
Comparative Analysis of the Cutaneous Microbiome in Psoriasis Patients and Healthy Individuals—Insights into Microbial Dysbiosis: Final Results
 ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michalek, I.M.; Loring, B.; John, S.M. A Systematic Review of Worldwide Epidemiology of Psoriasis. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Hölsken, S.; Krefting, F.; Schedlowski, M.; Sondermann, W. Common Fundamentals of Psoriasis and Depression. Acta Derm. Venereol. 2021, 101, adv00609. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, F. Psoriasis: Comorbidities. J. Dermatol. 2021, 48, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Barros, G.; Duran, P.; Vera, I.; Bermúdez, V. Exploring the Links between Obesity and Psoriasis: A Comprehensive Review. Int. J. Mol. Sci. 2022, 23, 7499. [Google Scholar] [CrossRef]
- Gill, P.A.; Inniss, S.; Kumagai, T.; Rahman, F.Z.; Smith, A.M. The Role of Diet and Gut Microbiota in Regulating Gastrointestinal and Inflammatory Disease. Front. Immunol. 2022, 13, 866059. [Google Scholar] [CrossRef]
- Lederberg, J. Ome Sweet’Omics--A Genealogical Treasury of Words. Scientist 2001, 15, 8. [Google Scholar]
- Roth, R.R.; James, W.D. Microbial ecology of the skin. Annu. Rev. Microbiol. 1988, 42, 441–464. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, L.S.; Dixit, S.; Farrer, A.G.; Cooper, A.J.; Cooper, A.J. The Skin Microbiome: Associations between Altered Microbial Communities and Disease. Australas. J. Dermatol. 2015, 56, 268–274. [Google Scholar] [CrossRef]
- McFadden, J.; Valdimarsson, H.; Fry, L. Cross-Reactivity between Streptococcal M Surface Antigen and Human Skin. Br. J. Dermatol. 1991, 125, 443–447. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Fry, L.; Baker, B.S.; Powles, A.V.; Fahlen, A.; Engstrand, L. Is Chronic Plaque Psoriasis Triggered by Microbiota in the Skin? Br. J. Dermatol. 2013, 169, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Cantabrana, C.; Gómez, J.; Delgado, S.; Requena-López, S.; Queiro-Silva, R.; Margolles, A.; Coto, E.; Sánchez, B.; Coto-Segura, P. Gut Microbiota Dysbiosis in a Cohort of Patients with Psoriasis. Br. J. Dermatol. 2019, 181, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shi, L.; Sun, T.; Guo, K.; Geng, S. Dysbiosis of Gut Microbiota and Its Correlation with Dysregulation of Cytokines in Psoriasis Patients. BMC Microbiol. 2021, 21, 78. [Google Scholar] [CrossRef] [PubMed]
- Schade, L.; Mesa, D.; Faria, A.R.; Santamaria, J.R.; Xavier, C.A.; Ribeiro, D.; Hajar, F.N.; Azevedo, V.F. The Gut Microbiota Profile in Psoriasis: A Brazilian Case-Control Study. Lett. Appl. Microbiol. 2022, 74, 498–504. [Google Scholar] [CrossRef] [PubMed]
- Paulino, L.C.; Tseng, C.H.; Strober, B.E.; Blaser, M.J. Molecular Analysis of Fungal Microbiota in Samples from Healthy Human Skin and Psoriatic Lesions. J. Clin. Microbiol. 2006, 44, 2933–2941. [Google Scholar] [CrossRef]
- Chen, L.; Li, J.; Zhu, W.; Kuang, Y.; Liu, T.; Zhang, W.; Chen, X.; Peng, C. Skin and Gut Microbiome in Psoriasis: Gaining Insight into the Pathophysiology of It and Finding Novel Therapeutic Strategies. Front. Microbiol. 2020, 11, 589726. [Google Scholar] [CrossRef]
