
The Activation of Muscarinic Acetylcholine Receptors Protects against Neuroinflammation in a Mouse Model through Attenuating Microglial Inflammation

 ,
, 
Abstract
1. Introduction
2. Results
2.1. The Activation of Muscarinic Receptors Attenuated Neuroinflammation in an LPS-Induced Mouse Model
2.2. The Activation of Muscarinic Receptors Regulated the Expression Level of Cytokines in the Brains and Spinal Cords of Mouse Neuroinflammation Models
2.3. The Levels of Inflammation in Primary Microglia Were Rescued by the Activation of Muscarinic Receptors
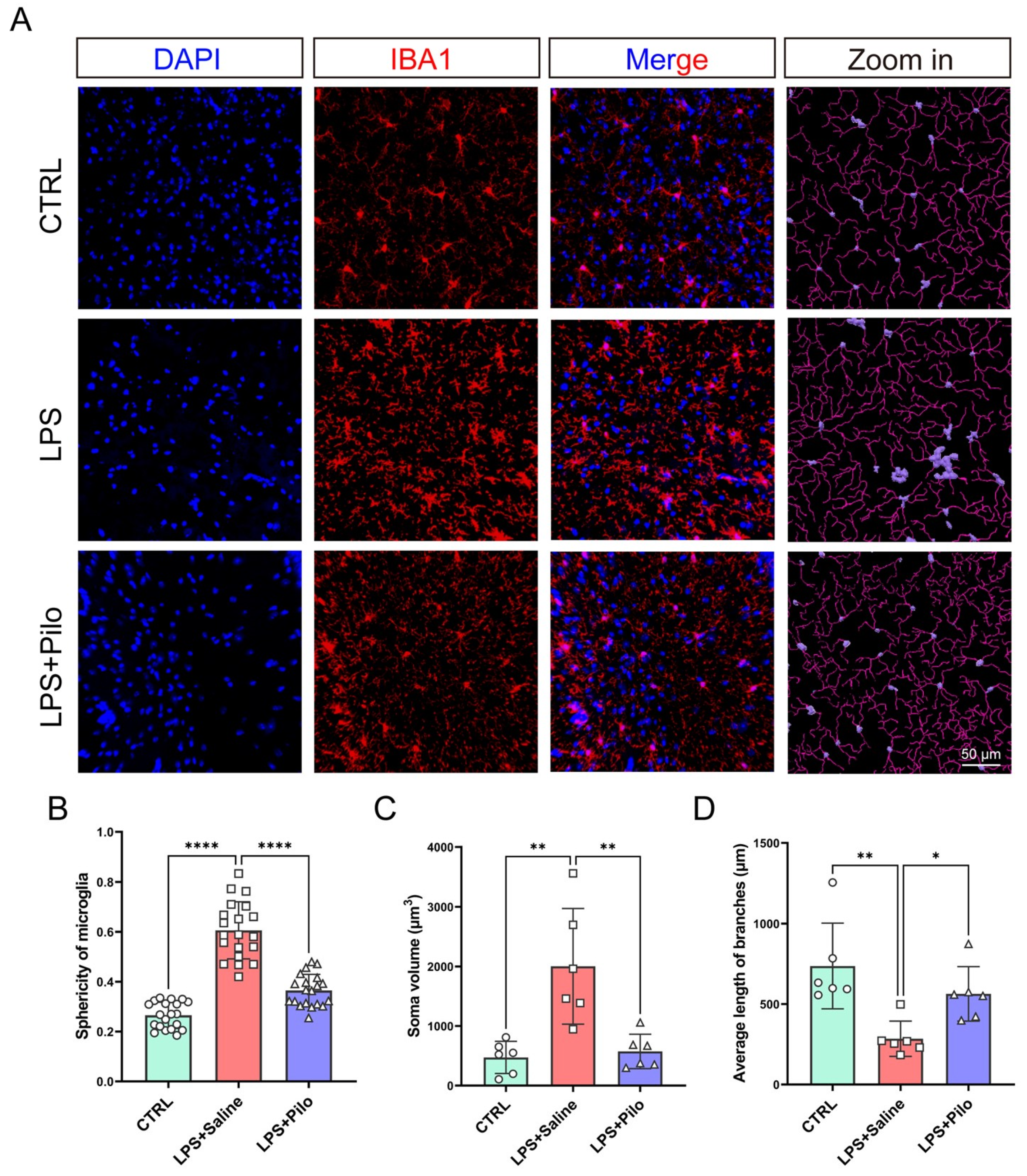
2.4. Pilocarpine-Attenuated Microgliosis in Mouse Neuroinflammation Model
2.5. Pilocarpine Improved Morphological Changes in Activated Microglia
2.6. Activation of Muscarinic Receptors Protected against Neuronal Damage through Direct and Indirect Effects
3. Discussion
4. Materials and Methods
4.1. Animals and LPS-Induced Neuroinflammation Model
4.2. Cell Culture and Treatment
4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. RNA Extraction and Quantitative Reverse-Transcription Polymerase Chain Reaction (RT-qPCR) Analysis
4.5. Tissue Preparation for Cryotomy
4.6. Morphological Analysis
4.7. Cell Viability Assay
4.8. Western Blot
4.9. Immunofluorescence Staining
4.10. TUNEL Staining
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol. Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Ordoñez, W.O.; Cajas-Salazar, N.; Velasco-Reyes, M.A. Genetic and epigenetic targets of natural dietary compounds as anti-Alzheimer’s agents. Neural Regen. Res. 2024, 19, 846–854. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.; Shireman, J.; Sierra Potchanant, E.A.; Lara-Velazquez, M.; Dey, M. Neuroinflammation in Autoimmune Disease and Primary Brain Tumors: The Quest for Striking the Right Balance. Front. Cell. Neurosci. 2021, 15, 716947. [Google Scholar] [CrossRef] [PubMed]
- Adler, O.; Zait, Y.; Cohen, N.; Blazquez, R.; Doron, H.; Monteran, L.; Scharff, Y.; Shami, T.; Mundhe, D.; Glehr, G.; et al. Reciprocal interactions between innate immune cells and astrocytes facilitate neuroinflammation and brain metastasis via lipocalin-2. Nat. Cancer 2023, 4, 401–418. [Google Scholar] [CrossRef] [PubMed]
- Sadlon, A.; Takousis, P.; Ankli, B.; Alexopoulos, P.; Perneczky, R. Association of Chronic Pain with Biomarkers of Neurodegeneration, Microglial Activation, and Inflammation in Cerebrospinal Fluid and Impaired Cognitive Function. Ann. Neurol. 2023, 95, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Chang, Q.; Sun, T.; He, X.; Wen, L.; An, J.; Feng, J.; Zhao, Y. Metabolic reprogramming and polarization of microglia in Parkinson’s disease: Role of inflammasome and iron. Ageing Res. Rev. 2023, 90, 102032. [Google Scholar] [CrossRef]
- Kloske, C.M.; Barnum, C.J.; Batista, A.F.; Bradshaw, E.M.; Brickman, A.M.; Bu, G.; Dennison, J.; Gearon, M.D.; Goate, A.M.; Haass, C.; et al. APOE and immunity: Research highlights. Alzheimers Dement. 2023, 19, 2677–2696. [Google Scholar] [CrossRef] [PubMed]
