Abstract
Andrographis paniculata is known for its diterpenoid medicinal compounds with antibacterial and anti-inflammatory properties. However, it faces production and cultivation challenges due to low temperatures (LTs). Cytochrome P450 monooxygenases (CYPs) are key enzymes in diterpenoid accumulation. Nevertheless, the functions and LT-related expression patterns of diterpenoid pathway CYPs in Andrographis paniculata remain poorly understood. In this study, 346 CYPs were discovered in Andrographis paniculata. Among them, 328 CYPs belonged to 42 known subfamilies. The remaining 17 CYPs might have represented novel subfamilies unique to this species. A total of 65 candidate CYPs associated with diterpenoid modification were identified. Of these, 50 were transmembrane proteins, and 57 were localized to chloroplasts. The CYP71 subfamily was the most abundant and had the highest motif diversity. Promoters of all candidate CYPs commonly contained elements responsive to gibberellins (GAs), methyl jasmonate (MeJA), and abiotic stresses. Notably, the XP_051152769 protein, corresponding to a CYP gene over 40,000 bp in length, featured an extraordinarily long intron (40,751 nts). Functional elements within this intron were related to LT, GAs, and dehydration pathways. Based on the promoter element arrangement and subfamily classification, 10 representative candidate CYPs were selected. Under LT stress, significant expression changes were observed in three representative CYPs: CYP71D, ent-kaurenoic acid oxidase (KAO), and ent-kaurene oxidase (KO). KAO and KO were significantly upregulated during early LT stress. KAO and KO interacted with each other and jointly interacted with GA20OX2-like. CYP71D acted as a negative response factor to LT stress. Among the 37 proteins interacting with CYP71D, 95% were CYPs. This study provides a critical preliminary foundation for investigating the functions of diterpenoid pathway CYPs in Andrographis paniculata, thereby facilitating the development of LT-tolerant cultivars.
1. Introduction
Andrographis paniculata is a common medicinal plant belonging to the Acanthaceae family within the Lamiales order. Native to the tropical regions of India and Southeast Asia, Andrographis paniculata is extensively cultivated in the Guangdong, Fujian, and Hainan provinces of China [1]. The primary active component, andrographolide (C20H30O5), is renowned for its significant anti-inflammatory and antipyretic properties [2]. Andrographolide is a labdane-type diterpenoid biosynthesized from the precursor geranylgeranyl pyrophosphate (GGPP). GGPP is produced via two pathways: the cytoplasmic mevalonate (MVA) pathway and the plastidial 2-C-methyl-D-erythritol-4-phosphate (MEP) pathway [3]. The subsequent synthesis of diterpenoids is predominantly catalyzed by enzymes from the Cytochrome P450 (CYP450) family. The CYP450 family includes CYPs and Cytochrome P450 reductases (CPRs) [4]. CYPs enhance the hydrophilicity of substrates by introducing an oxygen atom. CPRs are crucial for the reductive activation of CYPs by transferring electrons from NADPH to CYPs, serving as the rate-limiting enzymes. CYPs perform various functions, such as hydroxylation, epoxidation, deamination, dehydrogenation, dealkylation, and oxidative C-C bond cleavage [5].
In 2011, Nelson and Werck-Reichhart categorized the CYP family based on amino acid sequence similarities. Sequences with over 40% similarity are grouped into the same family, and those with over 55% similarity are grouped into the same subfamily [6]. Plant CYPs encompass 11 families, including seven single-branch families (CYP51, CYP74, CYP97, CYP710, CYP711, CYP727, and CYP746) and four multi-branch families (CYP71, CYP72, CYP85, and CYP86) [7]. Notably, members of the CYP71 family in terrestrial plants are essential in the modification of diterpenoids. These include the subfamilies CYP71, CYP76AH, CYP76AK, CYP76BK, CYP99A, CYP701A (KO), and CYP726A [8]. Within the CYP71D subfamily, representatives have been identified in Fontainea as excellent candidates for diterpenoid synthesis [9]. CYP76AH, CYP76AK, and CYP76BK are specific to the Lamiaceae family [10]. CYP99A has been identified only in rice (Oryza sativa) [11]. CYP726A is unique to the Euphorbiaceae family [12]. Members of the CYP72 family’s CYP714 subfamily and the CYP85 family’s CYP88A (KAO), CYP720B, and CYP725A subfamilies also contribute to diterpenoid modification [8]. CYP720B and CYP725A are exclusive to gymnosperms [13]. Subfamilies KO, CYP714, and KAO participate in the catalytic modification of the diterpenoid plant hormones GAs [14]. GAs are crucial endogenous hormones that promote plant growth and development. GGPP undergoes two catalytic steps to form ent-kaurene, which KO enzymatically converts to ent-kaurenoic acid, subsequently transformed into GA12 by KAO [15]. Members of the CYP714 subfamily catalyze the inactivation of GAs by targeting the C-13 position or adjacent carbons of GAs and/or ent-kaurenoic acid [16]. Consequently, the significance of the diterpenoid pathway CYPs in plants is clearly evident.
