Abstract
Insecticide resistance in insects, driven by the overexpression of P450 enzymes, presents a significant challenge due to the enhanced metabolic detoxification of insecticides. Although the transcriptional regulation of P450 genes is not yet fully understood, G-protein-coupled receptor (GPCR) genes have emerged as key regulators in this process. This study is the first to associate GPCR genes with insecticide resistance in Musca domestica. We identified two key rhodopsin-like GPCR genes, ALHF_02706.g1581 and ALHF_04422.g2918, which were significantly overexpressed in the resistant ALHF strain compared to sensitive strains. Notably, both ALHF_02706.g1581 and ALHF_04422.g2918 were mapped to autosome 2, where critical but unidentified regulatory factors controlling resistance and P450 gene regulation are located. This supports our hypothesis that GPCRs function as trans-regulatory factors for P450-mediated resistance. Functional analysis using transgenic Drosophila demonstrated that overexpression of these rhodopsin-like GPCR genes increased permethrin resistance by approximately two-fold. Specifically, ALHF_02706.g1581 overexpression significantly upregulated the Drosophila resistance-related P450 genes CYP12D1, CYP6A2, and CYP6A8, while ALHF_04422.g2918 increased CYP6G1 and CYP6A2 expression, thereby enhancing insecticide detoxification in rhodopsin-like GPCR transgenic Drosophila lines. These findings suggest that these rhodopsin-like GPCR genes on autosome 2 may act as trans-regulatory factors for P450-mediated resistance, underscoring their critical role in insecticide detoxification and resistance development in M. domestica.
1. Introduction
The house fly, Musca domestica, is a significant domestic, medical, and veterinary pest that serves as a vector for over 200 human and animal pathogens [1,2,3]. Efforts to manage house fly populations have primarily relied on the application of insecticides [4]. However, the development of resistance to these insecticides has posed a major challenge to effective fly management [5,6,7]. Due to its rapid development of resistance and cross-resistance, well-established linkage maps for five autosomes and two sex chromosomes (X and Y), relatively well-understood biochemistry and genetics of insecticide resistance, and the availability of transcriptome and genome sequences [8,9,10], the house fly has been a valuable model for studying resistance mechanisms.
Insect P450s play a critical role in the detoxification of insecticides. Building on our previous transcriptome and gene functional studies in house flies [8,11], we identified several overexpressed P450s as the major functional group involved in house fly resistance. The overexpression of P450s results in elevated levels of P450 proteins and activities, leading to enhanced metabolic detoxification of insecticides. This process is recognized as one of the most important mechanisms of insecticide resistance in insects [7,12,13]. Numerous studies have shown that the overexpression of P450 genes is regulated by trans- or cis-regulatory factors [8,11,14,15,16,17,18,19,20,21,22]. Identifying and characterizing these regulatory factors is crucial for advancing our understanding of the development of insecticide resistance in insects.
G-protein-coupled receptors (GPCRs) interact with G proteins, which subsequently activate downstream regulatory factors such as the G-protein alpha subunit (Gαs), adenylate cyclase (AC), cyclic adenosine monophosphate (cAMP), and protein kinase A (PKA), thereby regulating various physiological processes [23,24,25]. In insects, GPCRs have been reported to regulate growth and development [26], and they have been implicated in stress responses to desiccation [27] and blood-feeding behavior [28]. However, the downstream factors in these GPCR pathways are not yet fully understood. The role of GPCRs in insecticide resistance remains largely unexplored, and no insecticides have been specifically developed to target GPCRs. [29,30,31,32] reported the overexpression of a GPCR gene in a highly pyrethroid-resistant mosquito Culex quiquenfaciatus, and demonstrated the involvement of this GPCR and its downstream regulatory factors (Gαs, AC, cAMP, and PKA) in regulating the expression of resistance-related P450 genes, ultimately leading to P450-mediated insecticide resistance in resistant mosquitoes [29,30,33,34,35]. Employing whole transcriptome analyses [8,11] identified multiple P450 genes and GPCR genes that were overexpressed in pyrethroid-resistant house flies. This finding led to the proposal that these overexpressed GPCR genes might play a role in regulating P450-mediated resistance in house flies.
Building on our earlier transcriptome data analysis, which involved multiple detoxification genes—primarily P450s—and signaling regulatory genes of GPCRs in resistant house flies [8], our characterization of the role of P450 overexpression in resistance [11], and our discovery of cis/trans factors in the co-regulation of resistance-related P450 gene expression in resistant house flies [11], the current study aimed to explore the potential regulatory interaction between GPCR overexpression and P450 gene expression, which plays a crucial role in mediating insecticide resistance in house flies. We assessed the expression of two rhodopsin-like GPCR genes in both resistant and susceptible house flies and conducted in vivo functional studies of these GPCRs in P450-mediated resistance using transgenic Drosophila melanogaster lines. We further examined the relative expression of four resistance-related P450 genes in the GPCR transgenic Drosophila lines to pinpoint the role of these rhodopsin-like GPCRs in the regulation of resistance-related P450s. Furthermore, we conducted autosomal mapping of the overexpressed rhodopsin-like GPCR genes on Muller elements [36] to investigate the linkage of these rhodopsin-like GPCRs to trans-regulatory factors on autosome 2, which has been shown to play a role in the regulation of P450 gene expression in resistant house flies. We hypothesized that GPCRs function as trans-regulatory factors for P450-mediated resistance.
2. Results
2.1. Rhodopsin-like GPCR Gene Up-Regulation in Permethrin Resistant House Fly, M. domestica
In the current study, we built upon previous transcriptome analysis findings [8] that identified multiple genes, including metabolic detoxification P450 genes and signaling transduction GPCR genes, as overexpressed in resistant house flies. Additionally, linkage analysis revealed that these overexpressed resistance-related P450s are regulated by trans-regulatory factors on autosome 2 [11]. To further explore these findings, we examined the expression levels of the identified GPCR genes, ALHF_02706.g1581 and ALHF_04422.g2918, in house fly strains using quantitative real-time polymerase chain reaction (qRT-PCR) with specific primer pairs for each gene (Table 1).

Table 1.
The list of primers used in the present study.
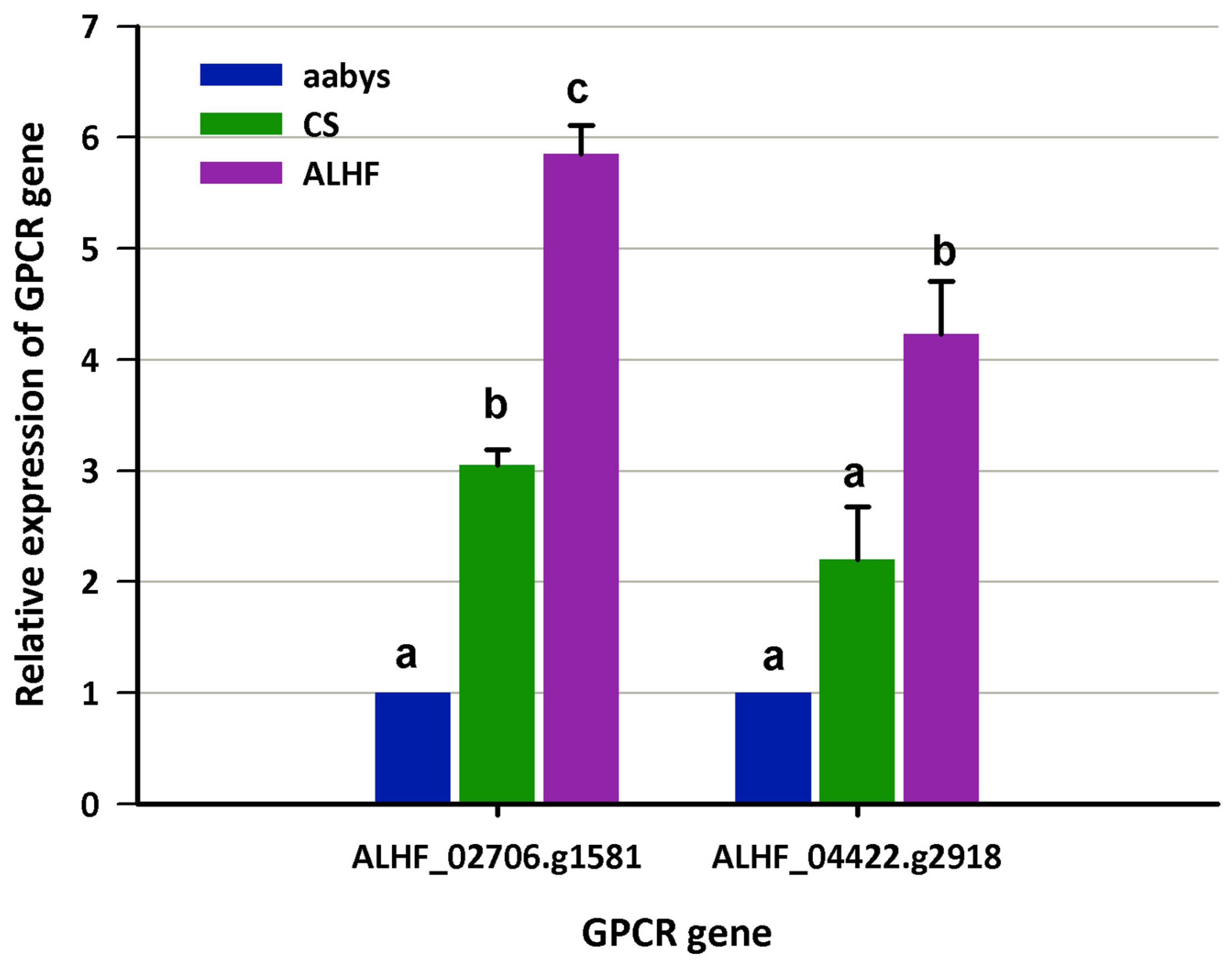
We compared the expression levels of these GPCR genes between the resistant ALHF strain and the susceptible aabys and CS strains. This approach was chosen to avoid false positive results that could arise from analyzing mosquitoes of different genetic backgrounds or inherent tolerances, rather than true resistance. qRT-PCR results confirmed that two GPCRs, ALHF_02706.g1581 and ALHF_04422.g2918, were significantly overexpressed in the ALHF-resistant strain compared to the two susceptible strains (Figure 1). Specifically, the relative expression levels of ALHF_02706.g1581 and ALHF_04422.g2918 were upregulated 5.9-fold and 4.2-fold, respectively, in ALHF compared to aabys, and 1.9-fold compared to the CS strain (Figure 1). These two GPCRs were the focus of further characterization to understand their roles in insecticide resistance in house flies.