- Mazur, M.; Tomczak, H.; Lodyga, M.; Czajkowski, R.; Żaba, R.; Adamski, Z. The microbiome of the human skin and its variability in psoriasis and atopic dermatitis. Postepy Dermatol. Alergol. 2021, 38, 205–209. [Google Scholar] [CrossRef]
- Chen, Y.; Knight, R.; Gallo, R.L. Evolving Approaches to Profiling the Microbiome in Skin Disease. Front. Immunol. 2023, 14, 1151527. [Google Scholar] [CrossRef]
- Dréno, B.; Araviiskaia, E.; Berardesca, E.; Gontijo, G.; Sanchez Viera, M.; Xiang, L.F.; Martin, R.; Bieber, T. Microbiome in healthy skin, update for dermatologists. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 2038–2047. [Google Scholar] [CrossRef]
- Zheng, Y.; Hunt, R.L.; Villaruz, A.E.; Fisher, E.L.; Liu, R.; Liu, Q.; Cheung, G.Y.C.; Li, M.; Otto, M. Commensal Staphylococcus epidermidis Contributes to Skin Barrier Homeostasis by Generating Protective Ceramides. Cell Host Microbe 2022, 30, 301–313.e9. [Google Scholar] [CrossRef]
- Bomar, L.; Brugger, S.D.; Yost, B.H.; Davies, S.S.; Lemon, K.P. Corynebacterium accolens Releases Antipneumococcal Free Fatty Acids from Human Nostril and Skin Surface Triacylglycerols. mBio 2016, 7, e01725-15. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Bouladoux, N.; Claesen, J.; Chen, Y.E.; Byrd, A.L.; Constantinides, M.G.; Merrill, E.D.; Tamoutounour, S.; Fischbach, M.A.; Belkaid, Y. Contextual Control of Skin Immunity and Inflammation by Corynebacterium. J. Exp. Med. 2018, 215, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Kim, M. Skin Barrier Function and the Microbiome. Int. J. Mol. Sci. 2022, 23, 13071. [Google Scholar] [CrossRef] [PubMed]
- Chehoud, C.; Rafail, S.; Tyldsley, A.S.; Seykora, J.T.; Lambris, J.D.; Grice, E.A. Complement Modulates the Cutaneous Microbiome and Inflammatory Milieu. Proc. Natl. Acad. Sci. USA 2013, 110, 15061–15066. [Google Scholar] [CrossRef]
- Huang, S.; Hon, K.; Bennett, C.; Hu, H.; Menberu, M.; Wormald, P.J.; Zhao, Y.; Vreugde, S.; Liu, S. Corynebacterium accolens Inhibits Staphylococcus aureus Induced Mucosal Barrier Disruption. Front. Microbiol. 2022, 13, 984741. [Google Scholar] [CrossRef]
- Lee, J.S.; Tato, C.M.; Joyce-Shaikh, B.; Gulen, M.F.; Cayatte, C.; Chen, Y.; Blumenschein, W.M.; Judo, M.; Ayanoglu, G.; McClanahan, T.K.; et al. Interleukin-23-Independent IL-17 Production Regulates Intestinal Epithelial Permeability. Immunity 2015, 43, 727–738. [Google Scholar] [CrossRef]
- Harrison, O.J.; Linehan, J.L.; Shih, H.Y.; Bouladoux, N.; Han, S.J.; Smelkinson, M.; Sen, S.K.; Byrd, A.L.; Enamorado, M.; Yao, C.; et al. Commensal-Specific T Cell Plasticity Promotes Rapid Tissue Adaptation to Injury. Science 2019, 363, eaat6280. [Google Scholar] [CrossRef]
- Fahlen, A.; Engstrand, L.; Baker, B.S.; Powles, A.; Fry, L. Comparison of Bacterial Microbiota in Skin Biopsies from Normal and Psoriatic Skin. Arch. Dermatol. Res. 2012, 304, 15–22. [Google Scholar] [CrossRef]
- Chang, H.W.; Yan, D.; Singh, R.; Liu, J.; Lu, X.; Ucmak, D.; Lee, K.; Afifi, L.; Fadrosh, D.; Leech, J.; et al. Alteration of the Cutaneous Microbiome in Psoriasis and Potential Role in Th17 Polarization. Microbiome 2018, 6, 154. [Google Scholar] [CrossRef]
