
Mitochondrial Genome Characteristics and Phylogenetic Analysis of Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae)

Abstract
:1. Introduction
2. Results
2.1. Mitochondrial Genome Composition
2.2. Analysis of Protein-Coding Genes
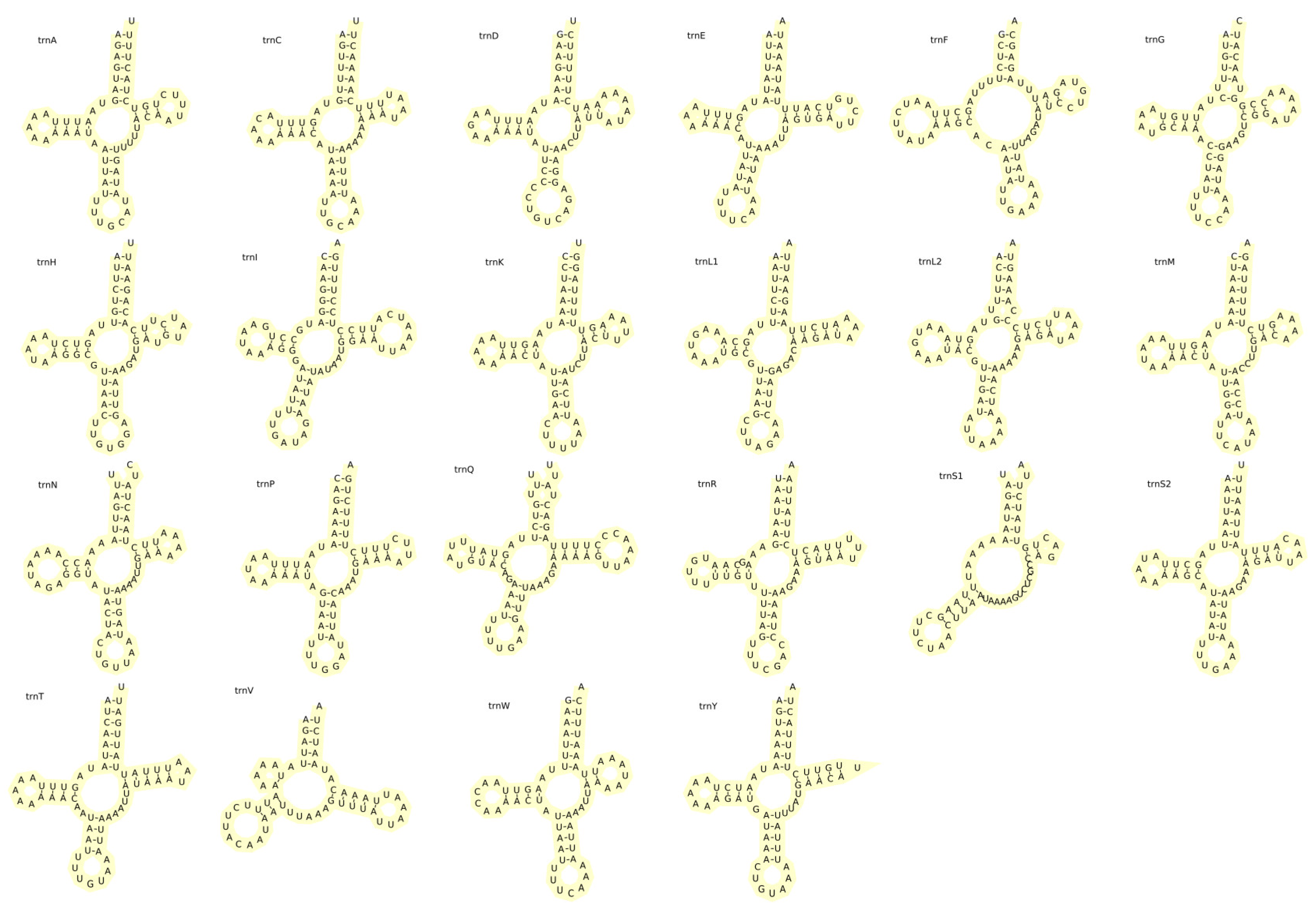
2.3. tRNA Gene and rRNA Gene
2.4. Gene Arrangement
2.5. Analysis of Phylogenetic
3. Discussion
4. Materials and Methods
4.1. Insect Sample Collection and DNA Extraction
4.2. High-Throughput Sequencing and Mitochondrial Genome Assembly
4.3. Mitochondrial Genome Annotation and Analysis
4.4. Phylogenetic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [PubMed]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.F.; Barker, S.C. The highly rearranged mitochondrial genome of the plague thrips, Thrips imaginis (Insecta: Thysanoptera): Convergence of two novel gene boundaries and an extraordinary arrangement of rRNA genes. Mol. Biol. Evol. 2003, 20, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Karimi, J.; Hassani-Kakhki, M.; Awal, M.M. Identifying thrips (Insecta: Thysanoptera) using DNA Barcodes. J. Cell Mol. Res. 2010, 2, 35–41. [Google Scholar]
- Rebijith, K.B.; Asokan, R.; Hande, H.R.; Kumar, N.K.K. The first report of miRNAs from a Thysanopteran insect, Thrips palmi Karny using high-throughput sequencing. PLoS ONE 2016, 11, e0163635. [Google Scholar] [CrossRef]
- Tyagi, K.; Chakraborty, R.; Cameron, S.L.; Sweet, A.D.; Chandra, K.; Kumar, V. Rearrangement and evolution of mitochondrial genomes in Thysanoptera (Insecta). Sci. Rep. 2020, 10, 695. [Google Scholar] [CrossRef]
- ThripsWiki. ThripsWiki–Providing Information on the World’s Thrips. Available online: https://bugguide.net/node/view/1694997 (accessed on 9 March 2023).
- An, Y.Y.; Li, Y.Q.; Dang, L.H. Analysis on the fauna composition of Thysanoptera in the Shaanxi Qinling-Daba Mountains area. J. Environ. Entomol. 2024. Available online: https://link.cnki.net/urlid/44.1640.Q.20240724.0854.002 (accessed on 24 July 2024).
- Han, Y.; Tang, L.D.; Wu, J.H. Researches advances on integrated pest management of thrips (Thysanoptera). Chin. Agric. Sci. Bull. 2015, 31, 163–174. [Google Scholar]
- Xie, D.L.; Wang, X.; Zhao, L.; Dang, L.H. Characterization and comparative analysis of the mitochondrial genomes on Thysanoptera (Insecta: Arthropoda). Chin. Bull. Life Sci. 2020, 32, 621–629. [Google Scholar]
- Chao, Y.F. Economic Insect Fauna of China Fasc. 55 Thysanoptera; Science Press: Beijing, China, 1997; pp. 1–514. [Google Scholar]