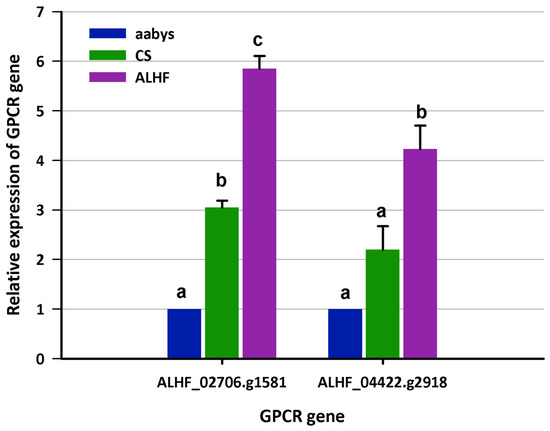
Figure 1.
The expression of GPCR genes in house flies, Musca domestica. The expression levels of GPCR genes ALHF_02706.g1581 and ALHF_04422.g2918 were assessed in the susceptible house fly strains aabys and CS, as well as in a resistant strain. mRNA levels of both GPCR genes were quantitatively measured using qRT-PCR. Results are presented as mean ± S.E. (n ≥ 3). Statistical significance in gene expression among samples, indicated by different letters (a, b, or c), was determined using one-way ANOVA with a p-value ≤ 0.05.
2.2. Localization and Mapping of Overexpressed GPCR Genes to the Autosomes of House Flies
The landmarks of Muller elements [37], known as the six highly conserved chromosomal arms of the well-annotated Drosophila genome [38], have enabled researchers to map genes from other species to autosomes [39]. Muller elements A–E are autosomal, while F is linked to the X chromosome. Meisel and Scott [36] have accurately mapped M. domestica genes to Muller elements, with Muller element A corresponding to house fly chromosome 3, B to chromosome 1, C to chromosome 5, D to chromosome 4, E to chromosome 2, and F to the X chromosome [36]. Using a similar methodology, we performed BLAST analysis [40] of two overexpressed GPCR genes, ALHF_02706.g1581 and ALHF_04422.g2918, against D. melanogaster sequences. The results showed that ALHF_02706.g1581 and ALHF_04422.g2918 aligned with the Drosophila homologs Dm Rh1 and Dm Rh2, respectively. Therefore, we refer to these two GPCR genes as rhodopsin-like GPCRs. These rhodopsin-like GPCR genes are located on chromosome 3R (Muller element E), which corresponds to autosome 2 of house flies [36] (Table 2).
Table 2.
Autosome assignments of overexpressed GPCR genes in house flies.
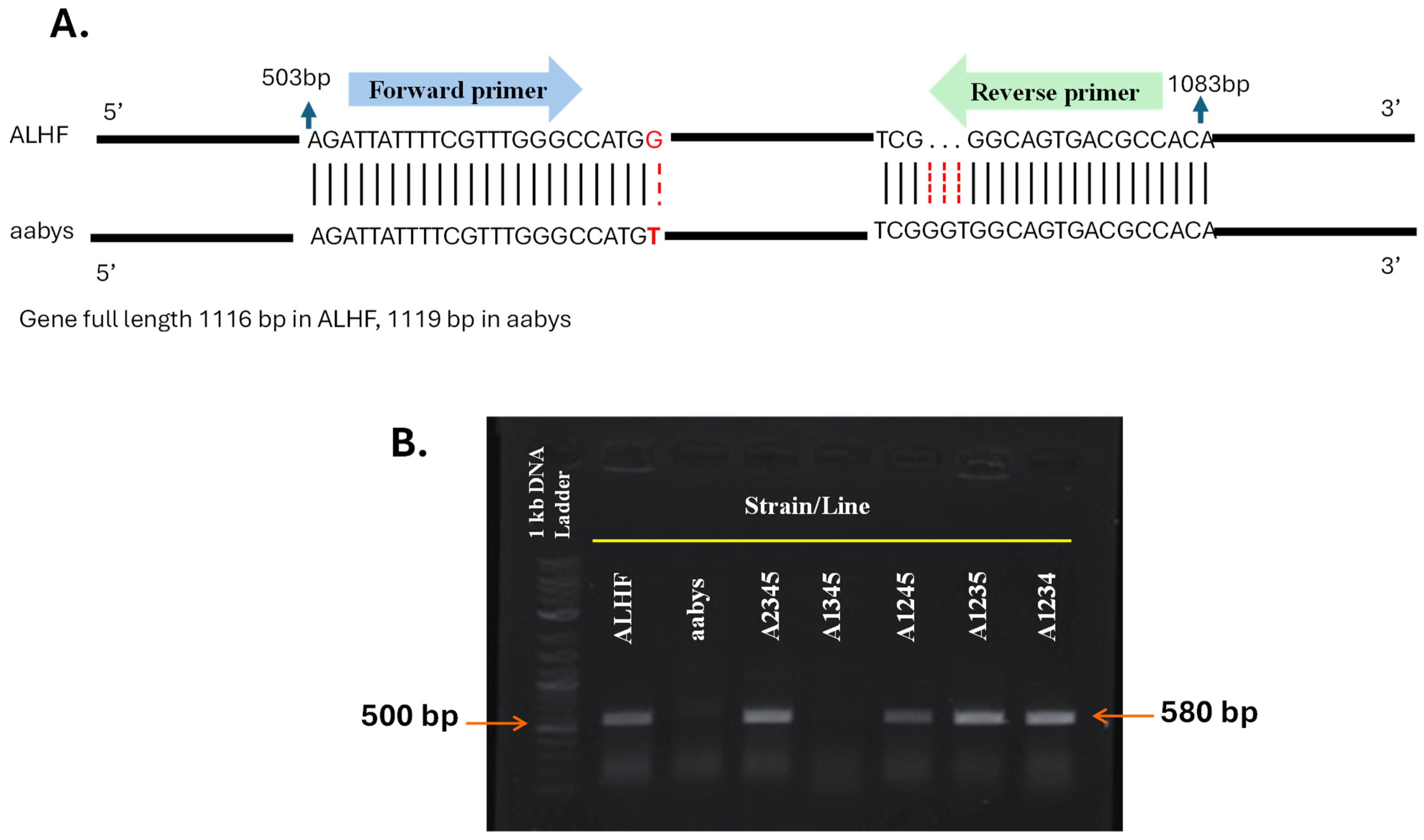
Validation of the autosomal location of the rhodopsin-like GPCR gene was further conducted using allele-specific PCR [19]. The rhodopsin-like GPCR gene ALHF_02706.g1581 was chosen for validation due to the presence of single nucleotide polymorphisms (SNPs) between the resistant ALHF strain and the aabys strain. In contrast, the ALHF_04422.g2918 gene showed identical sequences in both strains. The allele-specific PCR primers for the rhodopsin-like GPCR gene ALHF_02706.g1581 were designed based on the ALHF sequence, incorporating a specific SNP at the 3′ end of one primer to enable preferential amplification of ALHF alleles (Table 2, Figure 2A) [11,41]. Our results indicate that the ALHF allele-specific primer sets for ALHF_02706.g1581 successfully amplified specific DNA fragments (~580 bp) exclusively in flies carrying the wild-type autosome 2 markers from ALHF. No amplification occurred in flies homozygous for autosome 2 from either the aabys strain or the A1345 strain (Figure 2B).