- Alekseyenko, A.V.; Perez-Perez, G.I.; De Souza, A.; Strober, B.; Gao, Z.; Bihan, M.; Li, K.; Methé, B.A.; Blaser, M.J. Community Differentiation of the Cutaneous Microbiota in Psoriasis. Microbiome 2013, 1, 31. [Google Scholar] [CrossRef]
- Yang, Y.; Olah, P.; Radai, Z.; Maia, G.; Salava, A.; Salo, V.; Barker, J.; Lauerma, A.; Andersson, B.; Homey, B.; et al. Exploratory Multi-Omics Analysis Reveals Host-Microbe Interactions Associated with Disease Severity in Psoriatic Skin. EBioMedicine 2024, 105, 105222. [Google Scholar] [CrossRef] [PubMed]
- Thio, H.B. The Microbiome in Psoriasis and Psoriatic Arthritis: The Skin Perspective. J. Rheumatol. Suppl. 2018, 94, 30–31. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.J.E.; Kell, D.B.; Pretorius, E. Bacterial Dysbiosis and Translocation in Psoriasis Vulgaris. Front. Cell. Infect. Microbiol. 2019, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Fyhrquist, N.; Muirhead, G.; Prast-Nielsen, S.; Jeanmougin, M.; Olah, P.; Skoog, T.; Jules-Clement, G.; Feld, M.; Barrientos-Somarribas, M.; Sinkko, H.; et al. Microbe-Host Interplay in Atopic Dermatitis and Psoriasis. Nat. Commun. 2019, 10, 4703. [Google Scholar] [CrossRef] [PubMed]
- Quan, C.; Chen, X.Y.; Li, X.; Xue, F.; Chen, L.H.; Liu, N.; Wang, B.; Wang, L.Q.; Wang, X.P.; Yang, H.; et al. Psoriatic Lesions Are Characterized by Higher Bacterial Load and Imbalance between Cutibacterium and Corynebacterium. J. Am. Acad. Dermatol. 2020, 82, 955–961. [Google Scholar] [CrossRef]
- Drago, L.; De Grandi, R.; Altomare, G.; Pigatto, P.; Rossi, O.; Toscano, M. Skin Microbiota of First Cousins Affected by Psoriasis and Atopic Dermatitis. Clin. Mol. Allergy 2016, 14, 2. [Google Scholar] [CrossRef]
- Gao, Z.; Tseng, C.H.; Strober, B.E.; Pei, Z.; Blaser, M.J. Substantial Alterations of the Cutaneous Bacterial Biota in Psoriatic Lesions. PLoS ONE 2008, 3, e2719. [Google Scholar] [CrossRef]
- Tett, A.; Pasolli, E.; Farina, S.; Truong, D.T.; Asnicar, F.; Zolfo, M.; Beghini, F.; Armanini, F.; Jousson, O.; De Sanctis, V.; et al. Unexplored Diversity and Strain-Level Structure of the Skin Microbiome Associated with Psoriasis. NPJ Biofilms Microbiomes 2017, 3, 14. [Google Scholar] [CrossRef]
- Liu, S.H.; Yu, H.Y.; Chang, Y.C.; Chung-Yee Hui, R.; Huang, Y.C.; Huang, Y.H. Host Characteristics and Dynamics of Staphylococcus aureus Colonization in Patients with Moderate-to-Severe Psoriasis Before and After Treatment: A Prospective Cohort Study. J. Am. Acad. Dermatol. 2019, 81, 605–607. [Google Scholar] [CrossRef]
- Ng, C.Y.; Huang, Y.H.; Chu, C.F.; Wu, T.C.; Liu, S.H. Risks for Staphylococcus aureus Colonization in Patients with Psoriasis: A Systematic Review and Meta-Analysis. Br. J. Dermatol. 2017, 177, 967–977. [Google Scholar] [CrossRef]
- Ovčina-Kurtović, N.; Kasumagić-Halilović, E.; Helppikangans, H.; Begić, J. Prevalence of Candida Species in Patients with Psoriasis. Acta Dermatovenerol. Croat. 2016, 24, 209–213. [Google Scholar] [PubMed]