LTs severely restrict plant growth and yield globally, limiting geographic distribution and posing threats to agricultural development [17]. The results of this study indicated that andrographolide was primarily stored in the leaves. Its concentration significantly decreased when the leaves were exposed to prolonged LT. This exposure caused the leaves to change color from green to red. Therefore, analyzing the response of diterpenoid pathway CYPs in Andrographis paniculata to LT stress is vital for enhancing its yield and environmental adaptability. This study identified and classified the diterpenoid pathway CYPs in Andrographis paniculata. Subsequently, functional annotation and gene expression profiling under LT stress were performed. It establishes an essential molecular foundation for investigating the pathways of andrographolide and GAs in Andrographis paniculata.
2. Results
2.1. Identification and Classification of Total CYPs in Andrographis Paniculata
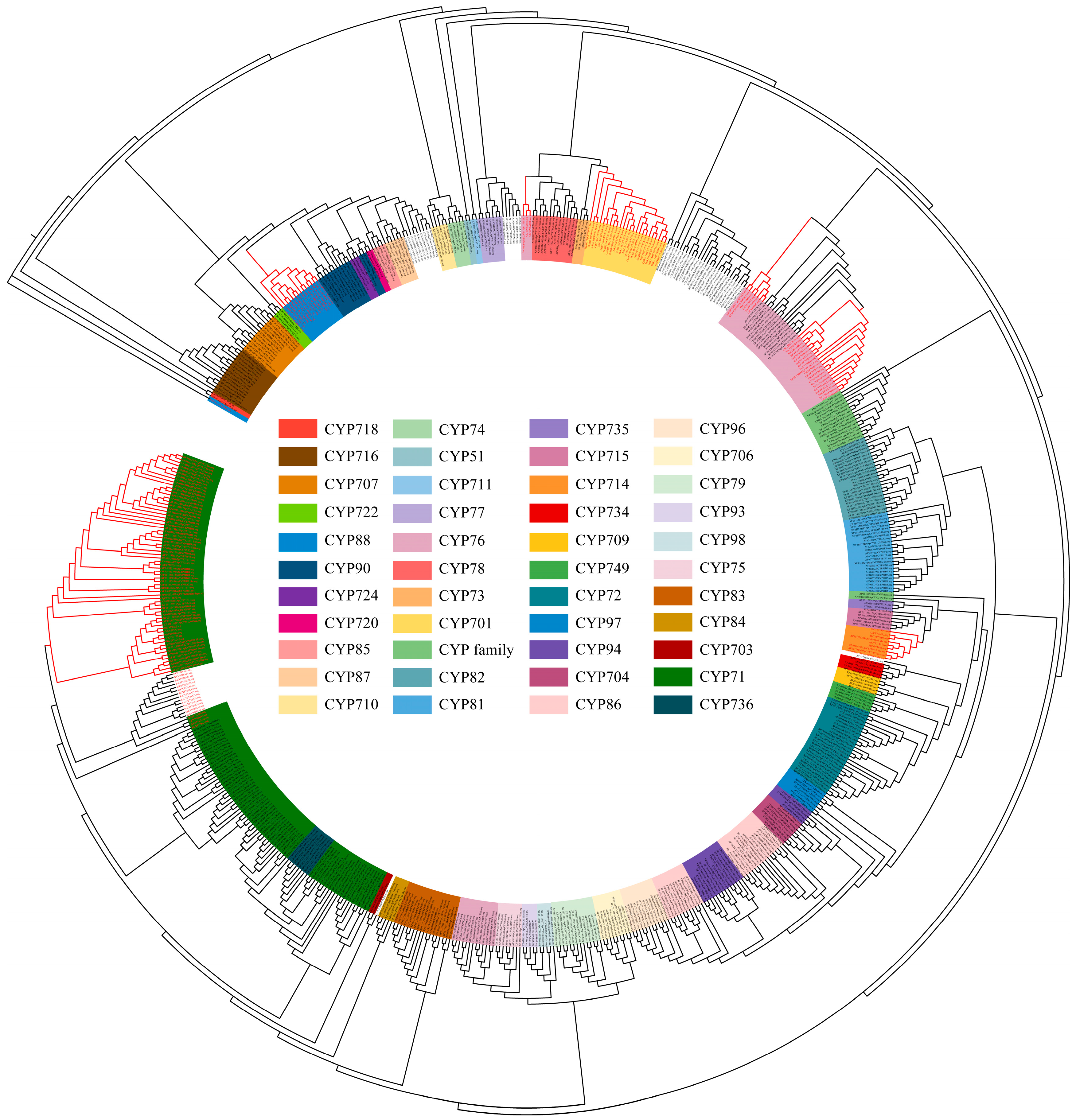
