
SHH Signaling as a Key Player in Endometrial Cancer: Unveiling the Correlation with Good Prognosis, Low Proliferation, and Anti-Tumor Immune Milieu
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
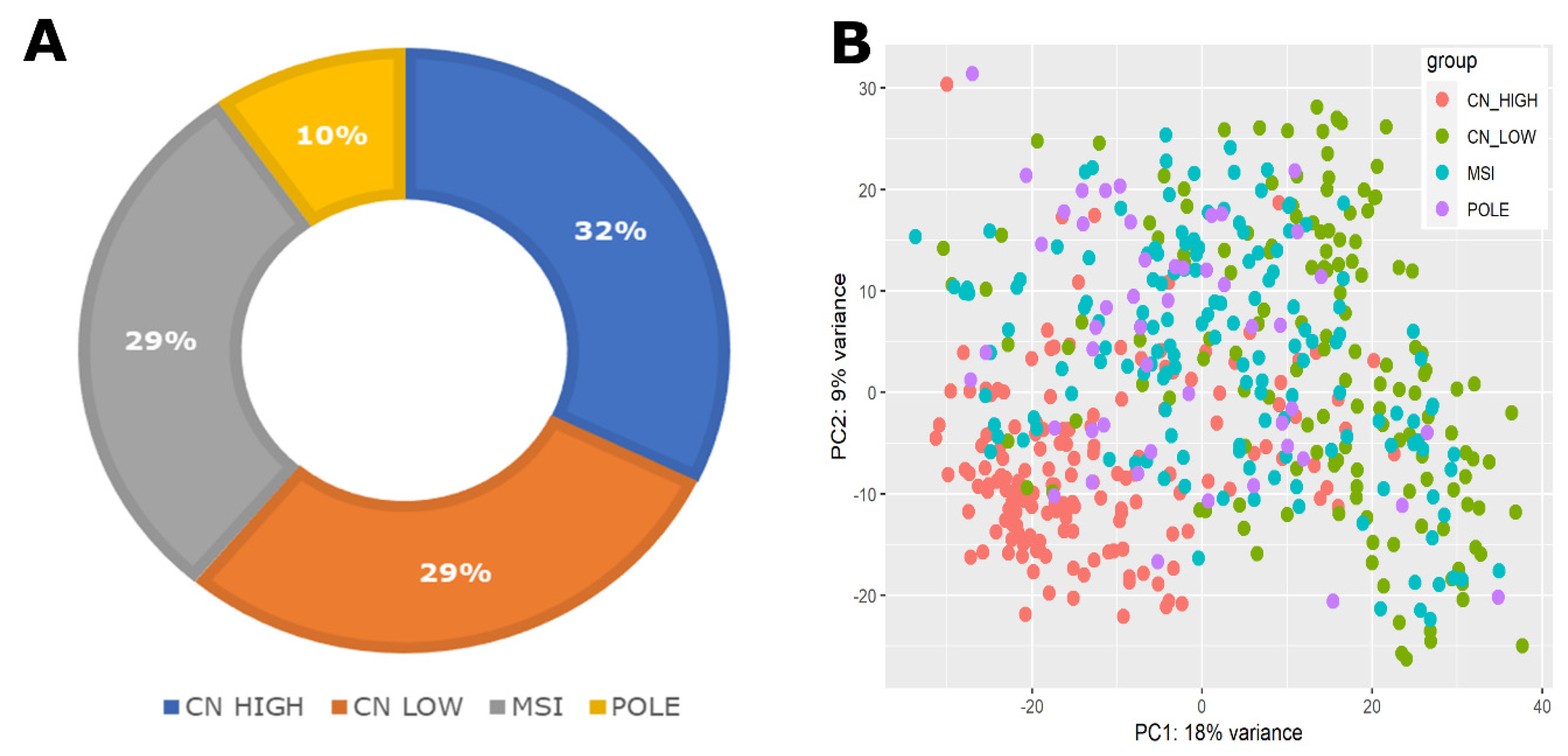
2.1. Comprehensive Dataset Overview and Clinical Characteristics
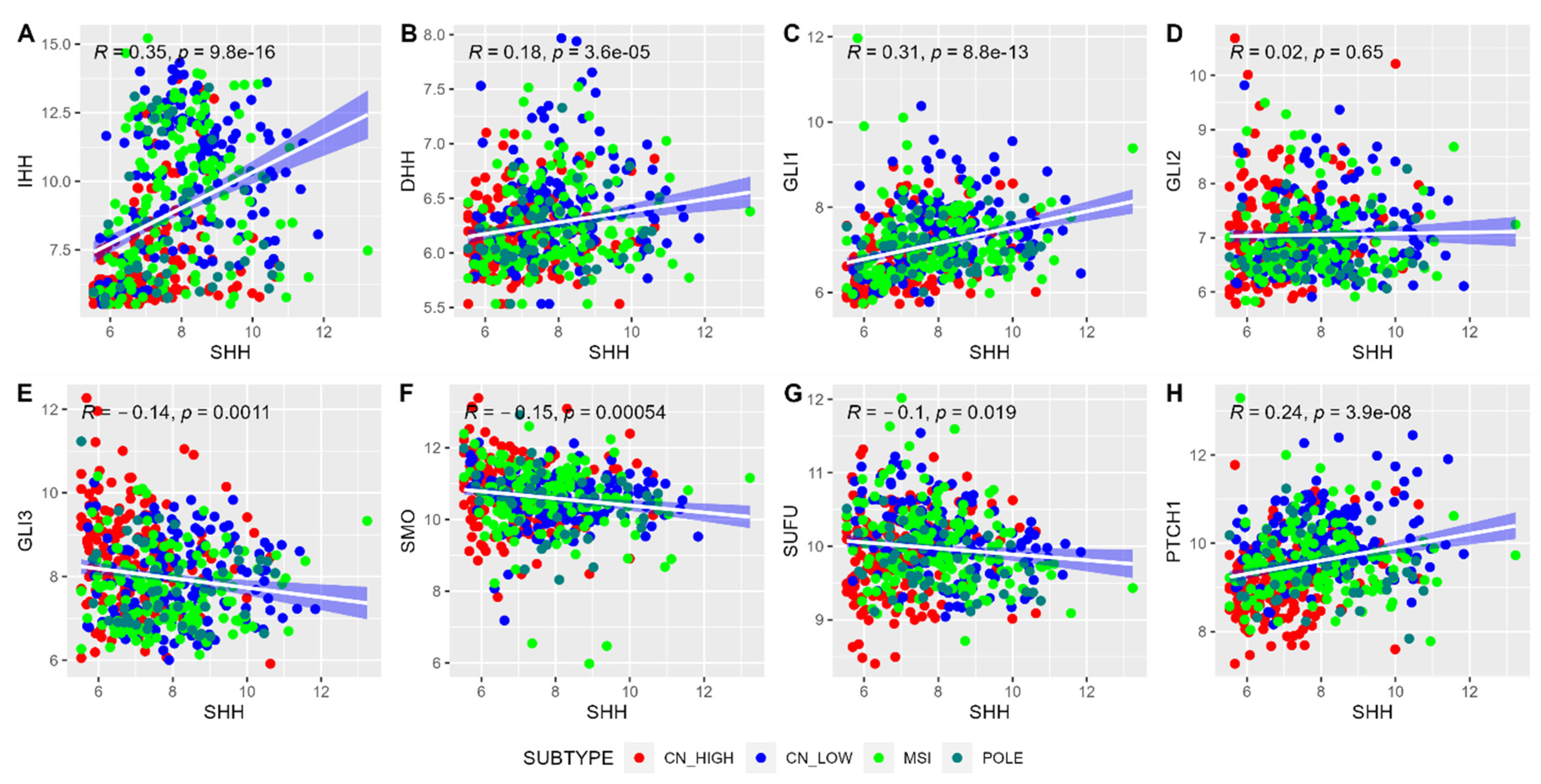
2.2. Correlation Analysis and Composite Gene Signature Score for SHH Pathway Genes
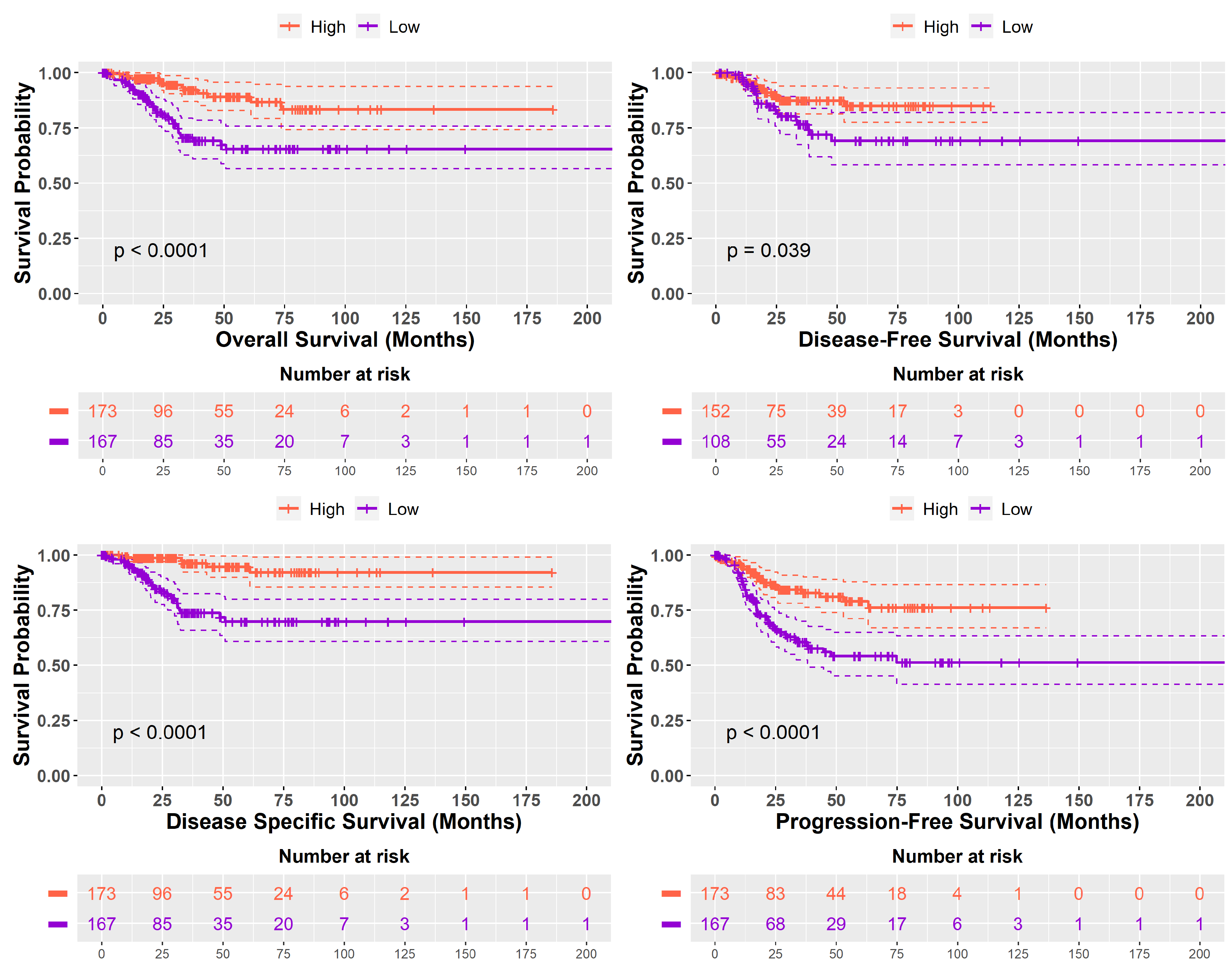
2.3. Prognostic Implications of High SHH and Low SHH Tumors Levels in Cancer
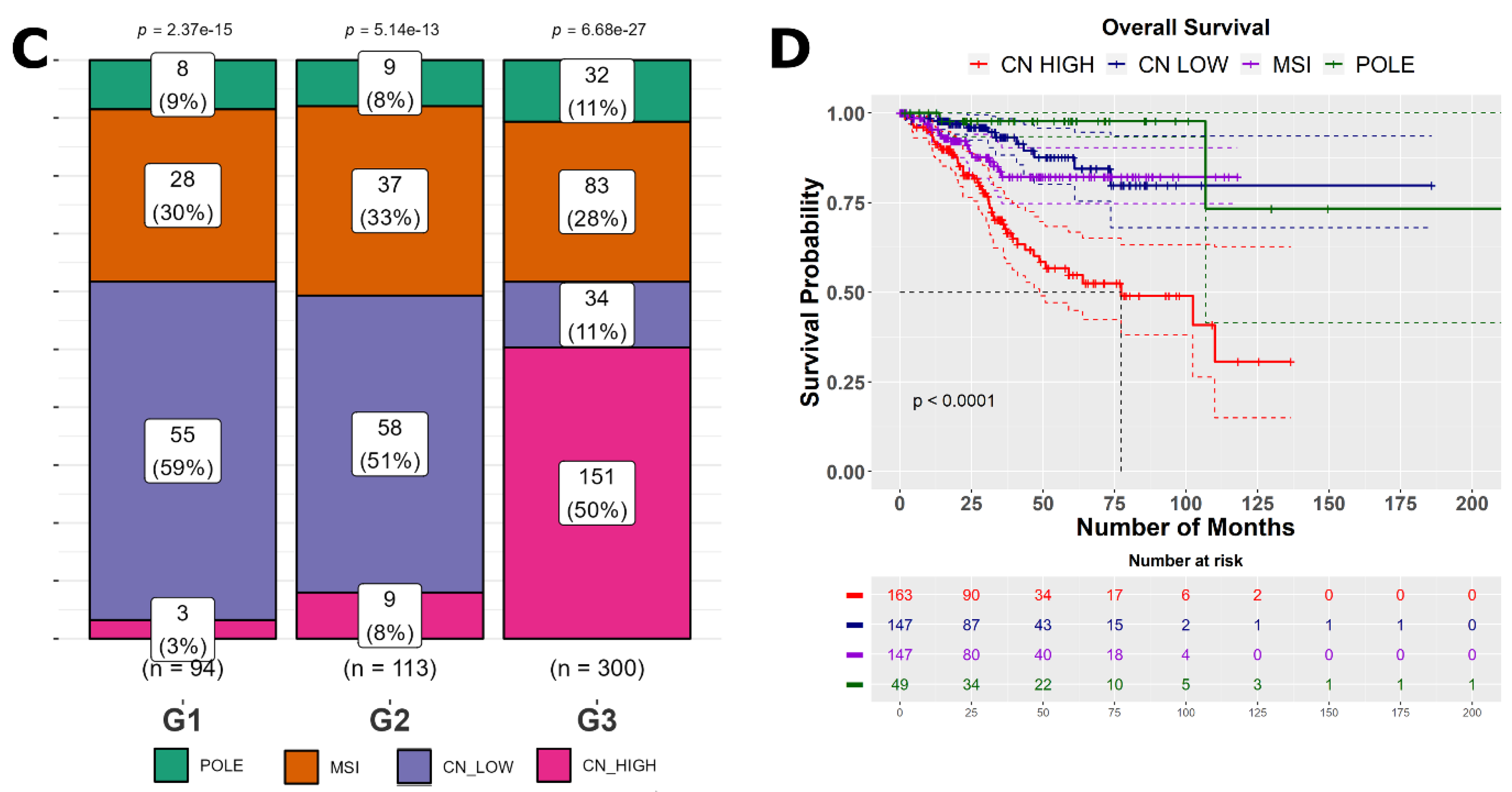
2.4. Clinical Characteristics of SHH High and SHH Low Tumors
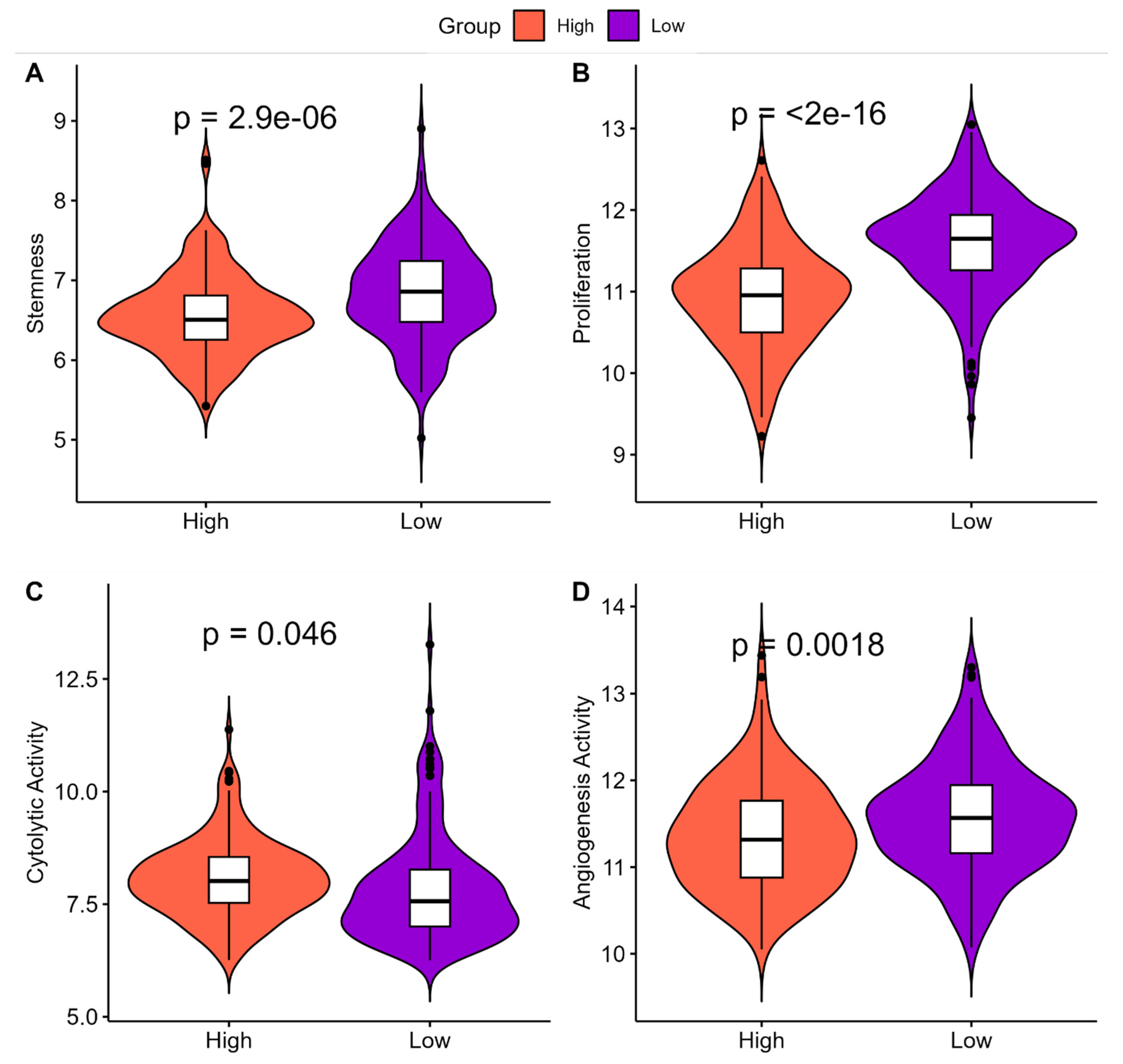
2.5. Understanding the Clinical Implications of ‘High SHH’ and ‘Low SHH’ Tumors
2.6. Genetic Stability and Tumor Mutational Burden in ‘High SHH’ Tumors
2.7. Impact of ‘High SHH’ Tumors on Immune Cell Composition
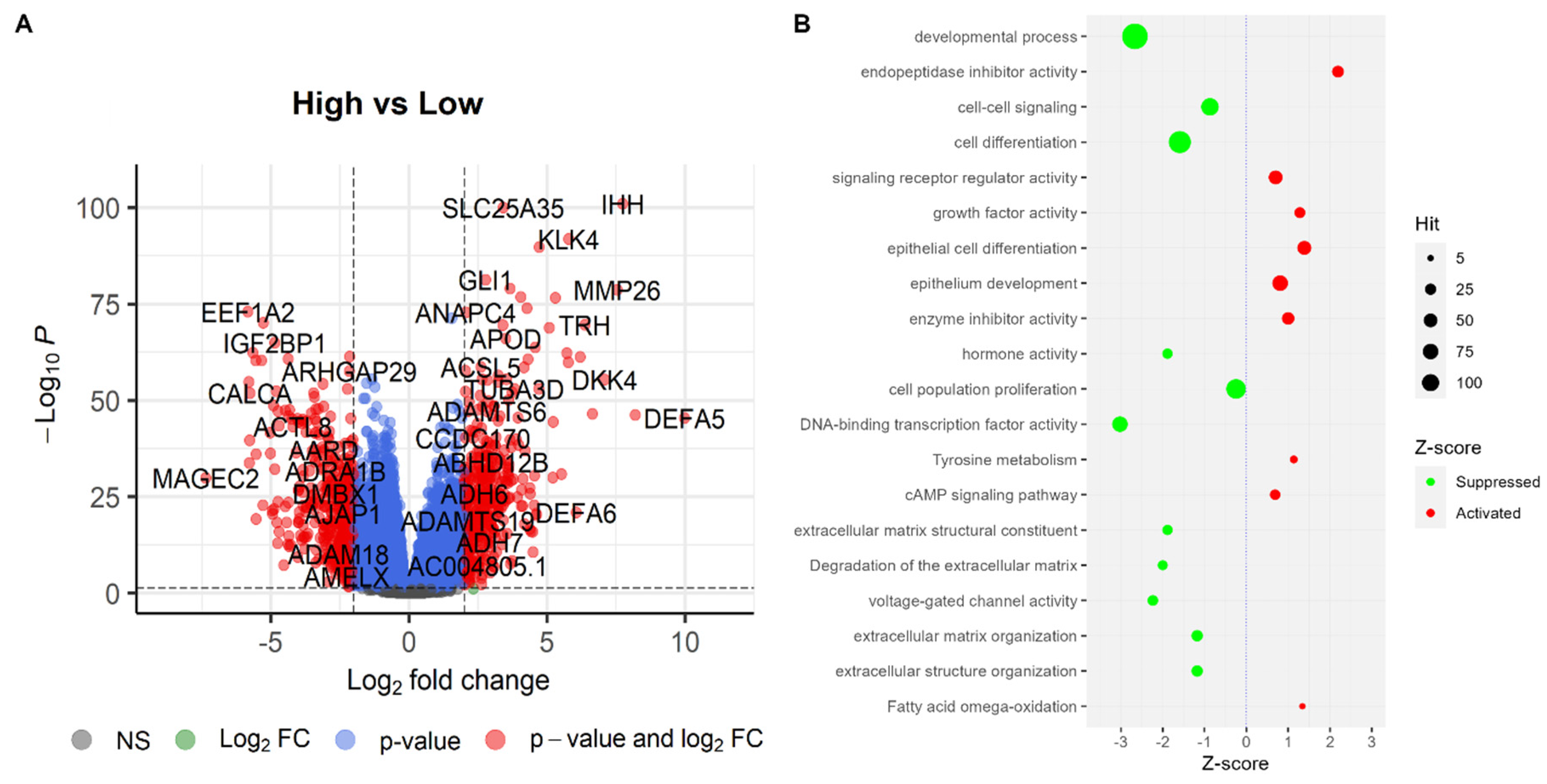
2.8. Differentially Expressed Genes and Pathways
2.9. Impact of Variables on Survival Outcomes in Endometrial Cancer
3. Discussion
4. Materials and Methods
4.1. Collection of Datasets
4.2. Data Processing and Normalization
4.3. Clinical Characteristics
4.4. Cytolytic Activity
4.5. Proliferation and Angiogenesis Score
4.6. Stemness Score
4.7. Genetic Alterations in Tumors
4.8. Identification of Immune Cell Composition
4.9. Identification of Differentially Expressed Genes
4.10. Functional Enrichment Analysis