- Lewis, D.J.; Chan, W.H.; Hinojosa, T.; Hsu, S.; Feldman, S.R. Mechanisms of microbial pathogenesis and the role of the skin microbiome in psoriasis: A review. Clin. Dermatol. 2019, 37, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Hurabielle, C.; Link, V.M.; Bouladoux, N.; Han, S.J.; Merrill, E.D.; Lightfoot, Y.L.; Seto, N.; Bleck, C.K.; Smelkinson, M.; Harrison, O.J.; et al. Immunity to commensal skin fungi promotes psoriasiform skin inflammation. Proc. Natl. Acad. Sci. USA 2020, 117, 16465–16474. [Google Scholar] [CrossRef] [PubMed]
- Saunte, D.M.; Mrowietz, U.; Puig, L.; Zachariae, C. Candida Infections in Patients with Psoriasis and Psoriatic Arthritis Treated with Interleukin-17 Inhibitors and Their Practical Management. Br. J. Dermatol. 2017, 177, 47–62. [Google Scholar] [CrossRef]
- Waldman, A.; Gilhar, A.; Duek, L.; Berdicevsky, I. Incidence of Candida in Psoriasis—A Study on the Fungal Flora of Psoriatic Patients. Mycoses 2001, 44, 77–81. [Google Scholar] [CrossRef]
- Hashiguchi, Y.; Yabe, R.; Chung, S.H.; Murayama, M.A.; Yoshida, K.; Matsuo, K.; Kubo, S.; Saijo, S.; Nakamura, Y.; Matsue, H.; et al. IL-36α from Skin-Resident Cells Plays an Important Role in the Pathogenesis of Imiquimod-Induced Psoriasiform Dermatitis by Forming a Local Autoamplification Loop. J. Immunol. 2018, 201, 167–182. [Google Scholar] [CrossRef]
- Kaminsky, A. Comparative Effects of Griseofulvin on Onychomycosis and on Psoriasis Unguium. AMA Arch. Dermatol. 1960, 81, 838–840. [Google Scholar] [CrossRef]
- Grice, E.A.; Kong, H.H.; Conlan, S.; Deming, C.B.; Davis, J.; Young, A.C.; NISC Comparative Sequencing Program; Bouffard, G.G.; Blakesley, R.W.; Murray, P.R.; et al. Topographical and Temporal Diversity of the Human Skin Microbiome. Science 2009, 324, 1190–1192. [Google Scholar] [CrossRef]
- Oh, J.; Byrd, A.L.; Deming, C.; Conlan, S.; NISC Comparative Sequencing Program; Kong, H.H.; Segre, J.A. Biogeography and Individuality Shape Function in the Human Skin Metagenome. Nature 2014, 514, 59–64. [Google Scholar] [CrossRef]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; NIH Intramural Sequencing Center Comparative Sequencing Program; Schoenfeld, D.; Nomicos, E.; Park, M.; et al. Topographic Diversity of Fungal and Bacterial Communities in Human Skin. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef]
- Boxberger, M.; Cenizo, V.; Cassir, N.; La Scola, B. Challenges in Exploring and Manipulating the Human Skin Microbiome. Microbiome 2021, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Forton, F.M.N.; De Maertelaer, V. Which Factors Influence Demodex Proliferation? A Retrospective Pilot Study Highlighting a Possible Role of Subtle Immune Variations and Sebaceous Gland Status. J. Dermatol. 2021, 48, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Lousada, M.B.; Edelkamp, J.; Lachnit, T.; Fehrholz, M.; Pastar, I.; Jimenez, F.; Erdmann, H.; Bosch, T.C.G.; Paus, R. Spatial Distribution and Functional Impact of Human Scalp Hair Follicle Microbiota. J. Investig. Dermatol. 2024, 144, 1353–1367.e15. [Google Scholar] [CrossRef]