- Yin, J.; Luo, Z.M.; Huang, Y.K.; Li, W.F.; Zhang, R.Y.; Wang, X.Y.; Shan, H.L.; Yang, K. Population dynamics of sugarcane thrips (Fulmekiola serrata) and the degree of damage inflicted by these on different sugarcane varieties. Chin. J. Appl. Entomol. 2015, 52, 1438–1443. [Google Scholar]
- Way, M.J.; Stiller, M.; Leslie, G.W.; Conlong, D.E.; Keeping, M.G.; Rutherford, R.S. Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae), a new pest in Southern African sugarcane. Afr. Entomol. 2006, 14, 401–403. [Google Scholar]
- Mirab-balou, M.; Tong, X.L.; Feng, J.N.; Chen, X.X. Thrips (Insecta: Thysanoptera) of China. Check List 2011, 7, 720–744. [Google Scholar] [CrossRef]
- Soto-Adames, F.N. Fulmekiola serrata (Kobus), Sugarcane Thrips; FDACS-P-01605, Florida Department of Agriculture and Consumer Services, Division of Plant Industry: Gainesville, FL, USA, 2017. [Google Scholar]
- Li, Y.R. Modern Sugarcane Science; China Agriculture Press: Beijing, China, 2010; p. 355. [Google Scholar]
- Huang, Y.K.; Li, W.F. Colored Atlas of Diseases, Insect Pests and Weeds of Modern Sugarcane; China Agriculture Press: Beijing, China, 2011; pp. 11–12. [Google Scholar]
- Way, M.J.; Rutherford, R.S.; Swepersad, C.; Leslie, G.W.; Keeping, M.G. Impact of sugarcane thrips, Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae), on sugarcane yield in field trials. Pro. S. Afr. Sug. Technol. Ass. 2010, 83, 244–256. [Google Scholar]
- Luo, Z.M.; Yin, J.; Li, Y.H.; Zhang, R.Y.; Li, J. Study on damage loss of sugarcane thrips. Sugarcane Canesugar 2022, 51, 20–23. [Google Scholar]
- Yin, J.; Luo, Z.M.; Huang, Y.K.; Li, W.F.; Zhang, R.Y.; Wang, X.Y.; Shan, H.L.; Cang, X.Y. Spatial distribution pattern and theoretical sampling of sugarcane thrips (Fulmekiola serrata Kobus) on different sugarcane varieties. J. Environ. Entomol. 2018, 40, 702–707. [Google Scholar]
- Luo, Z.M.; Wang, X.Y.; Huang, Y.K.; Zhang, R.Y.; Li, W.F.; Shan, H.L.; Cang, X.Y.; Li, J.; Yin, J. Field resistance of different sugarcane varieties to sugarcane thrips (Fulmekiola serratus) in China. Sugar Tech. 2019, 21, 527–531. [Google Scholar] [CrossRef]
- Timmanna, H.N.; Meshram, N.M.; Raju, S.; Shashank, P.R.; Jamuna, B. Diagnosis and morphometrics of sugarcane thrips, Fulmekiola serrata (Kobus, 1892) (Thysanoptera: Thripidae). Sugar Tech. 2021, 23, 1377–1382. [Google Scholar] [CrossRef]
- Luo, Z.M.; Yin, J.; Li, J.; Deng, J. The field control effects of five insecticides against sugarcane thrips. Sugarcane Canesugar 2021, 50, 57–60. [Google Scholar]
- Luo, Z.M.; Qin, W.; Yin, J.; Li, Y.H.; Zhang, R.Y.; Li, J. Tolerance of sugarcane germplasm to sugarcane thrips. Chin. Agric. Sci. Bull. 2022, 38, 107–112. [Google Scholar]
