
Defects in the Maturation of Mitochondrial Iron–Sulfur Proteins: Biophysical Investigation of the MMDS3 Causing Gly104Cys Variant of IBA57
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
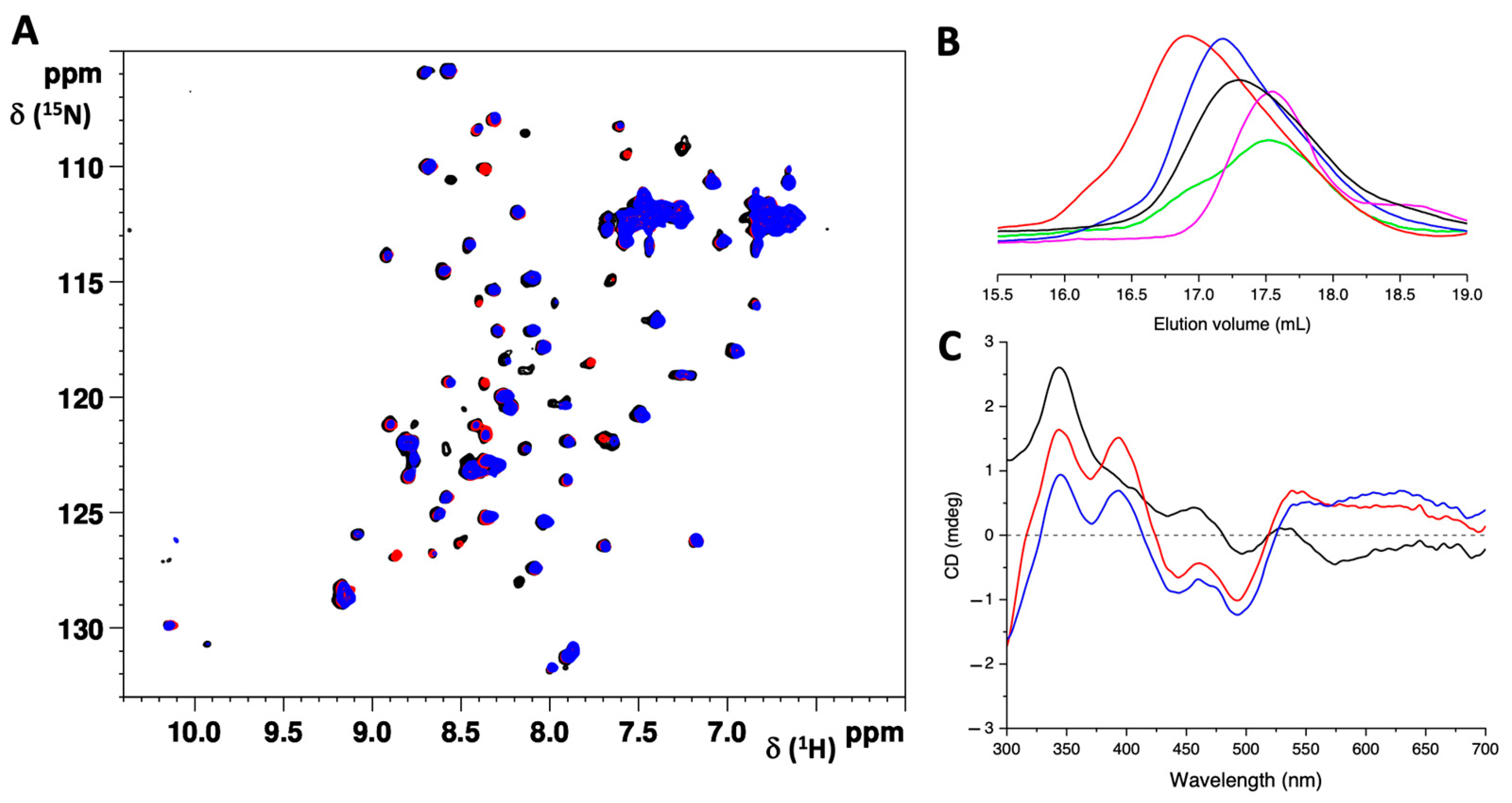
2.1. The G104C Substitution Does Not Significantly Change the Structure of IBA57 but Impacts Its Stability
2.2. G104C Substitution Impacts the Stability of IBA57 Protein
2.3. G104C Mutation of IBA57 Does Not Impair the Interaction with ISCA2 upon [2Fe-2S] Cluster Binding
3. Discussion
4. Materials and Methods
4.1. Proteins Expression and Purification
4.2. In Vitro Chemical Reconstitution of G104C-IBA57
4.3. Biochemical and Spectroscopic CD, Fluorescence and NMR Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lebigot, E.; Schiff, M.; Golinelli-Cohen, M.-P. A Review of Multiple Mitochondrial Dysfunction Syndromes, Syndromes Associated with Defective Fe-S Protein Maturation. Biomedicines 2021, 9, 989. [Google Scholar] [CrossRef] [PubMed]
- Camponeschi, F.; Ciofi-Baffoni, S.; Calderone, V.; Banci, L. Molecular Basis of Rare Diseases Associated to the Maturation of Mitochondrial [4Fe-4S]-Containing Proteins. Biomolecules 2022, 12, 1009. [Google Scholar] [CrossRef] [PubMed]
- Arribas-Carreira, L.; Dallabona, C.; Swanson, M.A.; Farris, J.; Østergaard, E.; Tsiakas, K.; Hempel, M.; Aquaviva-Bourdain, C.; Koutsoukos, S.; Stence, N.V.; et al. Pathogenic Variants in GCSH Encoding the Moonlighting H-Protein Cause Combined Nonketotic Hyperglycinemia and Lipoate Deficiency. Hum. Mol. Genet. 2023, 32, 917–933. [Google Scholar] [CrossRef]
- Gurgel-Giannetti, J.; Lynch, D.S.; de Paiva, A.R.B.; Lucato, L.T.; Yamamoto, G.; Thomsen, C.; Basu, S.; Freua, F.; Giannetti, A.V.; de Assis, B.D.R.; et al. A Novel Complex Neurological Phenotype Due to a Homozygous Mutation in FDX2. Brain 2018, 141, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Masnada, S.; Previtali, R.; Erba, P.; Beretta, E.; Camporesi, A.; Chiapparini, L.; Doneda, C.; Iascone, M.; Sartorio, M.U.A.; Spaccini, L.; et al. FDXR-Associated Disease: A Challenging Differential Diagnosis with Inflammatory Peripheral Neuropathy. Neurol. Sci. 2023, 44, 3037–3043. [Google Scholar] [CrossRef]
- Vögtle, F.-N.; Brändl, B.; Larson, A.; Pendziwiat, M.; Friederich, M.W.; White, S.M.; Basinger, A.; Kücükköse, C.; Muhle, H.; Jähn, J.A.; et al. Mutations in PMPCB Encoding the Catalytic Subunit of the Mitochondrial Presequence Protease Cause Neurodegeneration in Early Childhood. Am. J. Hum. Genet. 2018, 102, 557–573. [Google Scholar] [CrossRef]
- Ciofi-Baffoni, S.; Nasta, V.; Banci, L. Protein Networks in the Maturation of Human Iron–Sulfur Proteins. Metallomics 2018, 10, 49–72. [Google Scholar] [CrossRef]