4.11. Analysing Multiple Variables in EC Survival Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Saccardi, C.; Vitagliano, A.; Marchetti, M.; Lo Turco, A.; Tosatto, S.; Palumbo, M.; De Lorenzo, L.S.; Vitale, S.G.; Scioscia, M.; Noventa, M. Endometrial Cancer Risk Prediction According to Indication of Diagnostic Hysteroscopy in Post-Menopausal Women. Diagnostics 2020, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, V.; Sharma, A.; Parambath, S.V.; Gul, I.; Zhang, X.; Lobie, P.E.; Qin, P.; Pandey, V. Machine Learning for Endometrial Cancer Prediction and Prognostication. Front. Oncol. 2022, 12, 852746. [Google Scholar] [CrossRef]
- Bokhman, J. V Two Pathogenetic Types of Endometrial Carcinoma. Gynecol. Oncol. 1983, 15, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Morice, P.; Leary, A.; Creutzberg, C.; Abu-Rustum, N.; Darai, E. Endometrial Cancer. Lancet 2016, 387, 1094–1108. [Google Scholar] [CrossRef]
- Suarez, A.A.; Felix, A.S.; Cohn, D.E. Bokhman Redux: Endometrial Cancer “Types” in the 21st Century. Gynecol. Oncol. 2017, 144, 243–249. [Google Scholar] [CrossRef]
- Singh, S.; Snijesh, V.P.; Jannet Vennila, J. Rheumatoid Arthritis Candidate Genes Identification by Investigating Core and Periphery Interaction Structures. In Computational Intelligence in Medical Informatics; Muppalaneni, N.B., Gunjan, V.K., Eds.; Springer Singapore: Singapore, 2015; pp. 87–96. ISBN 978-981-287-260-9. [Google Scholar]
- Alsulaimany, F.A.; Zabermawi, N.M.O.; Almukadi, H.; Parambath, S.V.; Shetty, P.J.; Vaidyanathan, V.; Elango, R.; Babanaganapalli, B.; Shaik, N.A. Transcriptome-Based Molecular Networks Uncovered Interplay Between Druggable Genes of CD8+ T Cells and Changes in Immune Cell Landscape in Patients with Pulmonary Tuberculosis. Front. Med. 2021, 8, 812857. [Google Scholar] [CrossRef]