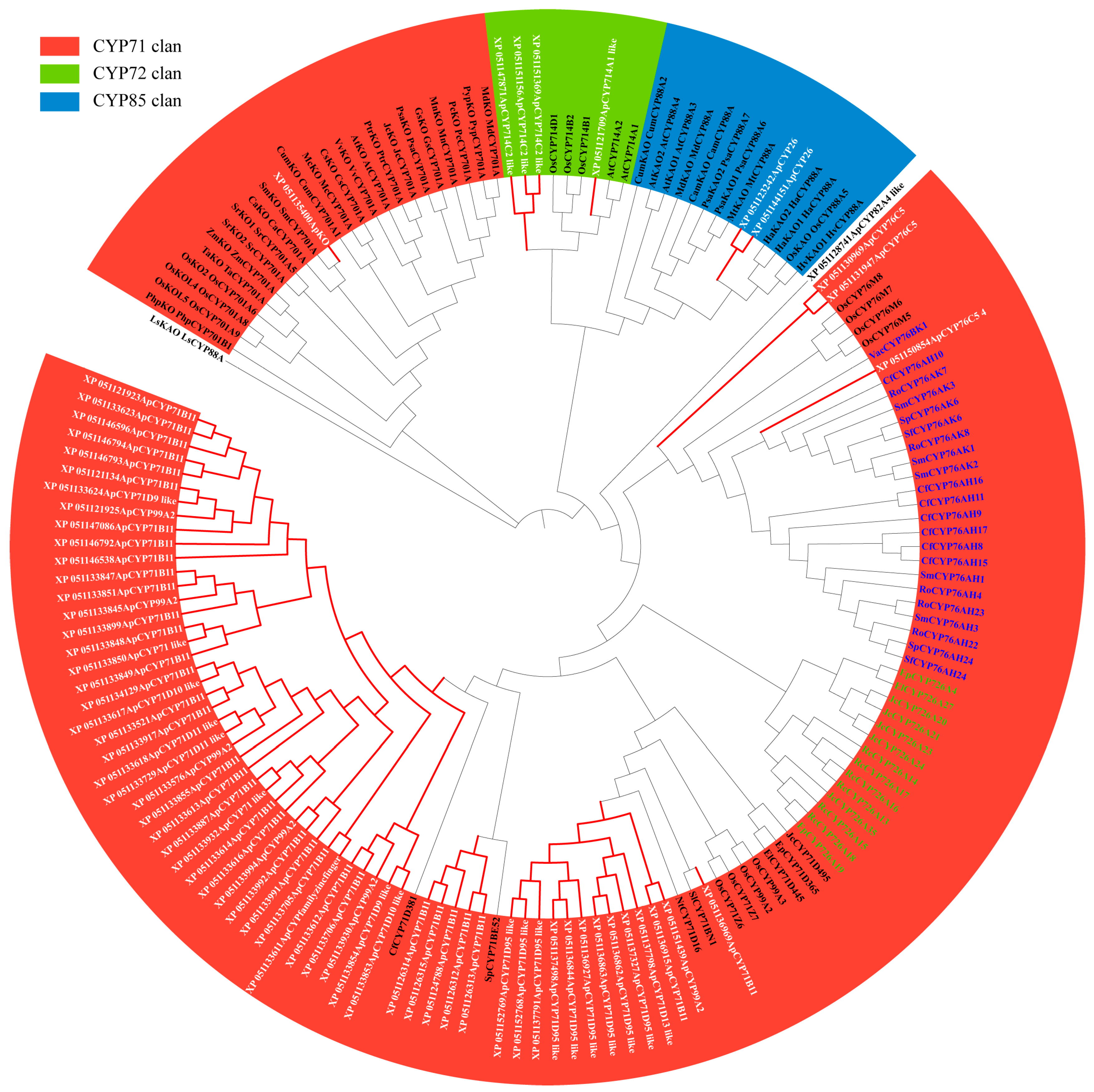
Based on the plant CYP HMM model, NCBI, and Interpro annotations, 346 CYPs in Andrographis paniculata were identified. These were analyzed alongside 92 terrestrial plant diterpenoid pathway CYPs (excluding gymnosperm-specific subfamilies) [8] and 230 Arabidopsis (Arabidopsis thaliana) CYPs, totaling 668 proteins, for phylogenetic relationships (Table S1). The MUSCLE alignment excluded two excessively short proteins (IDs in 4.2.), leaving 666 proteins for further analysis. Their conserved sequences were used to construct the phylogenetic tree. According to Figure 1 and Table 1, the 666 CYPs clustered into 50 subfamilies. Among these, 328 CYPs from Andrographis paniculata were assigned to 42 known subfamilies. The remaining 17 CYPs did not cluster into any recognized subfamily and were annotated as ‘CYP family’. Subfamilies CYP702 and CYP708 are unique to Arabidopsis [18], while subfamilies CYP736 and CYP749 contain only Andrographis paniculata CYPs. Diterpenoid pathway CYPs