- Maio, N.; Rouault, T.A. Outlining the Complex Pathway of Mammalian Fe-S Cluster Biogenesis. Trends Biochem. Sci. 2020, 45, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Braymer, J.J.; Lill, R. Iron-Sulfur Cluster Biogenesis and Trafficking in Mitochondria. J. Biol. Chem. 2017, 292, 12754–12763. [Google Scholar] [CrossRef]
- Beinert, H.; Holm, R.H.; Münck, E. Iron-Sulfur Clusters: Nature’s Modular, Multipurpose Structures. Science 1997, 277, 653–659. [Google Scholar] [CrossRef]
- Lill, R. Function and Biogenesis of Iron-Sulphur Proteins. Nature 2009, 460, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Rouault, T.A. Mammalian Iron-Sulphur Proteins: Novel Insights into Biogenesis and Function. Nat. Rev. Mol. Cell Biol. 2015, 16, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Boniecki, M.T.; Freibert, S.A.; Mühlenhoff, U.; Lill, R.; Cygler, M. Structure and Functional Dynamics of the Mitochondrial Fe/S Cluster Synthesis Complex. Nat. Commun. 2017, 8, 1287. [Google Scholar] [CrossRef] [PubMed]
- Cory, S.A.; Van Vranken, J.G.; Brignole, E.J.; Patra, S.; Winge, D.R.; Drennan, C.L.; Rutter, J.; Barondeau, D.P. Structure of Human Fe-S Assembly Subcomplex Reveals Unexpected Cysteine Desulfurase Architecture and Acyl-ACP-ISD11 Interactions. Proc. Natl. Acad. Sci. USA 2017, 114, E5325–E5334. [Google Scholar] [CrossRef]
- Van Vranken, J.G.; Jeong, M.-Y.; Wei, P.; Chen, Y.-C.; Gygi, S.P.; Winge, D.R.; Rutter, J. The Mitochondrial Acyl Carrier Protein (ACP) Coordinates Mitochondrial Fatty Acid Synthesis with Iron Sulfur Cluster Biogenesis. eLife 2016, 5, e17828. [Google Scholar] [CrossRef]
- Freibert, S.-A.; Boniecki, M.T.; Stümpfig, C.; Schulz, V.; Krapoth, N.; Winge, D.R.; Mühlenhoff, U.; Stehling, O.; Cygler, M.; Lill, R. N-Terminal Tyrosine of ISCU2 Triggers [2Fe-2S] Cluster Synthesis by ISCU2 Dimerization. Nat. Commun. 2021, 12, 6902. [Google Scholar] [CrossRef]
- Sheftel, A.D.; Stehling, O.; Pierik, A.J.; Elsässer, H.-P.; Mühlenhoff, U.; Webert, H.; Hobler, A.; Hannemann, F.; Bernhardt, R.; Lill, R. Humans Possess Two Mitochondrial Ferredoxins, Fdx1 and Fdx2, with Distinct Roles in Steroidogenesis, Heme, and Fe/S Cluster Biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 11775–11780. [Google Scholar] [CrossRef]
- Cai, K.; Tonelli, M.; Frederick, R.O.; Markley, J.L. Human Mitochondrial Ferredoxin 1 (FDX1) and Ferredoxin 2 (FDX2) Both Bind Cysteine Desulfurase and Donate Electrons for Iron-Sulfur Cluster Biosynthesis. Biochemistry 2017, 56, 487–499. [Google Scholar] [CrossRef]
- Gervason, S.; Larkem, D.; Mansour, A.B.; Botzanowski, T.; Müller, C.S.; Pecqueur, L.; Le Pavec, G.; Delaunay-Moisan, A.; Brun, O.; Agramunt, J.; et al. Physiologically Relevant Reconstitution of Iron-Sulfur Cluster Biosynthesis Uncovers Persulfide-Processing Functions of Ferredoxin-2 and Frataxin. Nat. Commun. 2019, 10, 3566. [Google Scholar] [CrossRef]