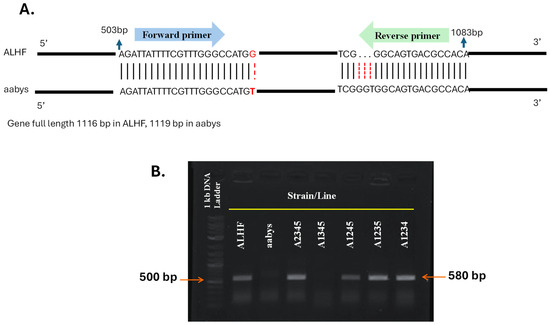
Figure 2.
Allele-specific RT-PCR for autosomal mapping of GPCR gene ALHF_02706.g1581 in M. domestica. The full length of the GPCR gene ALHF_02706.g1581 is 1116 bp in both the ALHF and aabys strains. (A), PCR fragments were generated using an allele-specific primer set designed according to the ALHF sequence of ALHF_02706.g1581. (B). The absence of a PCR product in a house fly line indicates that the gene is located on the corresponding autosome from aabys (e.g., the absence of a band in the A1345 line indicates that the gene is located on autosome 2).
PCR product-sequence analysis indicated that the amplified fragment corresponds to the house fly gene ALHF_02706.g1581. These findings confirm the mapping of the rhodopsin-like GPCR gene to autosome 2.
2.3. Up-Regulated House Fly Rhodopsin-like GPCR Genes in Transgenic D. melanogaster—An Approach for Defining the House Fly Rhodopsin-like GPCR Gene Function
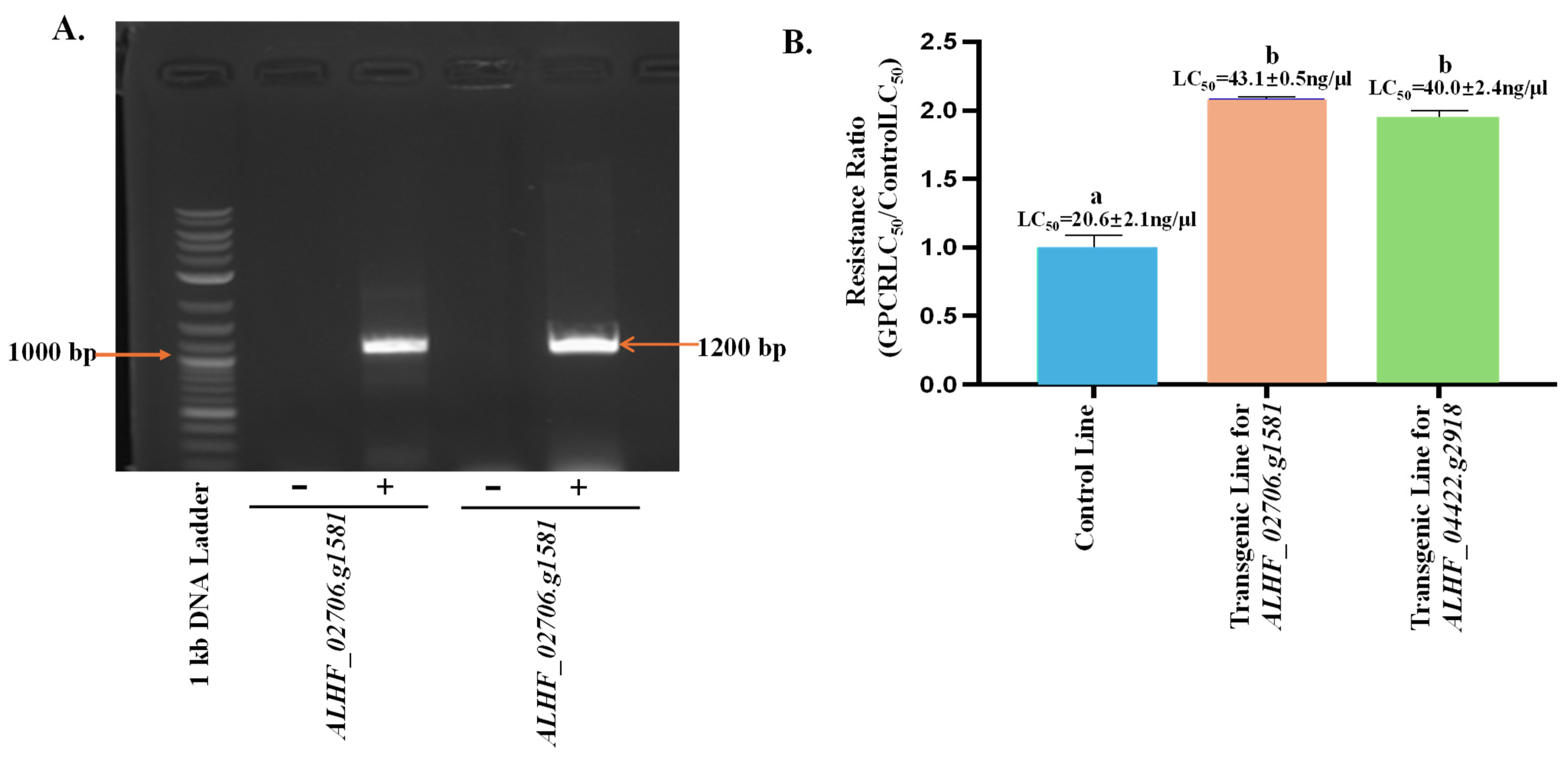
To characterize the function of the up-regulated genes ALHF_02706.g1581 and ALHF_04422.g2918 in the development of insecticide resistance in ALHF house flies, we conducted a transgenic study using the GAL4-UAS enhancer trap system in D. melanogaster. To confirm the presence of house fly rhodopsin-like GPCR genes in the transgenic Drosophila lines, we performed RT-PCR using full-length primer pairs for ALHF_02706.g1581 and ALHF_04422.g2918 (Table 1), and cDNA from three Drosophila lines: the control (F1 progeny of Bloomington stock #24484 D. melanogaster line containing an empty pUAST vector crossed with GAL4), transgenic ALHF_02706.g1581 (F1 progeny of homozygous transgenic ALHF_02706.g1581 crossed with GAL4), and transgenic ALHF_04422.g2918 (F1 progeny of homozygous transgenic ALHF_04422.g2918 crossed with GAL4). The results showed that the full lengths of ALHF_02706.g1581 and ALHF_04422.g2918 were only amplified in the ALHF_02706.g1581 and ALHF_04422.g2918 transgenic lines, respectively, with specific gene sizes ~1200 bp (Figure 3A), corresponding to the size of ~1200 bp for these two rhodopsin-like GPCR genes. Sequencing of the PCR products from these transgenic lines confirmed that they are indeed the full lengths of rhodopsin-like GPCR ALHF_02706.g1581 and ALHF_04422.g2918.
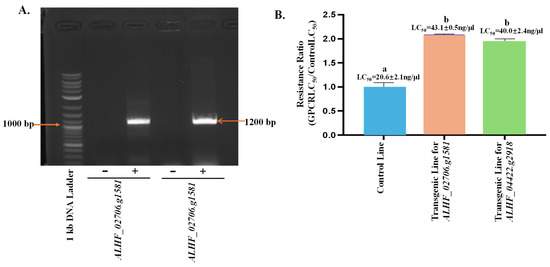
Figure 3.
