 126–129. [Google Scholar] [CrossRef]
- Yue, S.; Wang, X.; Zhang, B.; Wu, X. Carbohydrate content and pericarp thickness of kernels at milk maturity of fruit and vegetable-type glutinous maize and common maize. Guangdong Agric. Sci. 2004, 6, 40–42. [Google Scholar] [CrossRef]
- Yang, T.; Amanullah, S.; Pan, J.; Chen, G.; Liu, S.; Ma, S.; Wang, J.; Gao, P.; Wang, X. Identification of putative genetic regions for watermelon rind hardness and related traits by BSA-seq and QTL mapping. Euphytica 2021, 217, 19. [Google Scholar] [CrossRef]
- Liao, N.; Hu, Z.; Li, Y.; Hao, J.; Chen, S.; Xue, Q.; Ma, Y.; Zhang, K.; Mahmoud, A.; Ali, A.; et al. Ethylene-responsive factor 4 is associated with the desirable rind hardness trait conferring cracking resistance in fresh fruits of watermelon. Plant Biotechnol. J. 2020, 18, 1066–1077. [Google Scholar] [CrossRef]
- Nascimento, D.L.T.; Souza, F.D.F.; Dias, S.C.D.R.; Santos, D.S.J.; Melo, D.F.N. Inheritance of seed size in watermelon populations. Rev. Colomb. De Cienc. Hortícolas 2020, 14, 9–16. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Gao, L. Transcriptome Analysis and Fine Mapping of Major Genes Controlling Flesh Firmness and Sour Flesh in Watermelon. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Zhengzhou, China, 2018. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Description | Position |
|---|---|---|
| Cla97C02G043940 | Ethylene-responsive transcription factor 3-like | 32161415 .. 32162074 (+) |
| Cla97C02G043950 | Triacylglycerol lipase 2, putative | 32171738 .. 32173760 (+) |
| Cla97C02G043960 | Lipase | 32176692 .. 32179249 (+) |
| Cla97C02G043970 | Protein disulfide isomerase-like 2-3 | 32182402 .. 32186689 (+) |
| Cla97C02G043980 | Centromere/kinetochore protein zw10 homolog | 32188755 .. 32196057 (−) |
| Cla97C02G043990 | Lipid-A-disaccharide synthase | 32202021 .. 32205359 (+) |
| Cla97C02G044000 | Unknown protein | 32206780 .. 32207294 (+) |
| Cla97C02G044010 | Unknown protein | 32208194 .. 32208541 (+) |
| Cla97C02G044020 | Ubiquitin and WLM domain-containing protein | 32218483 .. 32221679 (+) |
| Cla97C02G044030 | 3-oxoacyl-[acyl-carrier-protein] synthase | 32223103 .. 32229573 (+) |
| Cla97C02G044040 | F-box family protein | 32234318 .. 32236622 (−) |
| Cla97C02G044050 | Heat stress transcription factor A-4c-like | 32238954 .. 32239963 (−) |
| Cla97C02G044060 | Transcription elongation factor B polypeptide 3-like | 32246890 .. 32248741 (+) |
| Cla97C02G044070 | Citrate synthase | 32252262 .. 32256301 (−) |
| Cla97C02G044080 | Ubiquinone biosynthesis O-methyltransferase, mitochondrial | 32264788 .. 32269245 (−) |
| Cla97C02G044090 | Leucine-rich repeat receptor-like protein kinase family protein | 32278703 .. 32280043 (−) |
| Cla97C02G044100 | Serine/threonine protein phosphatase 7 long form | 32283295 .. 32286529 (−) |
| Cla97C02G044110 | Serine/threonine-protein phosphatase | 32290370 .. 32300164 (−) |
| Cla97C02G044120 | Elongation factor 2 | 32305922 .. 32314246 (+) |
| Cla97C02G044130 | Protein kinase, putative | 32320831 .. 32328837 (+) |
| Cla97C02G044140 | Elongation factor 2 | 32332095 .. 32335291 (+) |
| Gene ID | Position | P1 | P2 | Name | Organism | Function |
|---|---|---|---|---|---|---|
| Cla97C02G043940 | 1112 | A * | T | CAAT-box | Pisum sativum | Common cis-acting element in promoter and enhancer regions |
| 1112 | A | T * | CAAT-box | Nicotiana giutinosa | - | |
| 1688 | G | A * | CAAT-box | Nicotiana giutinosa | - | |
| Cla97C02G044090 R | 793 | C * | T | TATA-box | Arabidopsis thaliana | Core promoter element around −30 of transcription start |
| 1724 | G | T * | GARE-motif | Brassica oleracea | Gibberellin-responsive element | |
| Cla97C02G044100 R | 141 | G * | A | CAAT-box | Pisum sativum | Common cis-acting element in promoter and enhancer regions |
| 393 | C * | T | CAAT-box | Nicotiana giutinosa | - | |
| 391 | C | T * | Box 4 | Petroselinum crispum | Part of a conserved DNA module involved in light responsiveness | |
| 407 | T * | A | MRE | Petroselinum crispum | MYB binding site involved in light responsiveness | |
| 410 | T | A * | CAAT-box | Pisum sativum | Common cis-acting element in promoter and enhancer regions | |
| Cla97C02G044110 R | 397 | C * | T | CAAT-box | Arabidopsis thaliana | Common cis-acting element in promoter and enhancer regions |
| 398 | C * | T | CAAT-box | Nicotiana giutinosa | - | |
| Cla97C02G044120 | 25 | C * | T | CAAT-box | Petunia hybrida | Common cis-acting element in promoter and enhancer regions |
| 1083 | C | A * | CTAG-motif | Avena sativa | - | |
| Cla97C02G044130 | 151 | C * | G | MYB-binding site | Nicotiana tabacum | - |
| 151 | A * | T | MYB-binding site | Nicotiana tabacum | - | |
| Cla97C02G044140 | 731 | T * | G | CAAT-box | Nicotiana glutinosa | - |
| 1097 | C * | T | Unnamed__4 | Petroselinum hortense | - | |
| 1285 | T | C * | CAAT-box | Pisum sativum | Common cis-acting element in promoter and enhancer regions |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Pei, S.; Song, Y.; Yang, T.; Gao, Y.; Chai, H.; Luan, F.; Zhu, Z.; Wang, X. Molecular Markers and Marker-Assisted Selection Provide Genetic Insights for Identifying Key Quantitative Trait Locus for Watermelon Rind Thickness. Int. J. Mol. Sci. 2024, 25, 10341. https://doi.org/10.3390/ijms251910341
Zhao Z, Pei S, Song Y, Yang T, Gao Y, Chai H, Luan F, Zhu Z, Wang X. Molecular Markers and Marker-Assisted Selection Provide Genetic Insights for Identifying Key Quantitative Trait Locus for Watermelon Rind Thickness. International Journal of Molecular Sciences. 2024; 25(19):10341. https://doi.org/10.3390/ijms251910341
Chicago/Turabian StyleZhao, Zhengxiang, Shuang Pei, Yuying Song, Tiantian Yang, Yuan Gao, Hao Chai, Feishi Luan, Zicheng Zhu, and Xuezheng Wang. 2024. "Molecular Markers and Marker-Assisted Selection Provide Genetic Insights for Identifying Key Quantitative Trait Locus for Watermelon Rind Thickness" International Journal of Molecular Sciences 25, no. 19: 10341. https://doi.org/10.3390/ijms251910341