- Paolicelli, R.C.; Sierra, A.; Stevens, B.; Tremblay, M.-E.; Aguzzi, A.; Ajami, B.; Amit, I.; Audinat, E.; Bechmann, I.; Bennett, M.; et al. Microglia states and nomenclature: A field at its crossroads. Neuron 2022, 110, 3458–3483. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Masuda, T.; Wheeler, M.A.; Quintana, F.J. Microglia and Central Nervous System-Associated Macrophages-From Origin to Disease Modulation. Annu. Rev. Immunol. 2021, 39, 251–277. [Google Scholar] [CrossRef]
- Wendimu, M.Y.; Hooks, S.B. Microglia Phenotypes in Aging and Neurodegenerative Diseases. Cells 2022, 11, 2091. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, S.; Basilico, B.; Marrone, M.C.; Ragozzino, D. Microglia-neuron crosstalk: Signaling mechanism and control of synaptic transmission. Semin. Cell Dev. Biol. 2019, 94, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef]
- Cserép, C.; Pósfai, B.; Lénárt, N.; Fekete, R.; László, Z.I.; Lele, Z.; Orsolits, B.; Molnár, G.; Heindl, S.; Schwarcz, A.D.; et al. Microglia monitor and protect neuronal function through specialized somatic purinergic junctions. Science 2020, 367, 528–537. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, D.; Mao, Q.; Xia, H. Role of neuroinflammation in neurodegeneration development. Signal Transduct. Target. Ther. 2023, 8, 267. [Google Scholar] [CrossRef] [PubMed]
- Ciryam, P.; Gerzanich, V.; Simard, J.M. Interleukin-6 in Traumatic Brain Injury: A Janus-Faced Player in Damage and Repair. J. Neurotrauma 2023, 40, 2249–2269. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Yu, P.; Lin, W.-J.; Chen, S.; Hu, X.; Chen, S.; Cheng, J.; Liu, Q.; Yang, Y.; Li, S.; et al. Microglia drive transient insult-induced brain injury by chemotactic recruitment of CD8+ T lymphocytes. Neuron 2023, 111, 696–710. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.C.; Lalor, S.J.; Lynch, M.A.; Mills, K.H.G. Infiltration of Th1 and Th17 cells and activation of microglia in the CNS during the course of experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2010, 24, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Moser, T.; Akgün, K.; Proschmann, U.; Sellner, J.; Ziemssen, T. The role of TH17 cells in multiple sclerosis: Therapeutic implications. Autoimmun. Rev. 2020, 19, 102647. [Google Scholar] [CrossRef]
- Amo-Aparicio, J.; Garcia-Garcia, J.; Francos-Quijorna, I.; Urpi, A.; Esteve-Codina, A.; Gut, M.; Quintana, A.; Lopez-Vales, R. Interleukin-4 and interleukin-13 induce different metabolic profiles in microglia and macrophages that relate with divergent outcomes after spinal cord injury. Theranostics 2021, 11, 9805–9820. [Google Scholar] [CrossRef]
- Shemer, A.; Scheyltjens, I.; Frumer, G.R.; Kim, J.-S.; Grozovski, J.; Ayanaw, S.; Dassa, B.; Van Hove, H.; Chappell-Maor, L.; Boura-Halfon, S.; et al. Interleukin-10 Prevents Pathological Microglia Hyperactivation following Peripheral Endotoxin Challenge. Immunity 2020, 53, 1033–1049. [Google Scholar] [CrossRef]