- Wang, X.; Chen, H.; Tian, R.; Zhang, Y.; Drutskaya, M.S.; Wang, C.; Ge, J.; Fan, Z.; Kong, D.; Wang, X.; et al. Macrophages Induce AKT/β-Catenin-Dependent Lgr5+ Stem Cell Activation and Hair Follicle Regeneration through TNF. Nat. Commun. 2017, 8, 14091. [Google Scholar] [CrossRef]
- Rahmani, W.; Liu, Y.; Rosin, N.L.; Kline, A.; Raharjo, E.; Yoon, J.; Stratton, J.A.; Sinha, S.; Biernaskie, J. Macrophages Promote Wound-Induced Hair Follicle Regeneration in a CX3CR1- and TGF-β1-Dependent Manner. J. Investig. Dermatol. 2018, 138, 2111–2122. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, X.; Han, J.; Fan, Z.; Sadia, S.; Zhang, R.; Guo, Y.; Jiang, Y.; Wu, Y. AKT and Its Related Molecular Feature in Aged Mice Skin. PLoS ONE 2017, 12, e0178969. [Google Scholar] [CrossRef]
- Robert, C.; Cascella, F.; Mellai, M.; Barizzone, N.; Mignone, F.; Massa, N.; Nobile, V.; Bona, E. Influence of Sex on the Microbiota of the Human Face. Microorganisms 2022, 10, 2470. [Google Scholar] [CrossRef]
- Skowron, K.; Bauza-Kaszewska, J.; Kraszewska, Z.; Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Kwiecińska-Piróg, J.; Wałecka-Zacharska, E.; Radtke, L.; Gospodarek-Komkowska, E. Human Skin Microbiome: Impact of Intrinsic and Extrinsic Factors on Skin Microbiota. Microorganisms 2021, 9, 543. [Google Scholar] [CrossRef]
- Fierer, N.; Hamady, M.; Lauber, C.L.; Knight, R. The Influence of Sex, Handedness, and Washing on the Diversity of Hand Surface Bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 17994–17999. [Google Scholar] [CrossRef]
- Ying, S.; Zeng, D.N.; Chi, L.; Tan, Y.; Galzote, C.; Cardona, C.; Lax, S.; Gilbert, J.; Quan, Z.X. The Influence of Age and Gender on Skin-Associated Microbial Communities in Urban and Rural Human Populations. PLoS ONE 2015, 10, e0141842. [Google Scholar] [CrossRef]
- Park, J.; Schwardt, N.H.; Jo, J.H.; Zhang, Z.; Pillai, V.; Phang, S.; Brady, S.M.; Portillo, J.A.; MacGibeny, M.A.; Liang, H.; et al. Shifts in the Skin Bacterial and Fungal Communities of Healthy Children Transitioning through Puberty. J. Investig. Dermatol. 2022, 142, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Holland, K.T.; Bojar, R.A. Cosmetics: What Is Their Influence on the Skin Microflora? Am. J. Clin. Dermatol. 2002, 3, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Babeluk, R.; Jutz, S.; Mertlitz, S.; Matiasek, J.; Klaus, C. Hand Hygiene—Evaluation of Three Disinfectant Hand Sanitizers in a Community Setting. PLoS ONE 2014, 9, e111969. [Google Scholar] [CrossRef]
- Burns, E.M.; Ahmed, H.; Isedeh, P.N.; Kohli, I.; Van Der Pol, W.; Shaheen, A.; Muzaffar, A.F.; Al-Sadek, C.; Foy, T.M.; Abdelgawwad, M.S.; et al. Ultraviolet Radiation, Both UVA and UVB, Influences the Composition of the Skin Microbiome. Exp. Dermatol. 2019, 28, 136–141. [Google Scholar] [CrossRef]
- Huang, S.; Haiminen, N.; Carrieri, A.P.; Hu, R.; Jiang, L.; Parida, L.; Kim, H.J.; Kim, J.J.; Myeong, N.R.; Kim, T.; et al. Human Skin, Oral, and Gut Microbiomes Predict Chronological Age. mSystems 2020, 5, e00630-19. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, J.J.; Myeong, N.R.; Kim, T.; Kim, D.; An, S.; Kim, H.; Park, T.; Jang, S.I.; Yeon, J.H.; et al. Segregation of Age-Related Skin Microbiome Characteristics by Functionality. Sci. Rep. 2019, 9, 16748. [Google Scholar] [CrossRef]