- Weiler, B.D.; Brück, M.-C.; Kothe, I.; Bill, E.; Lill, R.; Mühlenhoff, U. Mitochondrial [4Fe-4S] Protein Assembly Involves Reductive [2Fe-2S] Cluster Fusion on ISCA1-ISCA2 by Electron Flow from Ferredoxin FDX2. Proc. Natl. Acad. Sci. USA 2020, 117, 20555–20565. [Google Scholar] [CrossRef]
- Sheftel, A.D.; Wilbrecht, C.; Stehling, O.; Niggemeyer, B.; Elsässer, H.-P.; Mühlenhoff, U.; Lill, R. The Human Mitochondrial ISCA1, ISCA2, and IBA57 Proteins Are Required for [4Fe-4S] Protein Maturation. Mol. Biol. Cell 2012, 23, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, D.; Gallo, A.; Mikolajczyk, M.; Zovo, K.; Palumaa, P.; Novellino, E.; Piccioli, M.; Ciofi-Baffoni, S.; Banci, L. Formation of [4Fe-4S] Clusters in the Mitochondrial Iron-Sulfur Cluster Assembly Machinery. J. Am. Chem. Soc. 2014, 136, 16240–16250. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Brancaccio, D.; Ciofi-Baffoni, S.; Del Conte, R.; Gadepalli, R.; Mikolajczyk, M.; Neri, S.; Piccioli, M.; Winkelmann, J. [2Fe-2S] Cluster Transfer in Iron-Sulfur Protein Biogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 6203–6208. [Google Scholar] [CrossRef] [PubMed]
- Braymer, J.J.; Freibert, S.A.; Rakwalska-Bange, M.; Lill, R. Mechanistic Concepts of Iron-Sulfur Protein Biogenesis in Biology. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118863. [Google Scholar] [CrossRef] [PubMed]
- Mayr, J.A.; Feichtinger, R.G.; Tort, F.; Ribes, A.; Sperl, W. Lipoic Acid Biosynthesis Defects. J. Inherit. Metab. Dis. 2014, 37, 553–563. [Google Scholar] [CrossRef]
- Zhan, F.; Liu, X.; Ni, R.; Liu, T.; Cao, Y.; Wu, J.; Tian, W.; Luan, X.; Cao, L. Novel IBA57 Mutations in Two Chinese Patients and Literature Review of Multiple Mitochondrial Dysfunction Syndrome. Metab. Brain Dis. 2021, 37, 311–317. [Google Scholar] [CrossRef]
- Lang, S.H.; Camponeschi, F.; de Joya, E.; Borjas-Mendoza, P.; Tekin, M.; Thorson, W. Multiple Mitochondrial Dysfunction Syndrome Type 3: A Likely Pathogenic Homozygous Variant Affecting a Patient of Cuban Descent and Literature Review. Genes. 2022, 13, 2044. [Google Scholar] [CrossRef]
- Kristina Beilschmidt, L.; Choudens, S.; Fournier, M.; Sanakis, Y.; Hograindleur, M.-A.; Clémancey, M.; Blondin, G.; Schmucker, S.; Eisenmann, A.; Weiss, A.; et al. ISCA1 Is Essential for Mitochondrial Fe4S4 Biogenesis In Vivo. Nat. Commun. 2017, 8, 15124. [Google Scholar] [CrossRef]
- Brancaccio, D.; Gallo, A.; Piccioli, M.; Novellino, E.; Ciofi-Baffoni, S.; Banci, L. [4Fe-4S] Cluster Assembly in Mitochondria and Its Impairment by Copper. J. Am. Chem. Soc. 2017, 139, 719–730. [Google Scholar] [CrossRef]
- Roche, B.; Aussel, L.; Ezraty, B.; Mandin, P.; Py, B.; Barras, F. Iron/Sulfur Proteins Biogenesis in Prokaryotes: Formation, Regulation and Diversity. Biochim. Biophys. Acta 2013, 1827, 455–469. [Google Scholar] [CrossRef]
- Gourdoupis, S.; Nasta, V.; Calderone, V.; Ciofi-Baffoni, S.; Banci, L. IBA57 Recruits ISCA2 to Form a [2Fe-2S] Cluster-Mediated Complex. J. Am. Chem. Soc. 2018, 140, 14401–14412. [Google Scholar] [CrossRef] [PubMed]