originated from the CYP71 family’s CYP71, CYP76, CYP99, and CYP701A subfamilies, the CYP72 family’s CYP714 subfamily, and the CYP85 family’s CYP88A subfamily. The CYP99 subfamily has been identified only in monocots [19]. In the dicot Andrographis paniculata, six CYPs were annotated as CYP99 but clustered within the CYP71 subfamily. These CYPs were reassigned to the CYP71 subfamily (Figure 1 and Table S2). A total of 70 Andrographis paniculata CYPs that clustered with terrestrial plant diterpenoid pathway CYPs (marked in the red clade in Figure 1) were selected as candidates.
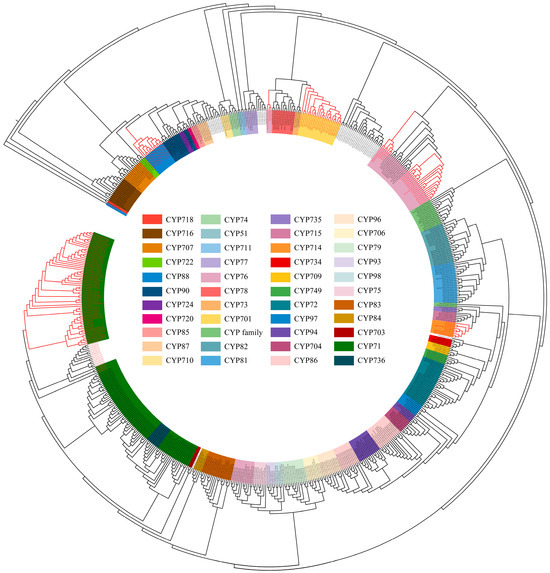
Figure 1.
Phylogenetic tree of conserved CYP protein sequences in Arabidopsis (total CYPs), Andrographis paniculata (total CYPs), and terrestrial plants (diterpenoid pathway CYPs) Note: red text—diterpenoid pathway CYPs in Andrographis paniculata and terrestrial plants; red branches—clades containing Andrographis paniculata CYPs within the diterpenoid pathway.

Table 1.
Distribution of CYP subfamilies.