- Shibagaki, N.; Suda, W.; Clavaud, C.; Bastien, P.; Takayasu, L.; Iioka, E.; Kurokawa, R.; Yamashita, N.; Hattori, Y.; Shindo, C.; et al. Aging-Related Changes in the Diversity of Women’s Skin Microbiomes Associated with Oral Bacteria. Sci. Rep. 2017, 7, 10567. [Google Scholar] [CrossRef] [PubMed]
- Jugé, R.; Rouaud-Tinguely, P.; Breugnot, J.; Servaes, K.; Grimaldi, C.; Roth, M.P.; Coppin, H.; Closs, B. Shift in Skin Microbiota of Western European Women across Aging. J. Appl. Microbiol. 2018, 125, 907–916. [Google Scholar] [CrossRef]
- Howard, B.; Bascom, C.C.; Hu, P.; Binder, R.L.; Fadayel, G.; Huggins, T.G.; Jarrold, B.B.; Osborne, R.; Rocchetta, H.L.; Swift, D.; et al. Aging-Associated Changes in the Adult Human Skin Microbiome and the Host Factors That Affect Skin Microbiome Composition. J. Investig. Dermatol. 2022, 142, 1934–1946.e21. [Google Scholar] [CrossRef]
- Trinh, P.; Zaneveld, J.R.; Safranek, S.; Rabinowitz, P.M. One Health Relationships between Human, Animal, and Environmental Microbiomes: A Mini-Review. Front. Public Health 2018, 6, 235. [Google Scholar] [CrossRef]
- Hogan, P.G.; Mork, R.L.; Boyle, M.G.; Muenks, C.E.; Morelli, J.J.; Thompson, R.M.; Sullivan, M.L.; Gehlert, S.J.; Merlo, J.R.; McKenzie, M.G.; et al. Interplay of Personal, Pet, and Environmental Colonization in Households Affected by Community-Associated Methicillin-Resistant Staphylococcus aureus. J. Infect. 2019, 78, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Buhaș, M.C.; Candrea, R.; Gavrilaș, L.I.; Miere, D.; Tătaru, A.; Boca, A.; Cătinean, A. Transforming Psoriasis Care: Probiotics and Prebiotics as Novel Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 11225. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
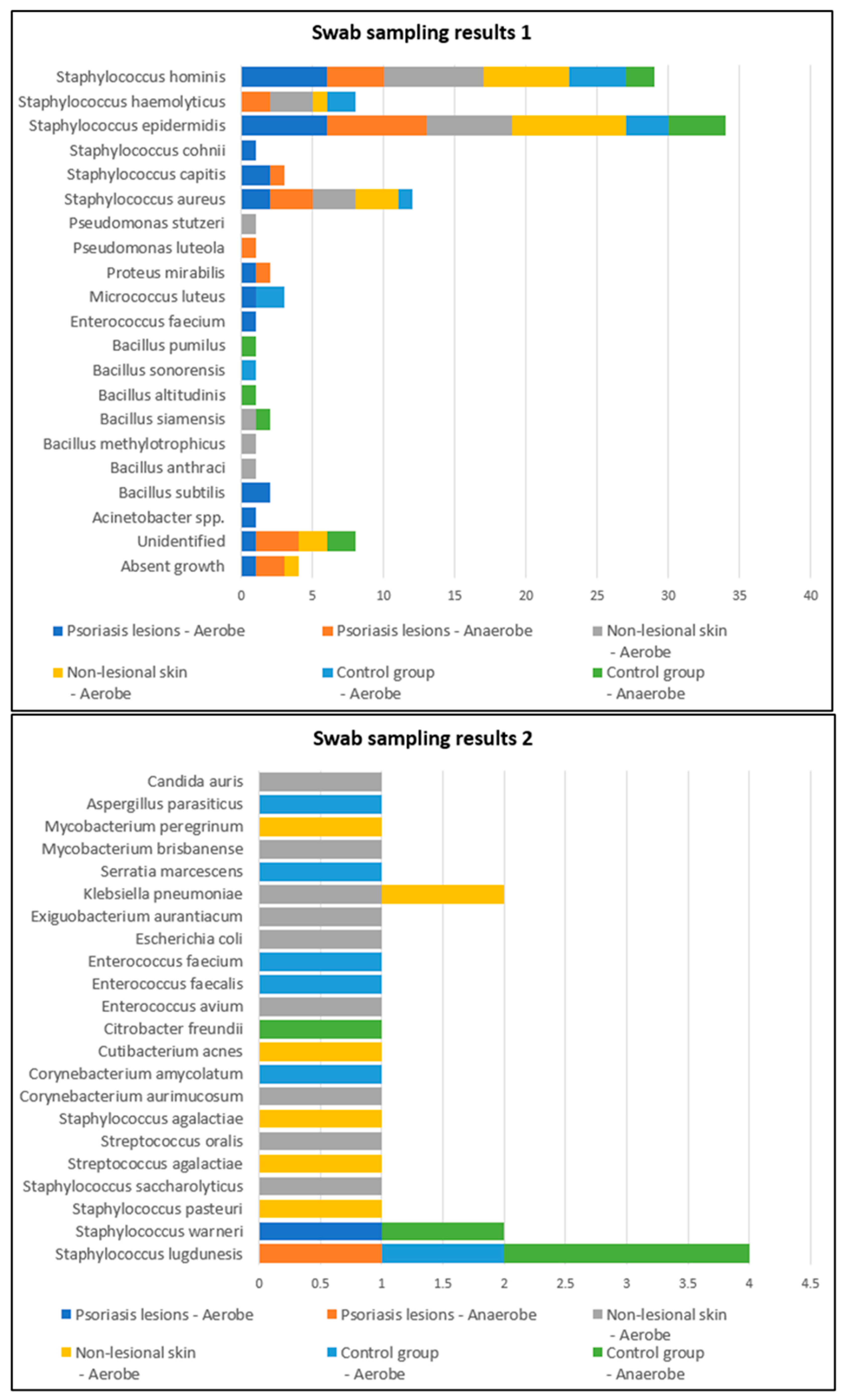
| Control | Lesional | Nonlesional | Pearson Chi2 Test | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Number of Strains | % | Number of Strains | % | Number of Strains | % | ||||
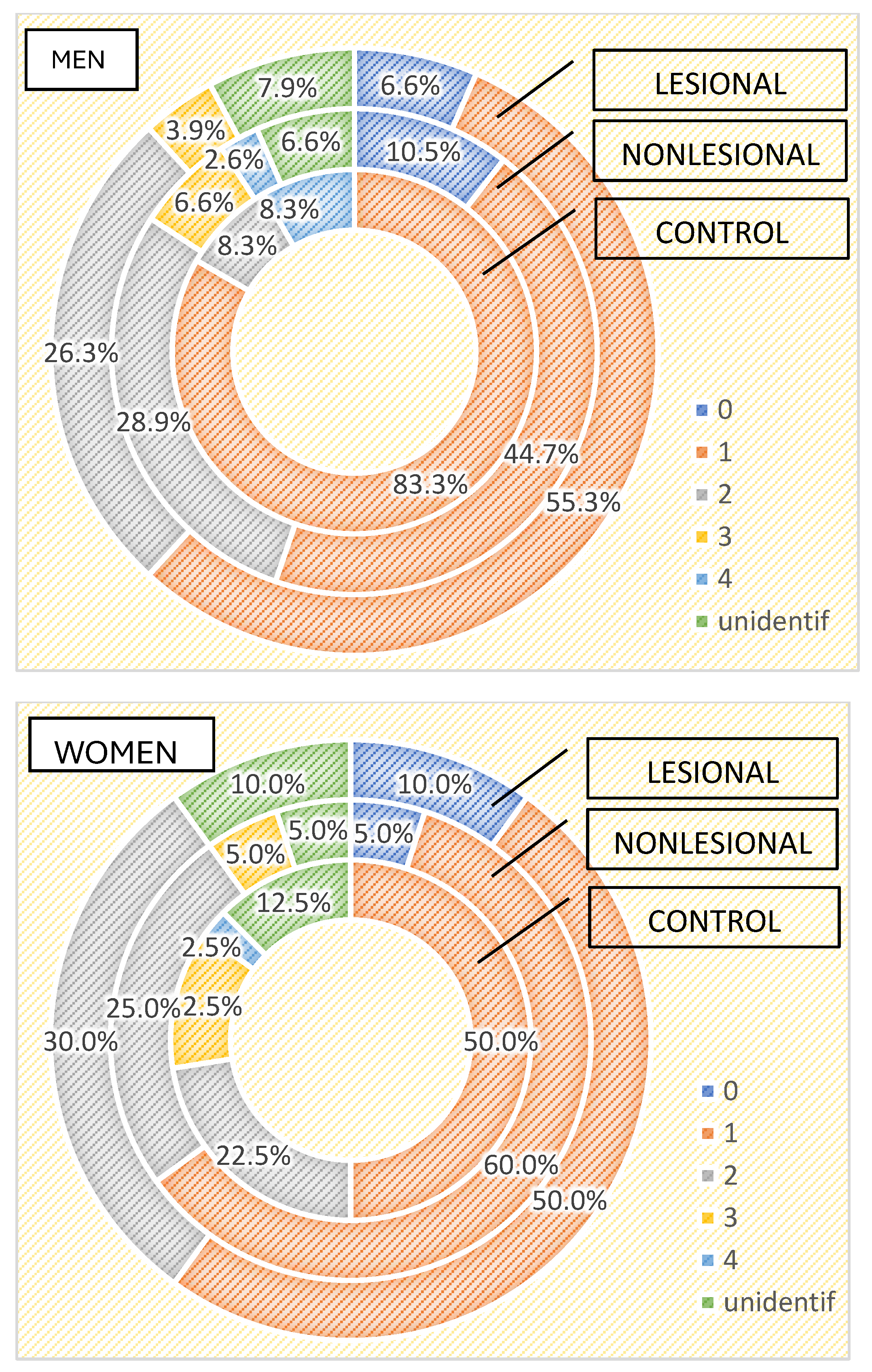
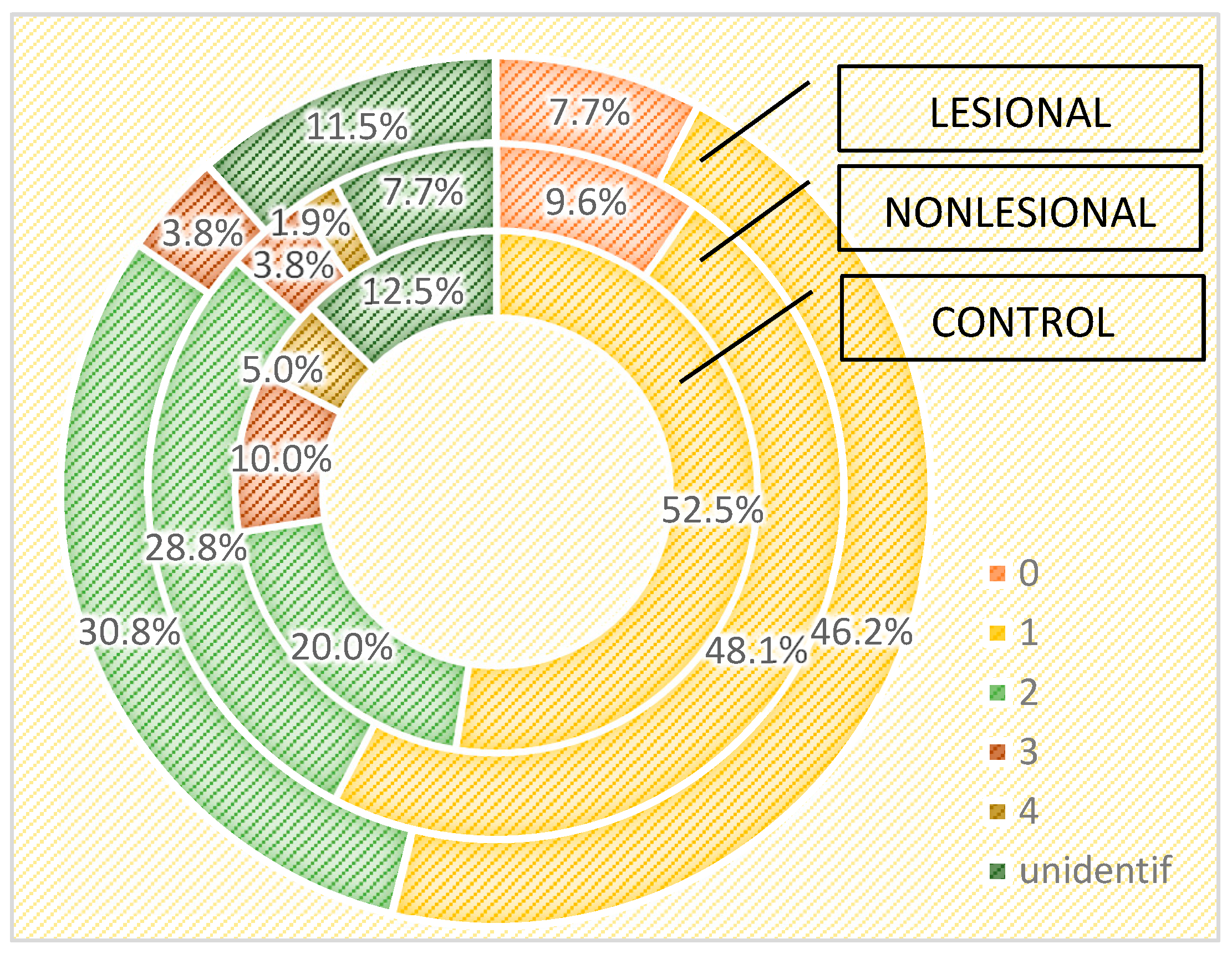
| TOTAL | Number of strains | 0 | 7 | 7.3% | 9 | 9.4% | Chi2 = 12.819 | ||
| 1 | 30 | 57.7% | 52 | 54.2% | 46 | 47.9% | p = 0.234 | ||
| 2 | 10 | 19.2% | 26 | 27.1% | 27 | 28.1% | |||
| 3 | 5 | 9.6% | 3 | 3.1% | 6 | 6.3% | |||
| 4 | 2 | 3.8% | 2 | 2.1% | |||||
| Unidentified | 5 | 9.6% | 8 | 8.3% | 6 | 6.3% | |||
| Total | 52 | 100.0% | 96 | 100.0% | 96 | 100.0% | |||
| Psoriasis Lesions | Nonlesional Skin of Psoriasis Patients | Controls | |
|---|---|---|---|
| Bacillus subtilis | 3 (3.1%) | 0 | 0 |
| Staphylococcus aureus | 13 (13.5%) | 12 (12.5%) | 1 (1.9%) |
| Staphylococcus epidermidis | 35 (36.5%) | 24 (25%) | 13 (25%) |
| Bacillus cereus | 4 (4.2%) | 3 (3.1%) | 8 (15.4%) |
| Klebsiela pneumoniae | 0 | 3 (3.1%) | 0 |
| Control | Lesional | Nonlesional | Pearson Chi 2 Test | |||||
|---|---|---|---|---|---|---|---|---|
| Number of Strains | % | Number of Strains | % | Number of Strains | % | |||
| MOIST REGION | Bacillus anthraci | - | ||||||