- Bima, A.I.H.; Elsamanoudy, A.Z.; Albaqami, W.F.; Khan, Z.; Parambath, S.V.; Al-Rayes, N.; Kaipa, P.R.; Elango, R.; Banaganapalli, B.; Shaik, N.A. Integrative System Biology and Mathematical Modeling of Genetic Networks Identifies Shared Biomarkers for Obesity and Diabetes. Math. Biosci. Eng. 2022, 19, 2310–2329. [Google Scholar] [CrossRef]
- George, G.; Valiya Parambath, S.; Lokappa, S.B.; Varkey, J. Construction of Parkinson’s Disease Marker-Based Weighted Protein-Protein Interaction Network for Prioritization of Co-Expressed Genes. Gene 2019, 697, 67–77. [Google Scholar] [CrossRef]
- Singh, S.; Vennila, J.J.; Snijesh, V.P.; George, G.; Sunny, C. Implying Analytic Measures for Unravelling Rheumatoid Arthritis Significant Proteins Through Drug-Target Interaction. Interdiscip. Sci. 2016, 8, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Nair, M.G.; Apoorva, D.; Chandrakala, M.; Snijesh, V.P.; Patil, S.; Anupama, C.E.; Mukherjee, G.; Kumar, R.V.; Prabhu, J.S.; Sridhar, T.S. MiR-18a Mediates Immune Evasion in ER-Positive Breast Cancer through Wnt Signaling. Cells 2022, 11, 1672. [Google Scholar] [CrossRef] [PubMed]
- Snijesh, V.P.; Matchado, M.S.; Singh, S. Classifying Rheumatoid Arthritis Gene Network Signatures for Identifying Key Regulatory Molecules and Their Altered Pathways by Adopting Network Biology Approach. Gene Rep. 2018, 13, 199–211. [Google Scholar] [CrossRef]
- Talhouk, A.; McConechy, M.K.; Leung, S.; Li-Chang, H.H.; Kwon, J.S.; Melnyk, N.; Yang, W.; Senz, J.; Boyd, N.; Karnezis, A.N.; et al. A Clinically Applicable Molecular-Based Classification for Endometrial Cancers. Br. J. Cancer 2015, 113, 299–310. [Google Scholar] [CrossRef]
- Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; Cancer Genome Atlas Research Network; et al. Integrated Genomic Characterization of Endometrial Carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.; Balcacer, P.; Castillo, P.; Huang, M.; Alessandrino, F. Endometrial Cancer from Early to Advanced-Stage Disease: An Update for Radiologists. Abdom. Radiol. 2021, 46, 5325–5336. [Google Scholar] [CrossRef]
- Petrov, K.; Wierbowski, B.M.; Salic, A. Sending and Receiving Hedgehog Signals. Annu. Rev. Cell Dev. Biol. 2017, 33, 145–168. [Google Scholar] [CrossRef] [PubMed]
- Carballo, G.B.; Honorato, J.R.; de Lopes, G.P.F.; Spohr, T.C.L. de S.E. A Highlight on Sonic Hedgehog Pathway. Cell Commun. Signal 2018, 16, 11. [Google Scholar] [CrossRef]
- Briscoe, J.; Thérond, P.P. The Mechanisms of Hedgehog Signalling and Its Roles in Development and Disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef]
- Wang, D.; Hu, G.; Du, Y.; Zhang, C.; Lu, Q.; Lv, N.; Luo, S. Aberrant Activation of Hedgehog Signaling Promotes Cell Proliferation via the Transcriptional Activation of Forkhead Box M1 in Colorectal Cancer Cells. J. Exp. Clin. Cancer Res. 2017, 36, 23. [Google Scholar] [CrossRef]
- Hebrok, M.; Kim, S.K.; St Jacques, B.; McMahon, A.P.; Melton, D.A. Regulation of Pancreas Development by Hedgehog Signaling. Development 2000, 127, 4905–4913. [Google Scholar] [CrossRef] [PubMed]
- Kawahira, H.; Ma, N.H.; Tzanakakis, E.S.; McMahon, A.P.; Chuang, P.-T.; Hebrok, M. Combined Activities of Hedgehog Signaling Inhibitors Regulate Pancreas Development. Development 2003, 130, 4871–4879. [Google Scholar] [CrossRef] [PubMed]