- Clary, D.O.; Wolstenholme, D.R. The mitochondrial DNA molecular of Drosophila yakuba: Nucleotide sequence, gene organization, and genetic code. J. Mol. Evol. 1985, 22, 252–271. [Google Scholar]
- Chen, Z.T.; Du, Y.Z. Rearrangement of mitochondrial genome in insects. J. Environ. Entomol. 2016, 38, 843–851. [Google Scholar]
- Hassanin, A.; Léger, N.; Deutsch, J. Evidence for multiple reversals of asymmetric mutational constraints during the evolution of the mitochondrial genome of metazoa, and consequences for phylogenetic inferences. Syst. Biol. 2005, 54, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.M.; Li, P.; Cai, D.C.; Li, J.H. Characterization of the complete mitochondrial genome and phylogenetic analysis of bean thrips Megalurothrips usitatus (Bagnall, 1913) (Thysanoptera: Thripidae). Mitochondrial DNA B 2023, 8, 1032–1035. [Google Scholar]
- Wang, Y.Y.; Wu, C.Y.; Xie, Y.L.; Liu, H.; Zhang, H.R. Complete mitochondrial genome sequence for the Thrips hawaiiensis (Thysanoptera: Thripidae). Mitochondrial DNA B 2021, 6, 2092–2094. [Google Scholar] [CrossRef]
- Pakrashi, A.; Tyagi, K.; Kumar, V. The complete mitochondrial genome of Taeniothrips tigris Bhatti, 1995 (Thysanoptera: Thripidae). Mitochondrial DNA B 2021, 6, 2256–2257. [Google Scholar] [CrossRef]
- Pakrashi, A.; Tyagi, K.; Kumar, V. The complete mitochondrial genome of Aeolothrips indicus Bhatti, 1964 (Thysanoptera: Thripidae). Mitochondrial DNA B 2021, 6, 2808–2809. [Google Scholar] [CrossRef]
- Hu, Q.L.; Ye, Z.X.; Zhang, C.X. High-throughput sequencing yields a complete mitochondrial genome of the rice thrips, Stenchaetothrips biformis (Thysanoptera: Thripidae). Mitochondrial DNA B 2023, 8, 204–206. [Google Scholar] [CrossRef]
- Pakrashi, A.; Patidar, A.; Singha, D.; Kumar, V.; Tyagi, K. Comparative analysis of the two suborders of Thysanoptera and characterization of the complete mitochondrial genome of Thrips parvispinus. Arch. Insect Biochem. Physiol. 2023, 114, 1–15. [Google Scholar] [CrossRef]
- Dang, L.H.; Li, C.W.; Wang, D.X.; Gao, Y.X.; Zhao, L.P.; Zhang, B. Characterizing the complete mitochondrial genome of Psephenothrips eriobotryae Dang & Qiao (Thysanoptera: Phlaeothripidae) with massive gene arrangement in Phlaeothripidae. Zool. Syst. 2024, 49, 116–128. [Google Scholar]
- Li, C.W.; Gao, Y.X.; Wang, D.X.; Dang, L.H. The extraordinary rearrangement of mitochondrial genome of the wheat pest, Aptinothrips stylifer and the mitochondrial phylogeny of Thripidae (Thysanoptera). Arch. Insect Biochem. Physiol. 2024, 115, e22086. [Google Scholar] [CrossRef]