| Bacillus cereus | 1 | 7.1% | 1 | 7.1% | 1 | 2.3% | p = 0.602 | |
| Bacillus pumilus | 1 | 2.3% | p = 0.724 | |||||
| Bacillus siamensis | 1 | 2.3% | p = 0.724 | |||||
| Bacillus subtilis | - | |||||||
| Bacillus spp. | 2 | 4.5% | p = 0.520 | |||||
| Candida spp. | 1 | 2.3% | p = 0.724 | |||||
| Corynebacterium spp. | 1 | 7.1% | 5 | 11.4% | p = 0.401 | |||
| Enterococcus spp. | 1 | 7.1% | 2 | 4.5% | p = 0.627 | |||
| Escherichia spp. | - | |||||||
| Klebsiella pneumoniae | 2 | 4.5% | p = 0.520 | |||||
| Micrococcus luteus | 1 | 7.1% | p = 0.122 | |||||
| Proteus mirabilis | 1 | 2.3% | p = 0.724 | |||||
| Pseudomonas spp. | 1 | 2.3% | p = 0.724 | |||||
| Staphylococcus aureus | 4 | 28.6% | 6 | 13.6% | p = 0.091 | |||
| Staphylococcus capitis | 2 | 4.5% | p = 0.520 | |||||
| Staphylococcus epidermidis | 2 | 14.3% | 3 | 21.4% | 13 | 29.5% | p = 0.487 | |
| Staphylococcus haemolyticus | 1 | 7.1% | 5 | 11.4% | p = 0.401 | |||
| Staphylococcus hominis | 4 | 28.6% | 4 | 28.6% | 14 | 31.8% | p = 0.958 | |
| Staphylococcus lugdunensis | 2 | 14.3% | p = 0.014 | |||||
| Staphylococcus warneri | 1 | 7.1% | 1 | 2.3% | p = 0.489 | |||
| Staphylococcus spp. | 5 | 11.4% | p = 0.181 | |||||
| Streptococcus spp. | - | |||||||
| Others | 3 | 21.4% | 2 | 14.3% | 3 | 6.8% | p = 0.290 | |
| Total | 14 | 100.0% | 14 | 100.0% | 44 | 100.0% | ||
| Control | Lesional | Nonlesional | Pearson Chi2 Test | |||||
|---|---|---|---|---|---|---|---|---|
| Number of Strains | % | Number of Strains | % | Number of Strains | % | |||
| DRY REGION | Bacillus anthraci | 2 | 5.3% | 1 | 1.2% | 1 | 1.9% | p = 0.382 |
| Bacillus cereus | 7 | 18.4% | 3 | 3.7% | 2 | 3.8% | p = 0.007 | |
| Bacillus pumilus | 2 | 5.3% | 1 | 1.2% | 4 | 7.7% | p = 0.166 | |
| Bacillus siamensis | 2 | 5.3% | 1 | 1.2% | p = 0.150 | |||
| Bacillus subtilis | 3 | 3.7% | p = 0.187 | |||||
| Bacillus spp. | 5 | 13.2% | 2 | 2.4% | 3 | 5.8% | p = 0.066 | |
| Candida spp. | 2 | 5.3% | 2 | 2.4% | p = 0.261 | |||
| Corynebacterium spp. | - | |||||||
| Enterococcus spp. | 2 | 5.3% | 1 | 1.2% | 1 | 1.9% | p = 0.382 | |
| Escherichia spp. | 1 | 2.6% | 2 | 2.4% | 1 | 1.9% | p = 0.972 | |
| Klebsiella pneumoniae | 1 | 1.9% | p = 0.313 | |||||
| Micrococcus luteus | 1 | 2.6% | 2 | 2.4% | 1 | 1.9% | p = 0.972 | |
| Proteus mirabilis | 3 | 3.7% | p = 0.187 | |||||
| Pseudomonas spp. | 1 | 2.6% | 1 | 1.2% | p = 0.515 | |||
| Staphylococcus aureus | 1 | 2.6% | 9 | 11.0% | 6 | 11.5% | p = 0.275 | |
| Staphylococcus capitis | 2 | 5.3% | 3 | 3.7% | 3 | 5.8% | p = 0.835 | |
| Staphylococcus epidermidis | 11 | 28.9% | 32 | 39.0% | 11 | 21.2% | p = 0.088 | |
| Staphylococcus haemolyticus | 1 | 2.6% | 6 | 7.3% | 4 | 7.7% | p = 0.559 | |
| Staphylococcus hominis | 5 | 13.2% | 11 | 13.4% | 9 | 17.3% | p = 0.793 | |
| Staphylococcus lugdunensis | 1 | 2.6% | 4 | 4.9% | p = 0.260 | |||
| Staphylococcus warneri | 2 | 5.3% | 3 | 3.7% | 1 | 1.9% | p = 0.690 | |
| Staphylococcus spp. (altele) | 1 | 2.6% | 2 | 2.4% | 1 | 1.9% | p = 0.972 | |
| Streptococcus spp. | 1 | 2.6% | 1 | 1.2% | 2 | 3.8% | p = 0.611 | |
| Others | 5 | 13.2% | 5 | 6.1% | 6 | 11.5% | p = 0.372 | |