- Zhao, L.-X.; Chen, M.-W.; Qian, Y.; Yang, Q.-H.; Ge, Y.-H.; Chen, H.-Z.; Qiu, Y. M1 Muscarinic Receptor Activation Rescues β-Amyloid-Induced Cognitive Impairment through AMPA Receptor GluA1 Subunit. Neuroscience 2019, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Lenina, O.A.; Petrov, K.A. Balanced modulation of neuromuscular synaptic transmission via M1 and M2 muscarinic receptors during inhibition of cholinesterases. Sci. Rep. 2022, 12, 1688. [Google Scholar] [CrossRef] [PubMed]
- Puhl, C.J.; Wefelmeyer, W.; Burrone, J. Cholinergic Stimulation Modulates the Functional Composition of CA3 Cell Types in the Hippocampus. J. Neurosci. 2023, 43, 4972–4983. [Google Scholar] [CrossRef]
- Dwomoh, L.; Rossi, M.; Scarpa, M.; Khajehali, E.; Molloy, C.; Herzyk, P.; Mistry, S.N.; Bottrill, A.R.; Sexton, P.M.; Christopoulos, A.; et al. M1 muscarinic receptor activation reduces the molecular pathology and slows the progression of prion-mediated neurodegenerative disease. Sci. Signal. 2022, 15, eabm3720. [Google Scholar] [CrossRef] [PubMed]
- Kruse, A.C.; Kobilka, B.K.; Gautam, D.; Sexton, P.M.; Christopoulos, A.; Wess, J. Muscarinic acetylcholine receptors: Novel opportunities for drug development. Nat. Rev. Drug Discov. 2014, 13, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tan, P.; Li, H.; Cui, Y.; Qiu, Y.; Wang, H.; Zhang, X.; Li, J.; Zhu, L.; Zhou, W.; et al. Direct SUMOylation of M1 muscarinic acetylcholine receptor increases its ligand-binding affinity and signal transduction. FASEB J. 2019, 33, 3237–3251. [Google Scholar] [CrossRef]
- Pannell, M.; Meier, M.A.; Szulzewsky, F.; Matyash, V.; Endres, M.; Kronenberg, G.; Prinz, V.; Waiczies, S.; Wolf, S.A.; Kettenmann, H. The subpopulation of microglia expressing functional muscarinic acetylcholine receptors expands in stroke and Alzheimer’s disease. Brain Struct. Funct. 2016, 221, 1157–1172. [Google Scholar] [CrossRef]
- Costa, A.; Haage, V.; Yang, S.; Wegner, S.; Ersoy, B.; Ugursu, B.; Rex, A.; Kronenberg, G.; Gertz, K.; Endres, M.; et al. Deletion of muscarinic acetylcholine receptor 3 in microglia impacts brain ischemic injury. Brain Behav. Immun. 2021, 91, 89–104. [Google Scholar] [CrossRef]
- Cox, M.A.; Duncan, G.S.; Lin, G.H.Y.; Steinberg, B.E.; Yu, L.X.; Brenner, D.; Buckler, L.N.; Elia, A.J.; Wakeham, A.C.; Nieman, B.; et al. Choline acetyltransferase-expressing T cells are required to control chronic viral infection. Science 2019, 363, 639–644. [Google Scholar] [CrossRef]