Effects of overexpressed GPCR genes on permethrin resistance and P450 gene expression in Drosophila melanogaster. (A) RT-PCR amplification of the GPCR genes ALHF_02706.g1581 and ALHF_04422.g2918 in transgenic Drosophila melanogaster lines. “−” indicates the non-transgenic control D. melanogaster line, while “+” represents the transgenic D. melanogaster lines containing house fly GPCR genes. The 1 kb DNA Plus ladder (Biolabs) was used as the molecular size reference, with numbers indicating DNA band sizes in bp. (B) Toxicity of permethrin to non-transgenic (control) and transgenic Drosophila melanogaster lines. Resistance ratios were calculated as LD50 of D. melanogaster lines/LD50 of the control line. Statistical significance in gene expression among samples, indicated by different letters (a, b, or c), was determined using one-way ANOVA with a p-value ≤ 0.05. No significant differences in expression levels were observed between groups labeled with the same letter (p < 0.05).
The sensitivity of the D. melanogaster transgenic lines to permethrin was assessed through bioassays conducted on three Drosophila lines: the control (CK), ALHF_02706.g1581, and ALHF_04422.g2918. The transgenic lines ALHF_02706.g1581 and ALHF_04422.g2918 exhibited 2.1-fold and 2.0-fold increases, respectively in permethrin tolerance, compared to the control line, as measured by the LC50 values (Figure 3B, p = 0.005). These results suggest that the up-regulated rhodopsin-like GPCR genes in the house fly enhance permethrin resistance in D. melanogaster, underscoring the significant role of these genes in permethrin resistance in M. domestica.
2.4. Relative Expression Levels of Drosophila P450 Genes in House Fly Rhodopsin-like GPCR Transgenic Lines
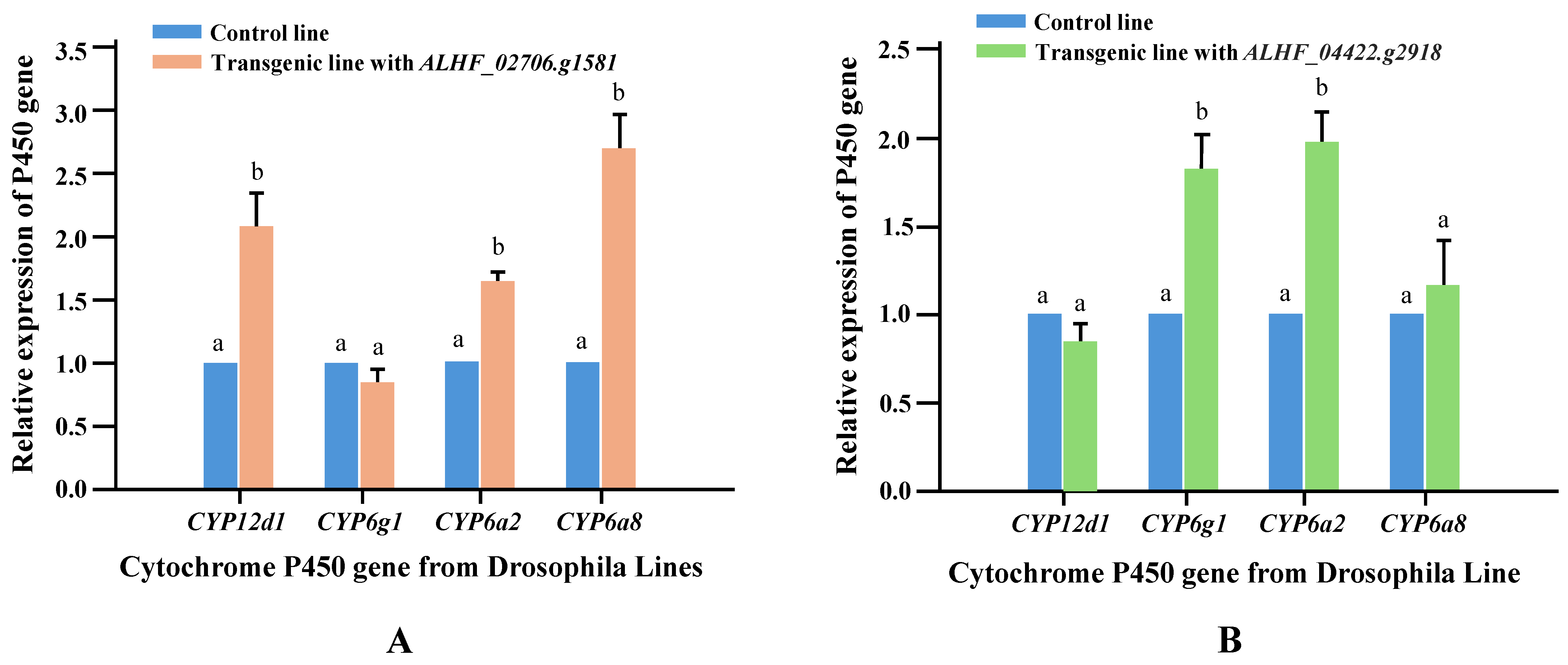
Li et al. [29] identified that resistance-related GPCRs can induce the overexpression of Drosophila P450s, which are known to be involved in insecticide resistance, and proposed that the molecular mechanism by which GPCRs contribute to insecticide resistance in transgenic Drosophila involves the regulation of these resistance P450 genes. To fully understand the role of GPCRs in permethrin resistance in transgenic Drosophila, we conducted qRT-PCR to measure the relative expression levels of four Drosophila resistance P450 genes (CYP12D1, CYP6G1, CYP6A2, and CYP6A8), which are associated with insecticide resistance in Drosophila [42]. The qRT-PCR was performed using cDNA from the control (CK), ALHF_02706.g1581, and ALHF_04422.g2918 transgenic Drosophila lines with primer pairs designed according to the sequences of CYP6A2, CYP6A8, CYP12D1, and CYP6G1 (Table 1). Significant increases (p < 0.001) in the expression of CYP12D1, CYP6A2, and CYP6A8 (2.1-, 1.7-, and 2.7-fold, respectively) were observed in the ALHF_02706.g1581 transgenic line compared to the control line (Figure 4A). In the ALHF_04422.g2918 transgenic line, increased expression levels were found for CYP6G1 (1.8-fold) and CYP6A2 (2.0-fold) (Figure 4B).
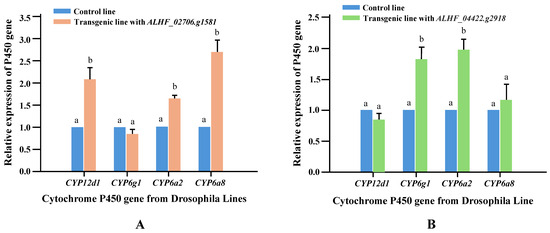
Figure 4.
Relative expression levels of four Drosophila resistance-related P450 genes in control and GPCR transgenic lines. The expression patterns of four P450 genes (CYP6A2, CYP6A8, CYP12D1, CYP6G1) were analyzed in control and transgenic Drosophila lines using qRT-PCR. (A) Expressing of the GPCR genes ALHF_02706.g1581 and (B) Expressing of the GPCR ALHF_04422.g2918. The relative expression levels in transgenic flies are shown relative to their expression in the control line. Results are presented as mean ± S.E. (n ≥ 3). Statistical significance in gene expression among samples, indicated by different letters (a or b), was determined using one-way ANOVA with p ≤ 0.05.
3. Discussion
The overexpression of P450 enzymes leads to elevated protein levels and enhanced metabolic detoxification of insecticides, establishing P450 detoxification as a key mechanism in insecticide resistance among insects. While our understanding of the regulation of insect P450 gene expression remains incomplete, studies have revealed the complex nature of these transcriptional regulation mechanisms [14,20,21,22,34,35,43,44]. Known regulatory factors include orphan nuclear receptors, Cap ‘n’ Collar C (CncC)/muscle aponeurosis fibromatosis (Maf), GPCRs, and cAMP-response element binding proteins, identified across various insect species [14,29,35,44]. For instance, Hu et al. [45] reported that the upregulation of CncC/Maf transcription factors contributes to the increased expression of CYP321A8 in a resistant strain of Spodoptera exigua, facilitated by specific promoter site mutations. Similarly, Yang et al. [44] demonstrated that the overexpression of CYP6CM1 in whiteflies, which confers resistance to neonicotinoid insecticides, is regulated by CREM, activated via the MAPK pathway. These findings underscore the intricate mechanisms involved in the transcriptional regulation of insect P450 genes, which are crucial for detoxifying insecticides and developing resistance.