2.2. Identification of Diterpenoid Pathway CYPs in Andrographis Paniculata
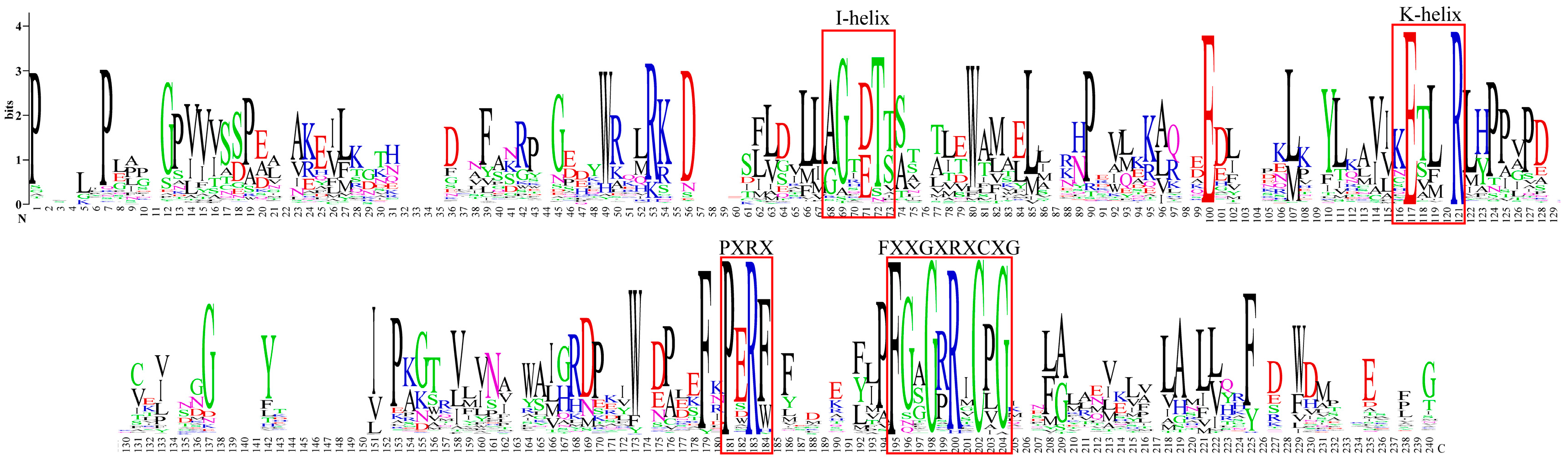
Phylogenetic analysis was conducted on 70 candidate CYPs from Andrographis paniculata alongside 92 terrestrial plant diterpenoid pathway CYPs using full-length protein sequences (Figure 2). XP_051128741 did not cluster with any subfamily and was excluded. XP_051150854 clustered with the CYP76AH, CYP76AK, and CYP76BK subfamilies, which are specifically expanded in the Lamiaceae of Lamiales. This suggested that these subfamilies might have a catalytic function unique to Lamiales rather than being restricted to Lamiaceae. Plant CYPs typically contain the I-helix, K-helix, and a critical domain near the C-terminus, FXXGXRXCXG [20]. In Andrographis paniculata, all 345 identified CYPs contained the I-helix and K-helix, but some lacked the complete FXXGXRXCXG. Based on the FXXGXRXCXG motif, 22 sequences were excluded (IDs: XP_051123363, XP_051127248, XP_051128741, XP_051133618, XP_051136989, XP_051140631, XP_051152697, XP_051123609, XP_051125649, XP_051126314, XP_051150718, XP_051131936, XP_051131945, XP_051133611, XP_051137552, XP_051141267, XP_051141281, XP_051146792, XP_051147073, XP_051149418, XP_051149781, and XP_051150050), leaving 323 high-confidence CYPs (Figure 3). Thus, the 70 candidate diterpenoid pathway CYPs were reduced to 65.
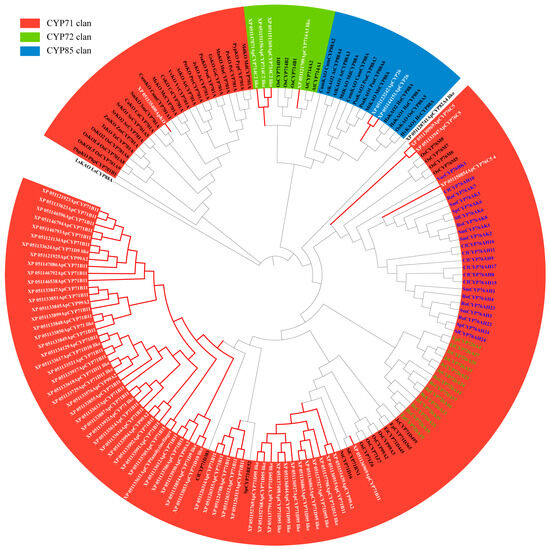
Figure 2.