- Cox, M.A.; Bassi, C.; Saunders, M.E.; Nechanitzky, R.; Morgado-Palacin, I.; Zheng, C.; Mak, T.W. Beyond neurotransmission: Acetylcholine in immunity and inflammation. J. Intern. Med. 2020, 287, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yao, Y.; Wang, S.; Hou, Y.; Zhao, L.; Wang, H.; Chen, H.; Xu, J. Structural Insights into M1 Muscarinic Acetylcholine Receptor Signaling Bias between Gαq and β-Arrestin through BRET Assays and Molecular Docking. Int. J. Mol. Sci. 2023, 24, 7356. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Chen, S.-H.; Zhao, Z.; Tu, D.; Song, S.; Wang, Y.; Wang, Q.; Feng, J.; Hong, J.-S. Complement C3 Enhances LPS-Elicited Neuroinflammation and Neurodegeneration Via the Mac1/NOX2 Pathway. Mol. Neurobiol. 2023, 60, 5167–5183. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Xue, Z.; Zheng, Y.; Li, S.; Zhou, L.; Cao, L.; Zou, Y. Selenium supplementation enhanced the expression of selenoproteins in hippocampus and played a neuroprotective role in LPS-induced neuroinflammation. Int. J. Biol. Macromol. 2023, 234, 123740. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-J.; Ding, Y.; Yin, Y.-Q.; Xiao, H.; Hu, G.; Zhou, J.-W. Cspg4high microglia contribute to microgliosis during neurodegeneration. Proc. Natl. Acad. Sci. USA 2023, 120, e2210643120. [Google Scholar] [CrossRef] [PubMed]
- Candelario-Jalil, E.; Dijkhuizen, R.M.; Magnus, T. Neuroinflammation, Stroke, Blood-Brain Barrier Dysfunction, and Imaging Modalities. Stroke 2022, 53, 1473–1486. [Google Scholar] [CrossRef] [PubMed]
- Qiang, Y.; Song, M.; Wang, S.; Liu, Z.; Shan, S.; Sun, Y.; Ni, W.; Chao, S.; Liu, Z.; Zhao, X.; et al. High-fat diet exacerbated motor dysfunction via necroptosis and neuroinflammation in acrylamide-induced neurotoxicity in mice. Ecotoxicol. Environ. Saf. 2024, 269, 115777. [Google Scholar] [CrossRef]
- Marchi, N.; Oby, E.; Batra, A.; Uva, L.; De Curtis, M.; Hernandez, N.; Van Boxel-Dezaire, A.; Najm, I.; Janigro, D. In vivo and in vitro effects of pilocarpine: Relevance to ictogenesis. Epilepsia 2007, 48, 1934–1946. [Google Scholar] [CrossRef] [PubMed]
- Block, M.L.; Zecca, L.; Hong, J.-S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Tan, P.-P.; Yuan, H.-H.; Zhu, X.; Cui, Y.-Y.; Li, H.; Feng, X.-M.; Qiu, Y.; Chen, H.-Z.; Zhou, W. Activation of muscarinic receptors protects against retinal neurons damage and optic nerve degeneration in vitro and in vivo models. CNS Neurosci. Ther. 2014, 20, 227–236. [Google Scholar] [CrossRef]
- Scott-Hewitt, N.; Mahoney, M.; Huang, Y.; Korte, N.; Yvanka de Soysa, T.; Wilton, D.K.; Knorr, E.; Mastro, K.; Chang, A.; Zhang, A.; et al. Microglial-derived C1q integrates into neuronal ribonucleoprotein complexes and impacts protein homeostasis in the aging brain. Cell 2024, 187, 4193–4212. [Google Scholar] [CrossRef]
- Zabot, G.C.; Medeiros, E.B.; Macarini, B.M.N.; Peruchi, B.B.; Keller, G.S.; Lídio, A.V.; Boaventura, A.; de Jesus, L.C.; de Bem Silveira, G.; Silveira, P.C.L.; et al. The involvement of neuroinflammation in an animal model of dementia and depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2024, 133, 110999. [Google Scholar] [CrossRef] [PubMed]