- St-Jacques, B.; Hammerschmidt, M.; McMahon, A.P. Indian Hedgehog Signaling Regulates Proliferation and Differentiation of Chondrocytes and Is Essential for Bone Formation. Genes Dev. 1999, 13, 2072–2086. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.H.-C.; Whoriskey, W.; Capel, B. Desert Hedgehog/Patched 1 Signaling Specifies Fetal Leydig Cell Fate in Testis Organogenesis. Genes Dev. 2002, 16, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Rimkus, T.K.; Carpenter, R.L.; Qasem, S.; Chan, M.; Lo, H.-W. Targeting the Sonic Hedgehog Signaling Pathway: Review of Smoothened and GLI Inhibitors. Cancers 2016, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Ingham, P.W.; McMahon, A.P. Hedgehog Signaling in Animal Development: Paradigms and Principles. Genes Dev. 2001, 15, 3059–3087. [Google Scholar] [CrossRef]
- Feng, Y.-Z.; Shiozawa, T.; Miyamoto, T.; Kashima, H.; Kurai, M.; Suzuki, A.; Ying-Song, J.; Konishi, I. Overexpression of Hedgehog Signaling Molecules and Its Involvement in the Proliferation of Endometrial Carcinoma Cells. Clin. Cancer Res. 2007, 13, 1389–1398. [Google Scholar] [CrossRef]
- Inoue, K.; Tsubamoto, H.; Sakata, K.; Sakane, R.; Hao, H.; Hirota, S.; Sonoda, T.; Shibahara, H. Expression of Hedgehog Signals and Growth Inhibition by Itraconazole in Endometrial Cancer. Anticancer Res. 2016, 36, 149–153. [Google Scholar] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Nimbalkar, V.P.; Snijesh, V.P.; Rajarajan, S.; Alexander, A.; Kaluve, R.; Ramesh, R.; Srinath, B.S.; Prabhu, J.S. Premenopausal Women with Breast Cancer in the Early Post-Partum Period Show Molecular Profiles of Invasion and Are Associated with Poor Prognosis. Breast Cancer Res. Treat. 2023, 200, 139–149. [Google Scholar] [CrossRef]