- Liu, H.R.; Li, H.; Song, F.; Gu, W.Y.; Feng, J.N.; Cai, W.Z.; Shao, R.F. Novel insights into mitochondrial gene rearrangement in thrips (Insecta: Thysanoptera) from the grass thrips, Anaphothrips obscurus. Sci. Rep. 2017, 7, 4284. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Li, H.; Shao, R.F.; Shi, A.M.; Bai, X.S.; Zheng, X.R.; Heiss, E.; Cai, W.Z. Rearrangement of mitochondrial tRNA genes in flat bugs (Hemiptera: Aradidae). Sci. Rep. 2016, 6, 25725. [Google Scholar] [CrossRef] [PubMed]
- Dowton, M.; Campbell, N.J.H. Intramitochondrial recombination—Is it why some mitochondrial genes sleep around? Trends Ecol. Evol. 2001, 16, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Buckman, R.S.; Mound, L.A.; Whiting, M.F. Phylogeny of thrips (Insecta: Thysanoptera) based on five molecular loci. Syst. Entomol. 2013, 38, 123–133. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Z.; Mao, C.; Pan, Z.; Yao, Y.; He, J.; Lin, Y.; Dong, Z.; Liu, G.; Zhao, R.; et al. Complete mitogenome and phylogenetic significance of Metoecus javanus (Pic, 1913) (Coleoptera: Ripiphoridae) from Southwest China, with notes on morphological traits of adult and immature stages. Zootaxa 2022, 5205, 231–248. [Google Scholar] [CrossRef]
- Pearson, W.R. Rapid and sensitive sequence comparison with FASTP and FASTA. Method. Enzymol. 1990, 183, 63–98. [Google Scholar]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Strand | Location | Size (bp) | Anticodon | Start Codon | Stop Codon | Intergenic Nucleotides |
|---|---|---|---|---|---|---|---|
| cox1 | J | 1–1500 | 1500 | - | ATC | TAA | 192 |
| nad3 | J | 1497–1848 | 352 | - | ATA | T-- | −4 |
| trnL2 | J | 1849–1913 | 65 | TAA | 0 | ||
| cox2 | J | 1914–2570 | 657 | - | ATA | TAA | 0 |
| trnG | J | 2572–2634 | 63 | TCC | 1 | ||
| trnK | J | 2634–2695 | 62 | TTT | −1 | ||
| cox3 | J | 2708–3490 | 783 | - | ATA | TAA | 12 |
| trnN | J | 3497–3560 | 64 | GTT | 6 | ||
| trnT | J | 3558–3622 | 65 | TGT | −3 | ||
| trnS1 | J | 3627–3682 | 56 | TCT | 4 | ||
| trnD | J | 3698–3761 | 64 | GTC | 15 | ||
| trnR | J | 3765–3830 | 66 | TCG | 3 | ||
| trnL1 | J | 3829–3893 | 65 | TAG | −2 | ||
| trnE | J | 3894–3958 | 65 | TTC | 0 | ||
| trnP | N | 4529–4592 | 64 | TGG | 570 | ||
| trnI | J | 4595–4662 | 68 | GAT | 2 | ||
| cob | J | 4664–5779 | 1116 | - | ATA | TAA | 1 |
| trnY | N | 5784–5846 | 63 | GTA | 4 | ||
| nad2 | J | 5874–6846 | 973 | - | ATA | T-- | 27 |
| trnW | J | 6847–6909 | 63 | TCA | 0 | ||
| nad1 | J | 6910–7829 | 920 | - | ATA | TA- | 0 |
| trnM | J | 7830–7892 | 63 | CAT | 0 | ||
| trnA | J | 7895–7957 | 63 | TGC | 2 | ||
| trnF | J | 7958–8024 | 67 | GAA | 0 | ||