- Ermakov, E.A.; Mednova, I.A.; Boiko, A.S.; Buneva, V.N.; Ivanova, S.A. Chemokine Dysregulation and Neuroinflammation in Schizophrenia: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 2215. [Google Scholar] [CrossRef]
- Gąssowska-Dobrowolska, M.; Chlubek, M.; Kolasa, A.; Tomasiak, P.; Korbecki, J.; Skowrońska, K.; Tarnowski, M.; Masztalewicz, M.; Baranowska-Bosiacka, I. Microglia and Astroglia-The Potential Role in Neuroinflammation Induced by Pre- and Neonatal Exposure to Lead (Pb). Int. J. Mol. Sci. 2023, 24, 9903. [Google Scholar] [CrossRef]
- Mu, X.; Liu, K.; Li, H.; Wang, F.-S.; Xu, R. Granulocyte-macrophage colony-stimulating factor: An immunotarget for sepsis and COVID-19. Cell Mol. Immunol. 2021, 18, 2057–2058. [Google Scholar] [CrossRef]
- Bhattacharya, P.; Thiruppathi, M.; Elshabrawy, H.A.; Alharshawi, K.; Kumar, P.; Prabhakar, B.S. GM-CSF: An immune modulatory cytokine that can suppress autoimmunity. Cytokine 2015, 75, 261–271. [Google Scholar] [CrossRef]
- Kann, O.; Almouhanna, F.; Chausse, B. Interferon γ: A master cytokine in microglia-mediated neural network dysfunction and neurodegeneration. Trends Neurosci. 2022, 45, 913–927. [Google Scholar] [CrossRef] [PubMed]
- Deczkowska, A.; Amit, I.; Schwartz, M. Microglial immune checkpoint mechanisms. Nat. Neurosci. 2018, 21, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Brandl, S.; Reindl, M. Blood-Brain Barrier Breakdown in Neuroinflammation: Current In Vitro Models. Int. J. Mol. Sci. 2023, 24, 12699. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef]
- Li, Q.; Lan, X.; Han, X.; Durham, F.; Wan, J.; Weiland, A.; Koehler, R.C.; Wang, J. Microglia-derived interleukin-10 accelerates post-intracerebral hemorrhage hematoma clearance by regulating CD36. Brain Behav. Immun. 2021, 94, 437–457. [Google Scholar] [CrossRef]
- Gao, Y.; Tu, D.; Yang, R.; Chu, C.-H.; Hong, J.-S.; Gao, H.-M. Through Reducing ROS Production, IL-10 Suppresses Caspase-1-Dependent IL-1β Maturation, thereby Preventing Chronic Neuroinflammation and Neurodegeneration. Int. J. Mol. Sci. 2020, 21, 465. [Google Scholar] [CrossRef] [PubMed]
- Burmeister, A.R.; Marriott, I. The Interleukin-10 Family of Cytokines and Their Role in the CNS. Front. Cell Neurosci. 2018, 12, 458. [Google Scholar] [CrossRef] [PubMed]
- Kneussel, M.; Friese, M.A. SnapShot: Neuronal dysfunction in inflammation. Neuron 2021, 109, 1754. [Google Scholar] [CrossRef] [PubMed]
- Petkova-Tuffy, A.; Gödecke, N.; Viotti, J.; Korte, M.; Dresbach, T. Neuroligin-1 mediates presynaptic maturation through brain-derived neurotrophic factor signaling. BMC Biol. 2021, 19, 215. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J.; Pogue, A.I. Vesicular Transport of Encapsulated microRNA between Glial and Neuronal Cells. Int. J. Mol. Sci. 2020, 21, 5078. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wei, Y.; Zhou, J.; Zou, H.; Ma, L.; Liu, C.; Xiao, Z.; Liu, X.; Tan, X.; Yu, T.; et al. Activation of locus coeruleus-spinal cord noradrenergic neurons alleviates neuropathic pain in mice via reducing neuroinflammation from astrocytes and microglia in spinal dorsal horn. J. Neuroinflamm. 