GPCR genes and signaling pathways play a pivotal role in regulating P450 genes associated with resistance in several insect species, including Lymantria dispar [46], Culex pipiens [47], Culex quinquefasciatus [29,30], and M. domestica [48]. GPCR pathways have emerged as significant regulators of P450 gene expression. The GPCR regulatory pathway involving Gαs, adenylate cyclase (AC), protein kinase A (PKA), and cyclic AMP (cAMP) in mosquitoes has been systematically studied and modeled [29,30,34,35], showing its involvement in regulating multiple resistance-related P450 gene expressions. This pathway contributes to enhanced insecticide detoxification in Culex quinquefasciatus. Functional studies by Li et al. [29,30,35], using transgenic Drosophila, demonstrated that rhodopsin-like GPCRs increase permethrin tolerance and elevate the expression of resistance P450 genes CYP12D1 and CYP6A8 of Drosophila melanogaster. Additional studies using transgenic Drosophila lines further confirm the involvement of GPCRs in regulating P450 gene expression and insecticide resistance in L. dispar and M. domestica.
Our results demonstrate that the two rhodopsin-like GPCR genes, ALHF_02706.g1581 and ALHF_04422.g2918, differentially regulate cytochrome P450 (CYP) genes, providing important insights into the molecular mechanisms underlying insecticide resistance. Specifically, ALHF_02706.g1581 primarily upregulated the resistance-related P450 genes CYP12D1, CYP6A2, and CYP6A8, while ALHF_04422.g2918 upregulated CYP6G1 and CYP6A2. The differential regulation of these CYP genes by distinct GPCRs indicates not only that resistance mechanisms in house flies are highly specialized, with specific GPCRs selectively enhancing P450 gene expression depending on the insecticide, but also that these GPCRs may be involved in distinct signaling pathways. This specificity underscores the complexity of GPCR-mediated regulatory networks and highlights the potential for targeting these pathways in future strategies to combat insecticide resistance. CYP6A2, CYP6A8, CYP6G1, and CYP12D1 are known to be involved in the detoxification of various insecticides, including organophosphates, pyrethroids, DDT, and neonicotinoids [42,49,50]. The upregulation of these P450 genes by ALHF_02706.g1581 and ALHF_04422.g2918 could suggest a role for the GPCR genes in enhancing resistance to these compounds by facilitating the breakdown of insecticides. Notably, the upregulation of CYP6G1 and CYP6A2 by ALHF_04422.g2918 is particularly significant, as CYP6G1 has been widely implicated in resistance to multiple insecticides, including DDT and neonicotinoids, due to its broad substrate specificity. The high expression of CYP6G1 in insecticide-resistant Drosophila and other insect species suggests that ALHF_04422.g2918 plays a key role in promoting resistance to a diverse range of insecticides.
Both trans- and cis-regulatory factors have been implicated in the overexpression of P450 genes in resistant insects. The overexpression of CYP6A1 and CYP6D1 in the house fly M. domestica, and CYP6A2 and CYP6A8 in the fruit fly D. melanogaster, is trans-regulated by factors on autosome 2 [15,18,19], although the precise regulatory factors remain undefined. Similar trans-regulation of P450-mediated resistance has been proposed in the mosquito Aedes aegypti [22] but the regulatory factors have yet to be identified. A recent systematic study at the whole transcriptome level on insecticide-resistant house flies aimed to develop a deeper understanding of how P450 gene upregulation, interaction, metabolism, and function contribute to the development of insecticide resistance [8,11]. Combining in vivo functional studies using the D. melanogaster transgenic system, in vitro functional metabolism studies, in silico homology modeling, and molecular docking methods, Li Ming et al. [11] demonstrated that the overexpression of P450 genes in house flies contributes to pyrethroid resistance through increased insecticide metabolism and multiple P450 interactions and regulation. Genetic linkage analysis further revealed that the overexpression of these P450 genes is regulated by trans- and/or cis-acting factors, particularly on autosome 2, consistent with findings in other insect strains or species.
Our current research is progressing towards highlighting the significant role of G-protein-coupled receptor (GPCR) gene overexpression in regulating P450-mediated insecticide resistance in house flies. Our findings of increased permethrin resistance in transgenic Drosophila expressing house fly rhodopsin-like GPCR genes, consistent with findings in Culex mosquitoes, further confirm the role of GPCRs in insecticide resistance. The increased expression of specific resistance-related Drosophila P450 genes (CYP12D1, CYP6A2, CYP6A8, CYP6G1) [42] induced by overexpressed rhodopsin-like GPCR genes in transgenic Drosophila underscores the role of rhodopsin-like GPCRs in regulating these P450 genes, thereby enhancing insecticide detoxification. Furthermore, genome mapping of these overexpressed rhodopsin-like GPCR genes on autosome 2 in house flies, confirmed through Muller element analysis and allele-specific PCR, has pinpointed their location corresponding to that of regulatory factors [11]. These findings support our hypothesis that overexpressed GPCRs act as trans-regulatory factors for P450-mediated insecticide resistance in house flies. Nevertheless, the precise regulatory role of GPCRs in house fly P450 gene expression, and the full spectrum of factors involved in GPCR pathways responsible, together with GPCR, for the development of resistance, remain to be fully elucidated.
4. Materials and Methods
4.1. House Flies
Three house fly strains—ALHF, aabys, and CS—were used in this study. ALHF was originally collected from a poultry farm in Alabama in 1998. This strain exhibited a high level of resistance after being selected with permethrin for six generations and has been annually selected with permethrin to maintain its resistance level (~1100-fold) [5,51]. The aabys strain is insecticide-susceptible and carries five recessive morphological markers: ali-curve (ac), aristapedia (ar), brown body (bwb), yellow eyes (ye), and snipped wings (snp), distributed on autosomes 1, 2, 3, 4, and 5, respectively. CS is a wild-type insecticide-susceptible strain that has been kept in laboratory breeding for over five decades, sharing the same morphological characteristics as ALHF. The aabys and CS strains were originally obtained from Dr. J. G. Scott (Cornell University).
Five backcross (BC1) lines were genetically isolated as described by Liu and Yue [5] and Tian et al. [51]. Briefly, approximately 400 ALHF virgin females were crossed with aabys males to generate the first filial generation (F1). Since crossing over rarely occurs in male flies [52], the F1 males (about 200 flies) were backcrossed with same similar amount of aabys virgin females. This ensured that the presence of a mutant phenotype indicated that the respective autosome with a mutant-type marker was derived from aabys. Five homozygous back-cross (BC1) lines were isolated with the following genotypes: ac/ac, +/a+, +/+, +/+, +/+ (A2345); +/a+, ar/ar, +/+, +/+, +/+ (A1345); +/a+, +/a+, bwb/bwb, +/+, +/+ (A1245); +/+, +/+, +/+, ye/ye, +/+ (A1235); and +/+, +/+, +/+, +/+, snp/snp (A1234). The name of each line indicates which of its autosomes has a wild-type marker from ALHF. For example, the A2345 strain has wild-type markers on autosomes 2, 3, 4, and 5 from ALHF and the mutant marker (ali-curve) on autosome 1 from aabys. All house flies were reared at 25 ± 2 °C under a photoperiod of 12:12 (L:D) h.
4.2. RNA Extraction and cDNA Preparation
Twenty three-day-old virgin female house flies from each of the three strains (ALHF, aabys, and CS) and five BC1 lines were collected for RNA extraction. Total RNA was isolated using the acidic guanidine thiocyanate-phenol-chloroform method [53]. Following extraction, DNA was removed from 5 μg of total RNA using the TURBO DNA-free kit (Ambion), according to the manufacturer’s instructions. cDNA synthesis was performed using the ProtoScript® II First Strand cDNA Synthesis Kit (New England Biolabs, Ipswich, MA, USA). The resulting cDNA was quantified using a Nanodrop spectrophotometer before proceeding to qRT-PCR and PCR analyses. Each experiment was independently replicated more than three times, involving separate insect sample collections, RNA extractions, and cDNA syntheses.
4.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
qRT-PCR was performed using the SYBR Green PCR Master Mix kit and an ABI 7500 Quantitative PCR System (Applied Biosystems, Foster City, CA, USA). Each qRT-PCR reaction (25 µL) contained 1x SYBR Green master mix, 1 µL of cDNA, and a GPCR gene-specific primer pair (Table 1) at a final concentration of 3–5 µM. The β-actin gene, an endogenous control [54], was used as it remains constant across different tissues and in both resistant and susceptible house flies [8,11,41,55]. A “no-template” reaction served as a negative control. All samples were run in triplicate. The reaction cycle included an initial melting step at 50 °C for 2 min, followed by 95 °C for 10 min, and then 40 cycles of 95 °C for 15 s and 60 °C for 1 min. The PCR reaction was assessed by a melting curve analysis using Dissociation Curves software. Relative expression levels for specific genes were calculated using the 2−ΔΔCT method with SDS RQ software v.2.3 [56]. Each experiment was repeated three times with different cDNA samples.