- Nasta, V.; Da Vela, S.; Gourdoupis, S.; Ciofi-Baffoni, S.; Svergun, D.I.; Banci, L. Structural Properties of [2Fe-2S] ISCA2-IBA57: A Complex of the Mitochondrial Iron-Sulfur Cluster Assembly Machinery. Sci. Rep. 2019, 9, 18986. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Aoki, R.; Nagano, N.; Takano, C.; Seimiya, A.; Kato, R.; Ogawa, E.; Ishige, M.; Okazaki, Y.; Murayama, K.; et al. Unique and Abnormal Subependymal Pseudocysts in a Newborn with Mitochondrial Disease. Sci. Prog. 2021, 104, 368504211011873. [Google Scholar] [CrossRef] [PubMed]
- Gourdoupis, S.; Nasta, V.; Ciofi-Baffoni, S.; Banci, L.; Calderone, V. In-House High-Energy-Remote SAD Phasing Using the Magic Triangle: How to Tackle the P1 Low Symmetry Using Multiple Orientations of the Same Crystal of Human IBA57 to Increase the Multiplicity. Acta Crystallogr. D Struct. Biol. 2019, 75, 317–324. [Google Scholar] [CrossRef]
- Micsonai, A.; Wien, F.; Kernya, L.; Lee, Y.-H.; Goto, Y.; Réfrégiers, M.; Kardos, J. Accurate Secondary Structure Prediction and Fold Recognition for Circular Dichroism Spectroscopy. Proc. Natl. Acad. Sci. USA 2015, 112, E3095–E3103. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using Circular Dichroism Collected as a Function of Temperature to Determine the Thermodynamics of Protein Unfolding and Binding Interactions. Nat. Protoc. 2006, 1, 2527–2535. [Google Scholar] [CrossRef]
- Royer, C.A. Probing Protein Folding and Conformational Transitions with Fluorescence. Chem. Rev. 2006, 106, 1769–1784. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tsai, C.-J.; Nussinov, R. Maximal Stabilities of Reversible Two-State Proteins. Biochemistry 2002, 41, 5359–5374. [Google Scholar] [CrossRef]
- Biederbick, A.; Stehling, O.; Rösser, R.; Niggemeyer, B.; Nakai, Y.; Elsässer, H.-P.; Lill, R. Role of Human Mitochondrial Nfs1 in Cytosolic Iron-Sulfur Protein Biogenesis and Iron Regulation. Mol. Cell. Biol. 2006, 26, 5675–5687. [Google Scholar] [CrossRef]
- Sheftel, A.D.; Stehling, O.; Pierik, A.J.; Netz, D.J.A.; Kerscher, S.; Elsässer, H.-P.; Wittig, I.; Balk, J.; Brandt, U.; Lill, R. Human Ind1, an Iron-Sulfur Cluster Assembly Factor for Respiratory Complex I. Mol. Cell Biol. 2009, 29, 6059–6073. [Google Scholar] [CrossRef]
- Mühlenhoff, U.; Richter, N.; Pines, O.; Pierik, A.J.; Lill, R. Specialized Function of Yeast Isa1 and Isa2 Proteins in the Maturation of Mitochondrial [4Fe-4S] Proteins. J. Biol. Chem. 2011, 286, 41205–41216. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, D.H. Chemical Chaperones: A Pharmacological Strategy for Disorders of Protein Folding and Trafficking. Pediatr. Res. 2002, 52, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Cortez, L.; Sim, V. The Therapeutic Potential of Chemical Chaperones in Protein Folding Diseases. Prion 2014, 8, 197–202. [Google Scholar] [CrossRef]