- Jing, J.; Wu, Z.; Wang, J.; Luo, G.; Lin, H.; Fan, Y.; Zhou, C. Hedgehog Signaling in Tissue Homeostasis, Cancers, and Targeted Therapies. Signal Transduct. Target. Ther. 2023, 8, 315. [Google Scholar] [CrossRef] [PubMed]
- Polychronidou, G.; Kotoula, V.; Manousou, K.; Kostopoulos, I.; Karayannopoulou, G.; Vrettou, E.; Bobos, M.; Raptou, G.; Efstratiou, I.; Dionysopoulos, D.; et al. Mismatch Repair Deficiency and Aberrations in the Notch and Hedgehog Pathways Are of Prognostic Value in Patients with Endometrial Cancer. PLoS ONE 2018, 13, e0208221. [Google Scholar] [CrossRef]
- Kim, K.H.; Kim, J.M.; Choi, Y.-L.; Shin, Y.K.; Lee, H.; Seong, I.O.; Kim, B.K.; Chae, S.W.; Chung, Y.-S.; Kim, S.-H. Expression of Sonic Hedgehog Signaling Molecules in Normal, Hyperplastic and Carcinomatous Endometrium. Pathol. Int. 2009, 59, 279–287. [Google Scholar] [CrossRef]
- Snijesh, V.P.; Nimbalkar, V.P.; Patil, S.; Rajarajan, S.; Anupama, C.E.; Mahalakshmi, S.; Alexander, A.; Soundharya, R.; Ramesh, R.; Srinath, B.S.; et al. Differential Role of Glucocorticoid Receptor Based on Its Cell Type Specific Expression on Tumor Cells and Infiltrating Lymphocytes. Transl. Oncol. 2024, 45, 101957. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Li, Y.; Zhang, X. Stemness-Related Markers in Cancer. Cancer Transl. Med. 2017, 3, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Basilio-de-Oliveira, R.P.; Pannain, V.L.N. Prognostic Angiogenic Markers (Endoglin, VEGF, CD31) and Tumor Cell Proliferation (Ki67) for Gastrointestinal Stromal Tumors. World J. Gastroenterol. 