The statistical significance of gene expression was calculated using a Student’s t-test for all two-sample comparisons and a one-way analysis of variance (ANOVA) for multiple sample comparisons, utilizing the Statistical Package for the Social Sciences (SPSS) software with both least significant difference (LSD) and Tukey tests to analyze the significance of means. A p-value ≤ 0.05 was considered statistically significant. Significant up-regulation was determined using a cut-off value of a two-fold change in expression [57].
4.4. Autosome Assignment and Mapping of Overexpressed GPCR Genes in M. domestica
Drosophila genomes are organized into six chromosomal arms, known as Muller elements [37], where Elements A–E are autosomal, and Element F corresponds to the ancestral X chromosome of Brachycera [58,59]. Mapping genes to these chromosome arms in Drosophila [39] facilitates the assignment of homologous genes from other species to these Muller elements. We mapped our GPCR gene sequences, ALHF_02706.g1581 (NCBI accession number: XP_005182983.1) and ALHF_04422.g2918 (NCBI accession number: XP_005191160.1), to homologous genes in the Drosophila genome, thereby assigning them to specific Muller elements. We then aligned these Muller elements to the corresponding house fly autosomes according to [36].
Allele-specific PCR was conducted to further validate the linkage mapping of the GPCR in house flies. GPCR gene ALHF_02706.g1581 was selected because its sequence showed nucleotide polymorphisms (SNPs) within the gene between the resistant ALHF and the aabys strains, whereas GPCR gene ALHF_04422.g2918 displayed identical sequences between the two strains. In this experiment, two house fly strains (ALHF and aabys) and five BC1 lines (A2345, A1345, A1245, A1235, and A1234) were used. Each BC1 line was homozygous for the recessive mutant allele from aabys and heterozygous for the dominant wild-type alleles from ALHF. Allele-specific PCR was performed using a two-round PCR strategy as described by Liu and Scott [19]. Briefly, the first PCR reaction was conducted using allele-independent primer pairs (Table 1) to generate GPCR cDNA fragments. This was followed by a second PCR reaction containing 0.5 µL of the first-round PCR reaction solution and the allele-specific primer pair (Table 1), which was designed based on the specific sequence of the genes from ALHF, with a specific nucleotide polymorphism at the 3’ end of the primer to allow preferential amplification of the allele from ALHF. Three biological replicates were conducted with different mRNA samples, and the PCR products were sequenced.
4.5. Amplification of Full Length of GPCR Genes
To study the function of GPCR genes, the full-length sequences of genes ALHF_02706.g1581 and ALHF_04422.g2918 were generated using Platinum Taq DNA Polymerase High-Fidelity (Invitrogen, Carlsbad, CA, USA) with cDNA from ALHF as the template. The PCR primers for full-length gene amplification were designed based on sequences from previous transcriptome data of the ALHF house fly strain. The forward primers included protecting bases CCG and an EcoRI restriction enzyme site (GAATTC), while the reverse primers contained CTA and an XbaI sequence (TCTAGA) (Table 1). The PCR process consisted of an initial denaturation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, 57 °C for 45 s, and 72 °C for 2 min and 30 s, with a final extension at 72 °C for 5 min. PCR products were purified using the QIAquick Gel Extraction Kit (Qiagen, Valencia, CA, USA) before being cloned for sequencing using the TOPO TA Cloning Kit (Invitrogen, Carlsbad, CA) and the One Shot TOPO 10 Chemically Competent E. coli Kit (Invitrogen, Carlsbad, CA). Plasmids from the TA clones were extracted using the E.Z.N.A. Plasmid DNA Mini Kit 1 (OMEGA Bio-tek, Inc., orcross, GA, USA). Cloning and sequence analyses were repeated at least three times and three TA clones from each replication to ensure the accuracy of GPCR cDNAs.
4.6. Construction of Transgenic Drosophila Flies
The plasmid containing the full length of each GPCR from the TA clone was digested using restriction enzymes EcoRI/XbaI and sub-cloned into a pUASTattB vector [60,61] [a gift from Dr. Johnnes Bischof, University of Zurich]. Plasmid DNA of pUASTattB-2706 and pUASTattB-4422 was extracted using the Endo-Free Plasmid Purification Kit (Qiagen, Valencia, CA) and were transformed into the germline of D. melanogaster (Bloomington stock #24484, genotype M{vas-int.Dm}ZH-2A, M{3xP3-RFP.attP’}ZH-58A), resulting in site-specific integration on chromosome 2R (Rainbow Transgenic Flies Inc., Camarillo, CA, USA) [62]. The following cross-breeding process was conducted as described by Li [29]. After microinjecting the plasmids pUASTattB-2706 and pUASTattB-4422 into fruit fly embryos, transgenic flies exhibiting orange eyes and straight wings were crossed with a W1118 strain (white eyes) to produce heterozygous transgenic Drosophila carrying the target GPCR transgenes, characterized by orange eyes and straight wings. These heterozygous transgenic flies were then crossed with a balancer strain with white eyes and curly wings (Bloomington stock #6312; genotype: w [1118]/Dp(1;Y)y[+]; sna[Sco]/CyO, P{ry[+t7.2]=sevRas1.V12}FK1). Offspring with orange eyes and curly wings were self-crossed to generate homozygous transgenic flies. The transgenic lines used for all subsequent assays were the F1 progeny from crosses between homozygous GPCR transgenic virgin females and GAL4-expressing males (ubiquitous Act5C driver line). This resulted in the lines ALHF_02706.g1581+GAL4 and ALHF_04422.g2918+GAL4. “The control group consisted of the F1 progeny from a cross between the control line (Bloomington stock #24484, D. melanogaster containing an empty pUAST vector) and GAL4-expressing males.
4.7. Toxicity of Permethrin on the Transgenic D. melanogaster
Permethrin toxicity bioassays were performed on 2–3 day post-eclosion female homozygous transgenic Drosophila of ALHF_02706.g1581 and ALHF_04422.g2918, and CK to assess their susceptibility to permethrin. Serial concentrations of permethrin solution in acetone, ranging from 12 to 70 ng/μL, were prepared, ensuring mortality rates between 0% and 100% for the tested Drosophila. Each concentration (200 μL) was evenly applied to the inner surface of individual 20 mL glass scintillation vials. Fifteen female flies were then placed into each prepared vial. The vials were sealed with cotton balls soaked in 5% sucrose. Control group vials were coated with acetone alone and plugged with identical 5% sucrose-soaked cotton balls. Mortality was assessed after 24 h of exposure to permethrin. Susceptibility to permethrin in transgenic flies was determined by comparing the LC50 values of GPCR transgenic flies to the control group, analyzed using standard probit analysis with a computerized version of [63]. Each bioassay was independently replicated three times. All Drosophila were reared on Jazz-Mix Drosophila food (Fisher, KS City, MO, USA) at 25 ± 2 °C under a 12:12 (L:D) photoperiod, following standard protocols [64].
4.8. Expression of Drosophila Insecticide-Related P450 Genes in the House Fly GPCR Transgenic Drosophila Lines
qRT-PCR was used to detect the relative expression levels of four Drosophila resistance P450 genes, CYP12D1, CYP6G1, CYP6A2, and CYP6A8 [42] in CK, ALHF_02706.g1581 and ALHF_04422.g2918 transgenic Drosophila lines. Female Drosophila, 2–3 days post-eclosion, were collected for the P450 gene expression analysis, following the same qRT-PCR method described previously. The primers used for the PCR reactions for CYP12D1, CYP6G1, CYP6A2, and CYP6A8 are listed in Table 1.
5. Conclusions
The two GPCR genes ALHF_02706.g1581 and ALHF_04422.g2918 are significantly overexpressed in the insecticide-resistant strain of M. domestica (ALHF). When overexpressed in transgenic Drosophila through the GAL4/UAS system, these genes improved the resistance level of Drosophila to permethrin and promoted the expression of specific P450 genes. Both ALHF_02706.g1581 and ALHF_04422.g2918 mapped on autosome 2, which is associated with the regulation of P450 genes in house flies, indicating that GPCR genes play a critical role in the regulatory pathway of P450 genes in M. domestica. This study sheds new light on the potential function of GPCR genes in insecticide resistance in M. domestica. Future research should aim to provide more evidence on the function of GPCRs in insecticide resistance development and clarify the exact regulatory pathways involved. Gene knockdown and in vitro gene expression studies are suggested for future functional studies of GPCR genes in M. domestica. Understanding the downstream factors in the GPCR regulatory pathway will be crucial for identifying new insecticide targets, supporting the development of novel insecticides, and helping control house fly populations and house fly-borne diseases.