| Total | 38 | 100.0% | 82 | 100.0% | 52 | 100.0% | ||
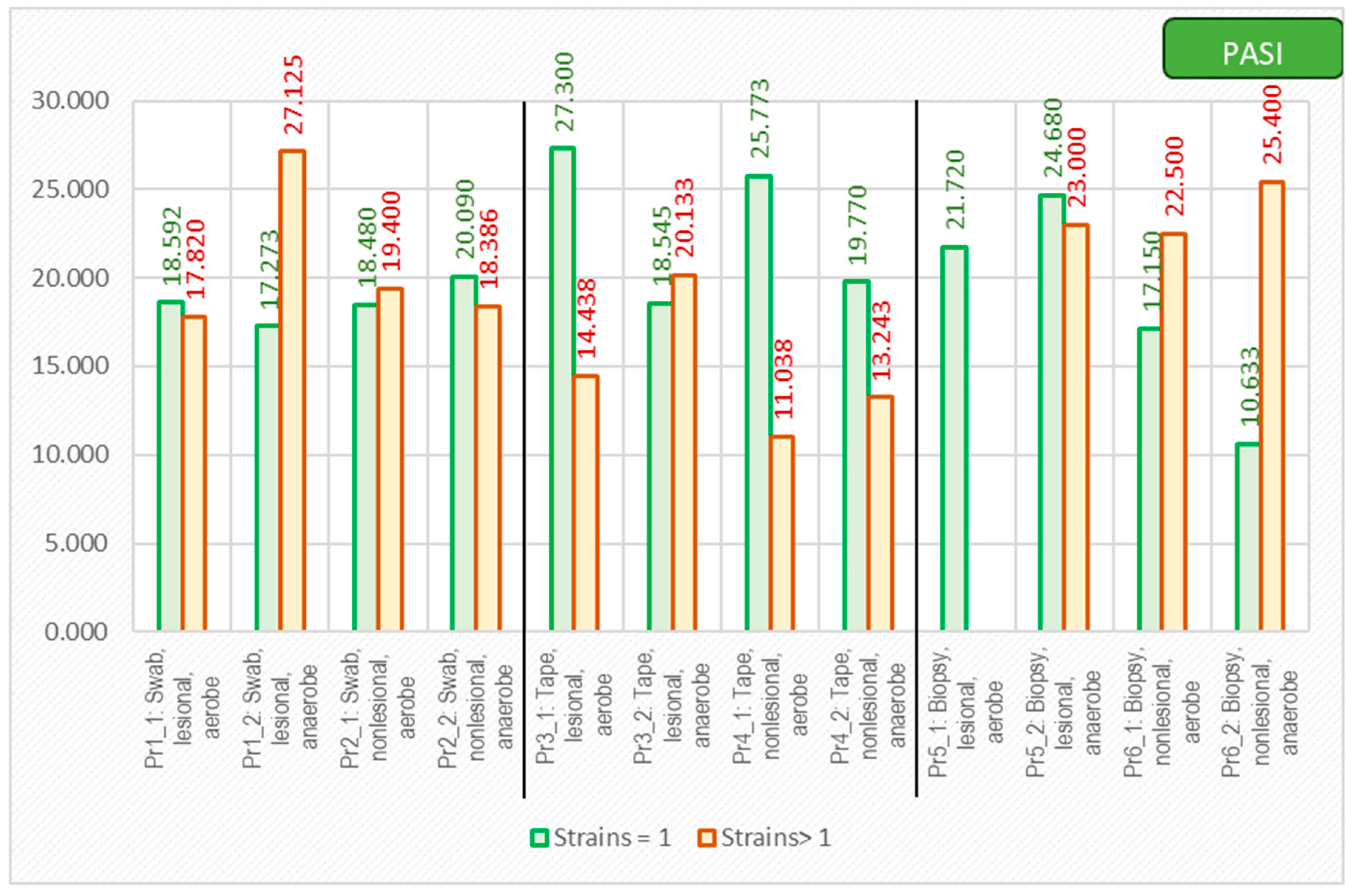
| Mean | Std. Error of Mean | Std. Deviation | Min | Max | Median | t-Student/Mann–Whitney Test | ||
|---|---|---|---|---|---|---|---|---|
| PASI | Total | 18.940 | 2.9008 | 12.9729 | 2.0 | 43.9 | 14.700 | |
| Male | 20.407 | 3.2877 | 12.7331 | 4.1 | 43.9 | 15.000 | t = 0.870 | |
| Female | 14.540 | 6.3195 | 14.1307 | 2.0 | 35.1 | 10.000 | p = 0.396 | |
| Urban | 15.609 | 3.5188 | 11.6706 | 2.0 | 35.1 | 14.400 | t = −1.292 | |
| Rural | 23.011 | 4.6613 | 13.9839 | 7.0 | 43.9 | 25.400 | p = 0.213 |
| Mean | Std. Error of Mean | Std. Deviation | Min | Max | Median | t-Student/Mann–Whitney test | ||
|---|---|---|---|---|---|---|---|---|
| NAPSI | Total | 9.45 | 2.775 | 12.412 | 0 | 40 | 4.00 | |
| Male | 9.80 | 3.538 | 13.702 | 0 | 40 | 0.00 | U = 35.00 | |
| Female | 8.40 | 3.816 | 8.532 | 0 | 20 | 10.00 | p = 0.852 | |
| Urban | 7.27 | 2.293 | 7.604 | 0 | 20 | 8.00 | U = 47.000 | |
| Rural | 12.11 | 5.569 | 16.707 | 0 | 40 | 0.00 | p = 0.839 |
| Mean | Std. Error of Mean | Std. Deviation | Min | Max | Median | t-Student/Mann–Whitney test | ||
|---|---|---|---|---|---|---|---|---|
| DLQI | Total | 15.50 | 1.713 | 7.661 | 2 | 28 | 15.00 | |
| Male | 14.20 | 1.979 | 7.664 | 2 | 28 | 15.00 | t = −1.342 | |
| Female | 19.40 | 3.092 | 6.914 | 10 | 25 | 24.00 | p = 0.196 | |
| Urban | 14.09 | 2.425 | 8.043 | 2 | 25 | 14.00 | t = −0.905 | |
| Rural | 17.22 | 2.414 | 7.242 | 5 | 28 | 15.00 | p = 0.377 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radaschin, D.S.; Iancu, A.V.; Ionescu, A.M.; Gurau, G.; Niculet, E.; Bujoreanu, F.C.; Beiu, C.; Tatu, A.L.; Popa, L.G. Comparative Analysis of the Cutaneous Microbiome in Psoriasis Patients and Healthy Individuals—Insights into Microbial Dysbiosis: Final Results. Int. J. Mol. Sci. 2024, 25, 10583. https://doi.org/10.3390/ijms251910583
Radaschin DS, Iancu AV, Ionescu AM, Gurau G, Niculet E, Bujoreanu FC, Beiu C, Tatu AL, Popa LG. Comparative Analysis of the Cutaneous Microbiome in Psoriasis Patients and Healthy Individuals—Insights into Microbial Dysbiosis: Final Results. International Journal of Molecular Sciences. 2024; 25(19):10583. https://doi.org/10.3390/ijms251910583
Chicago/Turabian StyleRadaschin, Diana Sabina, Alina Viorica Iancu, Alexandra Mariana Ionescu, Gabriela Gurau, Elena Niculet, Florin Ciprian Bujoreanu, Cristina Beiu, Alin Laurentiu Tatu, and Liliana Gabriela Popa. 2024. "Comparative Analysis of the Cutaneous Microbiome in Psoriasis Patients and Healthy Individuals—Insights into Microbial Dysbiosis: Final Results" International Journal of Molecular Sciences 25, no. 19: 10583. https://doi.org/10.3390/ijms251910583
APA StyleRadaschin, D. S., Iancu, A. V., Ionescu, A. M., Gurau, G., Niculet, E., Bujoreanu, F. C., Beiu, C., Tatu, A. L., & Popa, L. G. (2024). Comparative Analysis of the Cutaneous Microbiome in Psoriasis Patients and Healthy Individuals—Insights into Microbial Dysbiosis: Final Results. International Journal of Molecular Sciences, 25(19), 10583. https://doi.org/10.3390/ijms251910583