2015, 21, 6924–6930. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor Package for Integrative Analysis of TCGA Data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Rajarajan, S.; Snijesh, V.P.; Anupama, C.E.; Nair, M.G.; Mavatkar, A.D.; Naidu, C.M.; Patil, S.; Nimbalkar, V.P.; Alexander, A.; Pillai, M.; et al. An Androgen Receptor Regulated Gene Score Is Associated with Epithelial to Mesenchymal Transition Features in Triple Negative Breast Cancers. Transl. Oncol. 2023, 37, 101761. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor Angiogenesis: Causes, Consequences, Challenges and Opportunities. Cell Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P. VEGF as a Key Mediator of Angiogenesis in Cancer. Oncology 2005, 69 (Suppl. S3), 4–10. [Google Scholar] [CrossRef] [PubMed]
- Giam, M.; Rancati, G. Aneuploidy and Chromosomal Instability in Cancer: A Jackpot to Chaos. Cell Div. 2015, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Chen, S.; Xu, F.; Wei, J.; Zhou, X.; Wu, Z.; Zhao, L.; Liu, J.; Guo, W. Estimating Tumor Mutational Burden across Multiple Cancer Types Using Whole-Exome Sequencing. Ann. Transl. Med. 2021, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust Enumeration of Cell Subsets from Tissue Expression Profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G: Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snijesh, V.P.; Krishnamurthy, S.; Bhardwaj, V.; Punya, K.M.; Niranjana