Author Contributions
Conceived and designed the study: N.L. Performed experiments and data analysis: J.X., D.B., Y.W., X.W., M.L. and T.L. Prepared the materials: N.L. Wrote the paper and discussed the paper: J.X., D.B., Y.W., X.W., M.L., T.L. and N.L. All authors have read and agreed to the published version of the manuscript.
Funding
The project was supported by the following awards: AAES Hatch/Multistate Grants ALA015-1-16009 and ALA015-1-19148 to N.L.
Institutional Review Board Statement
Not applicable. The study did not require ethical approval.
Informed Consent Statement
Not applicable. This article does not contain any studies with human participants or animals performed by any of the authors.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, N.L., upon request.
Acknowledgments
Authors thank the support from College of Agriculture, Auburn University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Geden, C.J.; Nayduch, D.; Scott, J.G.; Burgess, E.R.; Gerry, A.C.; Kaufman, P.E.; Thomson, J.; Pickens, V.; Machtinger, E.T. House fly (Diptera: Muscidae): Biology, pest status, current management prospects, and research needs. J. Integr. Pest. Manag. 2021, 12, 39. [Google Scholar] [CrossRef]
- Gioia, G.; Freeman, J.; Sipka, A.; Santisteban, C.; Wieland, M.; Gallardo, V.A.; Monistero, V.; Scott, J.G.; Moroni, P. Pathogens associated with houseflies from different areas within a New York State dairy. JDS Commun. 2022, 3, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Nayduch, D.; Burrus, R.G. Flourishing in filth: House fly–microbe interactions across life history. Ann. Entomol. Soc. Am. 2017, 110, 6–18. [Google Scholar] [CrossRef]
- Freeman, J.C.; Ross, D.H.; Scott, J.G. Insecticide resistance monitoring of house fly populations from the United States. Pestic. Biochem. Phys. 2019, 158, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yue, X. Genetics of pyrethroid resistance in a strain (ALHF) of house flies (diptera: Muscidae). Pestic. Biochem. Phys. 2001, 70, 151–158. [Google Scholar] [CrossRef]
- Liu, N.; Yue, X. Insecticide resistance and cross-resistance in the house fly (Diptera: Muscidae). J. Econ. Entomol. 2000, 93, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.G. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Li, M.; Reid, W.R.; Zhang, L.; Scott, J.G.; Gao, X.; Kristensen, M.; Liu, N. A whole transcriptomal linkage analysis of gene co-regulation in insecticide resistant house flies, Musca domestica. BMC Genom. 2013, 14, 803. [Google Scholar] [CrossRef]
- Meisel, R.P.; Freeman, J.C.; Asgari, D.; Llaca, V.; Fengler, K.A.; Mann, D.; Rastogi, A.; Loso, M.; Geng, C.; Scott, J.G. New insights into immune genes and other expanded gene families of the house fly, Musca domestica, from an improved whole genome sequence. Arch. Insect Biochem. Physiol. 2023, 114, e22049. [Google Scholar] [CrossRef]
- Scott, J.G.; Warren, W.C.; Beukeboom, L.W.; Bopp, D.; Clark, A.G.; Giers, S.D.; Hediger, M.; Jones, A.K.; Kasai, S.; Leichter, C.A.; et al. Genome of the house fly, Musca domestica L., a global vector of diseases with adaptations to a septic environment. Genome Biol. 2014, 15, 466. [Google Scholar] [CrossRef]
- Li, M.; Feng, X.; Reid, W.R.; Tang, F.; Liu, N. Multiple-P450 gene co-up-regulation in the development of permethrin resistance in the house fly, Musca domestica. Int. J. Mol. Sci. 2023, 24, 3170. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect cytochrome P450. In Comprehensive Molecular Insect Science; Elsevier: Amsterdam, The Netherlands, 2005; pp. 1–77. [Google Scholar]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Br, M.; Mk, S.; Mn, O.; Oc, J.; Ris, A.; Rd, F. Understanding the mechanisms involved in the regulation of cytochrome p450 gene expression in Drosophila melanogaster (Diptera: Drosophilidae). Entomol. Ornithol. Herpetol. 2017, 6. [Google Scholar] [CrossRef]
- Carino, F.A.; Koener, J.F.; Plapp, F.W., Jr.; Feyereisen, R. Constitutive overexpression of the cytochrome P450 gene CYP6A1 in a house fly strain with metabolic resistance to insecticides. Insect Biochem. Mol. Biol. 1994, 24, 411–418. [Google Scholar] [CrossRef]
- Kalsi, M.; Palli, S.R. Transcription factors, CncC and Maf, regulate expression of CYP6BQ genes responsible for deltamethrin resistance in Tribolium castaneum. Insect Biochem. Mol. Biol. 2015, 65, 47–56. [Google Scholar] [CrossRef]
- Liu, N.; Li, M.; Gong, Y.; Liu, F.; Li, T. Cytochrome P450s—Their expression, regulation, and role in insecticide resistance. Pestic. Biochem. Phys. 2015, 120, 77–81. [Google Scholar] [CrossRef]
- Liu, N.; Scott, J.G. Phenobarbital induction of CYP6D1 is due to a trans acting factor on autosome 2 in house flies, Musca domestica. Insect Mol. Biol. 1997, 6, 77–81. [Google Scholar] [CrossRef]
- Liu, N.; Scott, J.G. Genetics of resistance to pyrethroid insecticides in the house fly, Musca domestica. Pestic. Biochem. Phys. 1995, 52, 116–124. [Google Scholar] [CrossRef]
- McDonnell, C.M.; Brown, R.P.; Berenbaum, M.R.; Schuler, M.A. Conserved regulatory elements in the promoters of two allelochemical-inducible cytochrome P450 genes differentially regulate transcription. Insect Biochem. Mol. Biol. 2004, 34, 1129–1139. [Google Scholar] [CrossRef]
- Pu, J.; Sun, H.; Wang, J.; Wu, M.; Wang, K.; Denholm, I.; Han, Z. Multiple cis-acting elements involved in up-regulation of a cytochrome P450 gene conferring resistance to deltamethrin in smal brown planthopper, Laodelphax striatellus (Fallén). Insect Biochem. Mol. Biol. 2016, 78, 20–28. [Google Scholar] [CrossRef]
- Smith, L.B.; Tyagi, R.; Kasai, S.; Scott, J.G. CYP-mediated permethrin resistance in Aedes aegypti and evidence for trans-regulation. PLoS Negl. Trop. Dis. 2018, 12, e0006933. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.; Wu, S.; Ling, L.; Danao, J.; Li, Y.; Yeh, W.C.; Tian, H.; Baribault, H. G-protein-coupled receptor GPR21 knockout mice display improved glucose tolerance and increased insulin response. Biochem. Biophys. Res. Commun. 2012, 418, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Millar, R.P.; Newton, C.L. The year in G protein-coupled receptor research. Mol. Endocrinol. 2010, 24, 261–274. [Google Scholar] [CrossRef]
- Syrovatkina, V.; Alegre, K.O.; Dey, R.; Huang, X.Y. Regulation, signaling, and physiological functions of G-proteins. J. Mol. Biol. 2016, 428, 3850–3868. [Google Scholar] [CrossRef]
- Bai, H.; Zhu, F.; Shah, K.; Palli, S.R. Large-scale RNAi screen of G protein-coupled receptors involved in larval growth, molting and metamorphosis in the red flour beetle. BMC Genom. 2011, 12, 388. [Google Scholar] [CrossRef]
- Terhzaz, S.; Cabrero, P.; Robben, J.H.; Radford, J.C.; Hudson, B.D.; Milligan, G.; Dow, J.A.; Davies, S.A. Mechanism and function of Drosophila capa GPCR: Adesiccation stress-responsive receptor with functionalhomology to human neuromedinU receptor. PLoS ONE 2012, 7, e29897. [Google Scholar] [CrossRef]
- Kersch, C.N.; Pietrantonio, P.V. Mosquito Aedes aegypti (L.) leucokinin receptor is critical for in vivo fluid excretion post blood feeding. FEBS Lett. 2011, 585, 3507–3512. [Google Scholar] [PubMed]
- Li, T.; Cao, C.; Yang, T.; Zhang, L.; He, L.; Xi, Z.; Bian, G.; Liu, N. A G-protein-coupled receptor regulation pathway in cytochrome P450-mediated permethrin-resistance in mosquitoes, Culex quinquefasciatus. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, L.; Zhang, L.; Liu, N. Role of G-protein-coupled receptor-related genes in insecticide resistance of the mosquito, Culex quinquefasciatus. Sci. Rep. 2014, 4, 6474. [Google Scholar] [CrossRef]