Phylogenetic tree of full-length CYP protein sequences from diterpene pathways in Andrographis paniculata and terrestrial plants Note: green text—Euphorbiaceae-specific expanded subfamily CYP726A; blue text—Lamiaceae-specific expanded subfamily CYP76AH, CYP76AK, and CYP76BK; red branches (white text)—Andrographis paniculata candidate diterpenoid pathway CYPs; black branches—terrestrial plants diterpenoid pathway CYPs.
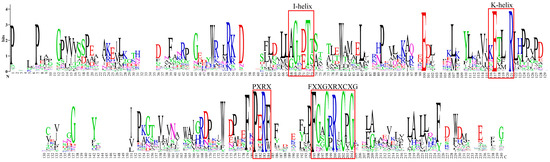
Figure 3.
Conserved domains of 323 Andrographis paniculata CYPs. Note: red squares—conserved domains.
2.3. Analysis of Basic Information for Diterpenoid Pathway CYPs
Based on database annotations and phylogenetic classification, the symbols of 65 candidate diterpenoid pathway CYPs were revised (Table S2). Except for XP_051133612 (267 amino acids, aa) and XP_051136915 (620 aa), the lengths of the remaining 63 candidate CYPs were evenly distributed around 500 aa, with an average length of 503 aa. The two members of the CYP88A subfamily, four members of the CYP714 subfamily, and three members of the CYP76 subfamily were localized to the chloroplasts. The sole member of the CYP701A subfamily, XP_051135400 (KO), was localized to the plasma membrane. Of the 55 CYP71 subfamily members, 48 were localized to the chloroplasts. XP_051121925, XP_051133624, and XP_051121134 were localized to the plasma membrane. XP_051126315 and XP_051133729 were found in the vacuolar membrane, while XP_051133853 and XP_051133854 are localized to the endoplasmic reticulum. These results