Murthy, A.S.; Almutadares, M.; Habhab, W.T.; Nasser, K.K.; Banaganapalli, B.; Shaik, N.A.; et al. SHH Signaling as a Key Player in Endometrial Cancer: Unveiling the Correlation with Good Prognosis, Low Proliferation, and Anti-Tumor Immune Milieu. Int. J. Mol. Sci. 2024, 25, 10443. https://doi.org/10.3390/ijms251910443
Snijesh VP, Krishnamurthy S, Bhardwaj V, Punya KM, Niranjana Murthy AS, Almutadares M, Habhab WT, Nasser KK, Banaganapalli B, Shaik NA, et al. SHH Signaling as a Key Player in Endometrial Cancer: Unveiling the Correlation with Good Prognosis, Low Proliferation, and Anti-Tumor Immune Milieu. International Journal of Molecular Sciences. 2024; 25(19):10443. https://doi.org/10.3390/ijms251910443
Chicago/Turabian StyleSnijesh, V. P., Shivakumar Krishnamurthy, Vipul Bhardwaj, K. M. Punya, Ashitha S. Niranjana Murthy, Mahmoud Almutadares, Wisam Tahir Habhab, Khalidah Khalid Nasser, Babajan Banaganapalli, Noor Ahmad Shaik, and et al. 2024. "SHH Signaling as a Key Player in Endometrial Cancer: Unveiling the Correlation with Good Prognosis, Low Proliferation, and Anti-Tumor Immune Milieu" International Journal of Molecular Sciences 25, no. 19: 10443. https://doi.org/10.3390/ijms251910443
APA StyleSnijesh, V. P., Krishnamurthy, S., Bhardwaj, V., Punya, K. M., Niranjana Murthy, A. S., Almutadares, M., Habhab, W. T., Nasser, K. K., Banaganapalli, B., Shaik, N. A., & Albaqami, W. F. (2024). SHH Signaling as a Key Player in Endometrial Cancer: Unveiling the Correlation with Good Prognosis, Low Proliferation, and Anti-Tumor Immune Milieu. International Journal of Molecular Sciences, 25(19), 10443. https://doi.org/10.3390/ijms251910443