- Liu, N.; Li, T.; Wang, Y.; Liu, S. G-protein coupled receptors (GPCRs) in insects—A potential target for new insecticide development. Molecules 2021, 26, 2993. [Google Scholar] [CrossRef]
- Liu, N.; Wang, Y.; Li, T.; Feng, X. G-protein coupled receptors (GPCRs): Signaling pathways, characterization, and functions in insect physiology and toxicology. Int. J. Mol. Sci. 2021, 22, 5260. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, N. Role of the G-protein-coupled receptor signaling pathway in insecticide resistance. Int. J. Mol. Sci. 2019, 20, 4300. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, N. The function of G-protein-coupled receptor-regulatory cascade in southern house mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2018, 55, 862–870. [Google Scholar] [CrossRef]
- Li, T.; Liu, N. Regulation of P450-mediated permethrin resistance in Culex quinquefasciatus by the GPCR/Gαs/AC/cAMP/PKA signaling cascade. Biochem. Biophys. Rep. 2017, 12, 12–19. [Google Scholar] [CrossRef]
- Meisel, R.P.; Scott, J.G. Using genomic data to study insecticide resistance in the house fly, Musca domestica. Pestic. Biochem. Phys. 2018, 151, 76–81. [Google Scholar] [CrossRef]
- Schaeffer, S.W. Muller “Elements” in Drosophila: How the search for the genetic basis for speciation led to the birth of comparative genomics. Genetics 2018, 210, 3–13. [Google Scholar] [CrossRef]
- Riddle, N.C.; Elgin, S.C.R. The Drosophila dot chromosome: Where genes flourish amidst repeats. Genetics 2018, 210, 757–772. [Google Scholar] [CrossRef]
- Schaeffer, S.W.; Bhutkar, A.; McAllister, B.F.; Matsuda, M.; Matzkin, L.M.; O’Grady, P.M.; Rohde, C.; Valente, V.L.; Aguadé, M.; Anderson, W.W.; et al. Polytene chromosomal maps of 11 Drosophila species: The order of genomic scaffolds inferred from genetic and physical maps. Genetics 2008, 179, 1601–1655. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C. BLAST plus: Architecture and applications. BMC Bioinform. 2009, 10, 1. [Google Scholar] [CrossRef]
- Zhu, F.; Liu, N. Differential expression of CYP6A5 and CYP6A5v2 in pyrethroid-resistant house flies, Musca domestica. Arch. Insect Biochem. Physiol. 2008, 67, 107–119. [Google Scholar] [CrossRef]
- Daborn, P.J.; Lumb, C.; Boey, A.; Wong, W.; Ffrench-Constant, R.H.; Batterham, P. Evaluating the insecticide resistance potential of eight Drosophila melanogaster cytochrome P450 genes by transgenic over-expression. Insect Biochem. Mole. Biol. 2007, 37, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Palli, S.R. CncC/Maf-mediated xenobiotic response pathway in insects. Arch. Insect Biochem. Physiol. 2020, 104, e21674. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Norris, E.J.; Jiang, S.; Bernier, U.R.; Linthicum, K.J.; Bloomquist, J.R. Reduced effectiveness of repellents in a pyrethroid-resistant strain of Aedes aegypti (Diptera: Culicidae) and its correlation with olfactory sensitivity. Pest. Manag. Sci. 2020, 76, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Huang, H.; Hu, S.; Ren, M.; Wei, Q.; Tian, X.; Elzaki, M.E.A.; Bass, C.; Su, J.; Palli, S.R. Changes in both trans- and cis-regulatory elements mediate insecticide resistance in a lepidopteron pest, Spodoptera exigua. PLOS Genet. 2021, 17, e1009403. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Sun, L.; Du, H.; Moural, T.W.; Bai, H.; Liu, P.; Zhu, F. Physiological functions of a methuselah-like G protein coupled receptor in Lymantria dispar Linnaeus. Pestic. Biochem. Phys. 2019, 160, 1–10. [Google Scholar] [CrossRef]
- Sun, Y.; Zou, P.; Yu, X.Y.; Chen, C.; Yu, J.; Shi, L.N.; Hong, S.C.; Zhou, D.; Chang, X.L.; Wang, W.J.; et al. Functional characterization of an arrestin gene on insecticide resistance of Culex pipiens pallens. Parasit. Vectors 2012, 5, 134. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, Y.; You, C.; Zeng, X.; Gao, X. The role of G protein-coupled receptor-related genes in cytochrome P450-mediated resistance of the house fly, Musca domestica (Diptera: Muscidae), to imidacloprid. Insect Mol. Biol. 2020, 29, 92–103. [Google Scholar] [CrossRef]
- Chung, H.; Bogwitz, M.R.; McCart, C.; Andrianopoulos, A.; Ffrench-Constant, R.H.; Batterham, P.; Daborn, P.J. Cis-regulatory elements in the Accord retrotransposon result in tissue-specific expression of the Drosophila melanogaster insecticide resistance gene Cyp6g1. Genetics 2007, 175, 1071–1077. [Google Scholar] [CrossRef]
- Maitra, S.; Dombrowski, S.M.; Waters, L.C.; Ganguly, R. Three second chromosome-linked clustered Cyp6 genes show differential constitutive and barbital-induced expression in DDT-resistant and susceptible strains of Drosophila melanogaster. Gene 1996, 180, 165–171. [Google Scholar] [CrossRef]
- Tian, L.; Cao, C.; He, L.; Li, M.; Zhang, L.; Zhang, L.; Liu, H.; Liu, N. Autosomal interactions and mechanisms of pyrethroid resistance in house flies, Musca domestica. Int. J. Biol. Sci. 2011, 7, 902–911. [Google Scholar] [CrossRef]
- Gao, J.; Scott, J.G. Role of the transcriptional repressor mdGfi-1 in CYP6D1v1-mediated insecticide resistance in the house fly, Musca domestica. Insect Biochem. Mol. Biol. 2006, 36, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Liu, H.; Zhu, F.; Zhang, L. Differential expression of genes in pyrethroid resistant and susceptible mosquitoes, Culex quinquefasciatus (S.). Gene 2007, 394, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Aerts, J.L.; Gonzales, M.I.; Topalian, S.L. Selection of appropriate control genes to assess expression of tumor antigens using real-time RT-PCR. Biotechniques 2004, 36, 84–91. [Google Scholar] [CrossRef]
- Feng, X.; Li, M.; Liu, N. Carboxylesterase genes in pyrethroid resistant house flies, Musca domestica. Insect Biochem. Mol. Biol. 2018, 92, 30–39. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Strode, C.; Wondji, C.S.; David, J.P.; Hawkes, N.J.; Lumjuan, N.; Nelson, D.R.; Drane, D.R.; Karunaratne, S.H.; Hemingway, J.; Black, W.C.; et al. Genomic analysis of detoxification genes in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 113–123. [Google Scholar] [CrossRef]
- Foster, G.G.; Whitten, M.J.; Konovalov, C.; Arnold, J.T.A.; Maffi, G. Autosomal genetic maps of the Australian Sheep Blowfly, Lucilia cuprina dorsalis R.-D. (Diptera: Calliphoridae), and possible correlations with the linkage maps of Musca domestica L. and Drosophila melanogaster (Mg.). Genet. Res. 1981, 37, 55–69. [Google Scholar] [CrossRef]
- Vicoso, B.; Bachtrog, D. Numerous transitions of sex chromosomes in Diptera. PLoS Biol. 2015, 13, e1002078. [Google Scholar] [CrossRef]
- Bischof, J.; Maeda, R.K.; Hediger, M.; Karch, F.; Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific φC31 integrases. Proc. Natl. Acad. Sci. USA 2007, 104, 3312–3317. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [CrossRef]
- Bateman, J.R.; Lee, A.M.; Wu, C.T. Site-specific transformation of Drosophila via ϕC31 integrase-mediated cassette exchange. Genetics 2006, 173, 769–777. [Google Scholar] [CrossRef]
- Raymond, M. Prsentation d’un programme basic d’analyse log-probit pour micro-ordinateur. ORSTOM Ser. Entomol. Med. Parasitol. 1985, 23, 117–121. [Google Scholar]
- Ashburner, M. Drosophila: A Laboratory Handbook; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2005; Volume 52, 1448p. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).