- Liguori, L.; Monticelli, M.; Allocca, M.; Hay Mele, B.; Lukas, J.; Cubellis, M.V.; Andreotti, G. Pharmacological Chaperones: A Therapeutic Approach for Diseases Caused by Destabilizing Missense Mutations. Int. J. Mol. Sci. 2020, 21, 489. [Google Scholar] [CrossRef]
- Chen, J.; Acton, T.B.; Basu, S.K.; Montelione, G.T.; Inouye, M. Enhancement of the Solubility of Proteins Overexpressed in Escherichia Coli by Heat Shock. J. Mol. Microbiol. Biotechnol. 2002, 4, 519–524. [Google Scholar] [PubMed]
- Case, D.A.; Cheatham, T.E., III; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber Biomolecular Simulation Programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef]
- Salomon-Ferrer, R.; Case, D.A.; Walker, R.C. An Overview of the Amber Biomolecular Simulation Package. WIREs Comput. Mol. Sci. 2013, 3, 198–210. [Google Scholar] [CrossRef]
- Svilenov, H.; Markoja, U.; Winter, G. Isothermal Chemical Denaturation as a Complementary Tool to Overcome Limitations of Thermal Differential Scanning Fluorimetry in Predicting Physical Stability of Protein Formulations. Eur. J. Pharm. Biopharm. 2018, 125, 106–113. [Google Scholar] [CrossRef]
- Freire, E.; Schön, A.; Hutchins, B.M.; Brown, R.K. Chemical Denaturation as a Tool in the Formulation Optimization of Biologics. Drug Discov. Today 2013, 18, 1007–1013. [Google Scholar] [CrossRef]
- Kurtin, W.E.; Lee, J.M. The Free Energy of Denaturation of Lysozyme: An Undergraduate Experiment in Biophysical Chemistry. Biochem. Mol. Biol. Educ. 2002, 30, 244–247. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | α-Helical | Antiparallel β-Strand | Parallel β-Strand | Turn | Others a |
|---|---|---|---|---|---|
| G104C-IBA57 | 9.3% | 25.4% | 1.8% | 13.4% | 50.1% |
| WT-IBA57 | 10.0% | 26.2% | 0.5% | 14.1% | 49.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bargagna, B.; Staderini, T.; Lang, S.H.; Banci, L.; Camponeschi, F. Defects in the Maturation of Mitochondrial Iron–Sulfur Proteins: Biophysical Investigation of the MMDS3 Causing Gly104Cys Variant of IBA57. Int. J. Mol. Sci. 2024, 25, 10466. https://doi.org/10.3390/ijms251910466
Bargagna B, Staderini T, Lang SH, Banci L, Camponeschi F. Defects in the Maturation of Mitochondrial Iron–Sulfur Proteins: Biophysical Investigation of the MMDS3 Causing Gly104Cys Variant of IBA57. International Journal of Molecular Sciences. 2024; 25(19):10466. https://doi.org/10.3390/ijms251910466
Chicago/Turabian StyleBargagna, Beatrice, Tommaso Staderini, Steven H. Lang, Lucia Banci, and Francesca Camponeschi. 2024. "Defects in the Maturation of Mitochondrial Iron–Sulfur Proteins: Biophysical Investigation of the MMDS3 Causing Gly104Cys Variant of IBA57" International Journal of Molecular Sciences 25, no. 19: 10466. https://doi.org/10.3390/ijms251910466