2022, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Catorce, M.N.; Gevorkian, G. LPS-induced Murine Neuroinflammation Model: Main Features and Suitability for Pre-clinical Assessment of Nutraceuticals. Curr. Neuropharmacol. 2016, 14, 155–164. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Liu, D.; Li, X.; He, L.; Pan, J.; Shen, Q.; Peng, Y. CB2R activation ameliorates late adolescent chronic alcohol exposure-induced anxiety-like behaviors during withdrawal by preventing morphological changes and suppressing NLRP3 inflammasome activation in prefrontal cortex microglia in mice. Brain Behav. Immun. 2023, 110, 60–79. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name 1 | Primer Sequence (5′–3′) |
|---|---|
| IL-1β | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
| IL-4 | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
| IL-6 | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
| IL-10 | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
| IL-17 | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
| IL-23 | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
| TNF-α | Forward: GGAACTGGCAGAAGAGGCACTC Reverse: GGAATGAGAAGAG-GCTGAGACATAGG |
| GM-CSF | Forward: GCUUAUCAUCAGAAAGGUA Reverse: UACCUUUCUGAUGAUAAGC |
| IFN-γ | Forward: GCAUCAUCGUUUCCUACAA Reverse: UUGUAGGAAACGAUGAUGC |
| Ccl2 | Forward: GCAUCAUCGUUUCCUACAA Reverse: UUGUAGGAAACGAUGAUGC |
| Ccl7 | Forward: GAGACTTCCATCCAGTTGCCTTCT Reverse: GTGTAATTAAGCCTCCGACTTGTGAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Xie, Y.; Chen, X.; Ouyang, X.; Zhao, L.; Chen, H.; Xu, J. The Activation of Muscarinic Acetylcholine Receptors Protects against Neuroinflammation in a Mouse Model through Attenuating Microglial Inflammation. Int. J. Mol. Sci. 2024, 25, 10432. https://doi.org/10.3390/ijms251910432
Wang K, Xie Y, Chen X, Ouyang X, Zhao L, Chen H, Xu J. The Activation of Muscarinic Acetylcholine Receptors Protects against Neuroinflammation in a Mouse Model through Attenuating Microglial Inflammation. International Journal of Molecular Sciences. 2024; 25(19):10432. https://doi.org/10.3390/ijms251910432
Chicago/Turabian StyleWang, Kaichun, Yuanyuan Xie, Xixiang Chen, Xiaoyan Ouyang, Lanxue Zhao, Hongzhuan Chen, and Jianrong Xu. 2024. "The Activation of Muscarinic Acetylcholine Receptors Protects against Neuroinflammation in a Mouse Model through Attenuating Microglial Inflammation" International Journal of Molecular Sciences 25, no. 19: 10432. https://doi.org/10.3390/ijms251910432
APA StyleWang, K., Xie, Y., Chen, X., Ouyang, X., Zhao, L., Chen, H., & Xu, J. (2024). The Activation of Muscarinic Acetylcholine Receptors Protects against Neuroinflammation in a Mouse Model through Attenuating Microglial Inflammation. International Journal of Molecular Sciences, 25(19), 10432. https://doi.org/10.3390/ijms251910432