| rrnS | J | 8025–8746 | 722 | - | 0 | ||
| atp8 | J | 8747–8915 | 169 | - | ATT | T-- | 0 |
| atp6 | J | 8909–9568 | 660 | - | ATT | TAA | −7 |
| trnQ | J | 9569–9636 | 68 | TTG | 0 | ||
| trnS2 | J | 9667–9730 | 64 | TGA | 30 | ||
| D-loop | J | 9731–10,980 | 1250 | - | 0 | ||
| nad5 | N | 10,981–12,663 | 1683 | - | TTG | TAA | 0 |
| trnH | N | 12,795–12,857 | 63 | GTG | 131 | ||
| nad4 | N | 12,987–14,306 | 1320 | - | ATT | TAA | 129 |
| nad4l | N | 14,300–14,575 | 276 | - | ATG | TAA | −7 |
| trnC | J | 14,603–14,664 | 62 | GCA | 27 | ||
| nad6 | J | 14,684–15,169 | 486 | - | ATA | TAA | 19 |
| trnV | J | 15,213–15,268 | 56 | TAC | 43 | ||
| rrnL | J | 15,269–16,404 | 1136 | - | 0 |
| Fulmekiola serrata | Size (bp) | A% | T% | G% | C% | A+T% | G+C% | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|---|---|
| Whole mitogenome | 16,596 | 42.28 | 32.72 | 11 | 14 | 75 | 25 | 0.128 | −0.12 |
| PCGs | 10,895 | 31.75 | 42.47 | 11.85 | 13.93 | 74.22 | 25.78 | −0.144 | −0.081 |
| tRNAs | 1399 | 42.6 | 36.81 | 10.65 | 9.94 | 79.41 | 20.59 | 0.073 | 0.035 |
| rRNAs | 1858 | 46.72 | 31.81 | 11.09 | 10.39 | 78.53 | 21.47 | 0.19 | 0.033 |
| Control region | 1250 | 41.68 | 31.84 | 10.96 | 15.52 | 73.52 | 26.48 | 0.134 | −0.172 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.; Luo, Z.-M.; Li, Y.-H.; Wang, C.-M.; Li, J.; Zhang, R.-Y.; Shan, H.-L.; Wang, X.-Y.; Chen, Y.-Q. Mitochondrial Genome Characteristics and Phylogenetic Analysis of Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae). Int. J. Mol. Sci. 2024, 25, 10431. https://doi.org/10.3390/ijms251910431
Yin J, Luo Z-M, Li Y-H, Wang C-M, Li J, Zhang R-Y, Shan H-L, Wang X-Y, Chen Y-Q. Mitochondrial Genome Characteristics and Phylogenetic Analysis of Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae). International Journal of Molecular Sciences. 2024; 25(19):10431. https://doi.org/10.3390/ijms251910431
Chicago/Turabian StyleYin, Jiong, Zhi-Ming Luo, Yin-Hu Li, Chang-Mi Wang, Jie Li, Rong-Yue Zhang, Hong-Li Shan, Xiao-Yan Wang, and You-Qing Chen. 2024. "Mitochondrial Genome Characteristics and Phylogenetic Analysis of Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae)" International Journal of Molecular Sciences 25, no. 19: 10431. https://doi.org/10.3390/ijms251910431
APA StyleYin, J., Luo, Z. -M., Li, Y. -H., Wang, C. -M., Li, J., Zhang, R. -Y., Shan, H. -L., Wang, X. -Y., & Chen, Y. -Q. (2024). Mitochondrial Genome Characteristics and Phylogenetic Analysis of Fulmekiola serrata (Kobus) (Thysanoptera: Thripidae). International Journal of Molecular Sciences, 25(19), 10431. https://doi.org/10.3390/ijms251910431