indicated that the 65 candidate CYPs primarily catalyze products of the MEP pathway. A total of 50 candidate CYPs were transmembrane proteins, with 47 CYPs having a transmembrane domain within the first 30 aa. The remaining three candidate CYPs had two transmembrane domains. However, 57 candidate CYPs lacked signal peptides, while the remaining eight were predicted to have signal peptide sequences with a probability of only 51–69%.
2.4. Analysis of Feature Information for Diterpenoid Pathway CYPs
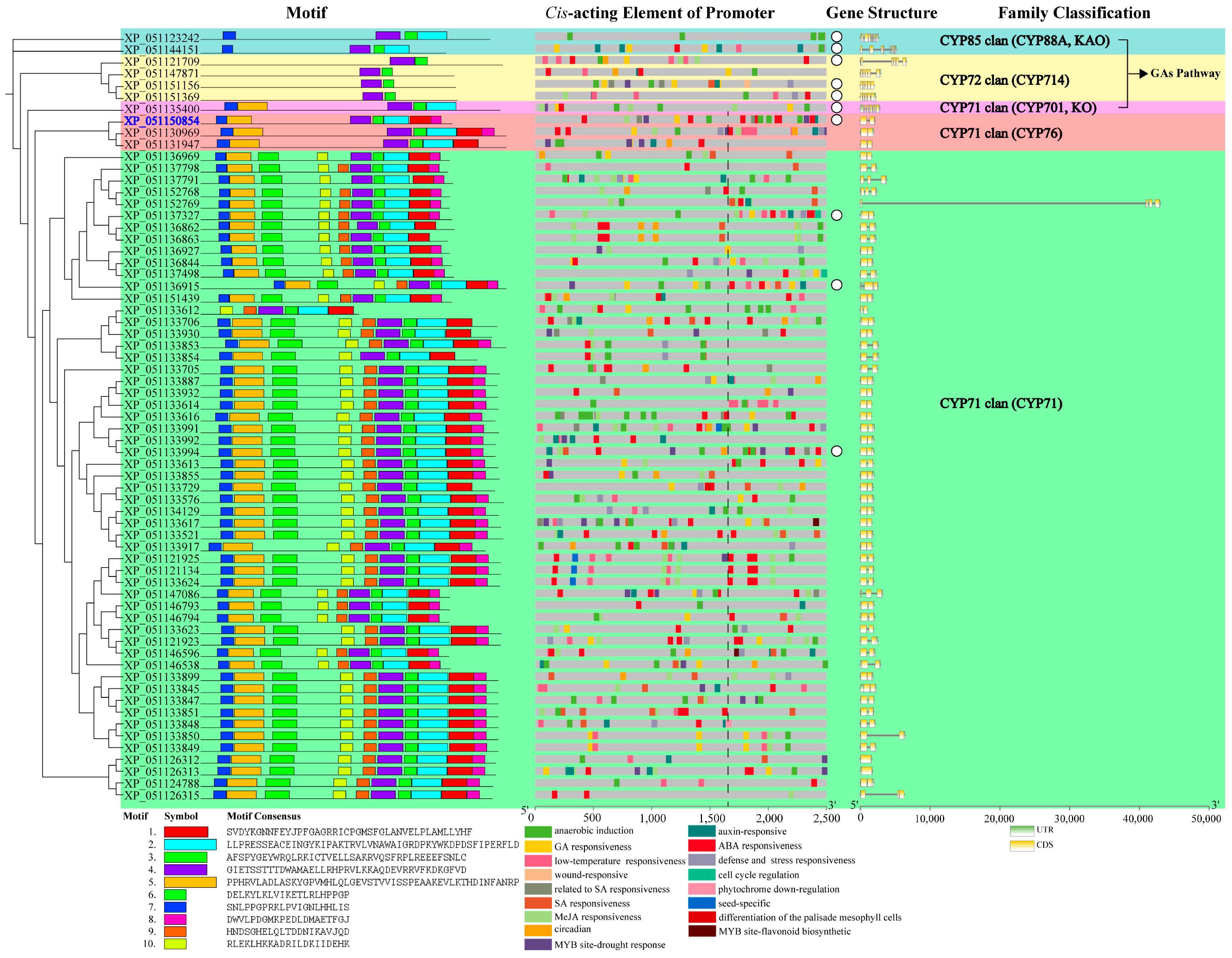
Motif distribution within candidate CYPs was subfamily-specific, elucidating systematic evolutionary relationships (Figure 4). CYPs in the three GA pathway subfamilies exhibited fewer motifs compared to those in the CYP76 and CYP71 subfamilies. The CYP714 subfamily contained only motifs 4 and 6. The CYP88A subfamily included motifs 2, 4, 6, and 7. The CYP701A subfamily had an additional motif 5 compared to CYP88A. The CYP76 subfamily conservatively included motifs 1, 2, 4, 5, 6, and 7. Within the CYP71 subfamily, all members, except for the significantly truncated XP_051133612, consistently contained motifs 1, 2, 3, 4, 5, 6, 7, and 10.
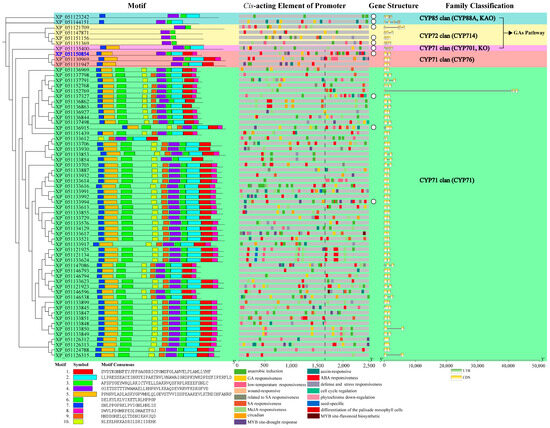
Figure 4.
The distribution of protein motifs, promoter cis-acting elements, and gene structures of candidate diterpenoid pathway CYPs Note: blue ID—candidate CYP clustered in the Lamiaceae-specific expanded CYP subfamily; promoter region to the right of the dashed line—core and proximal promoters; white circles—10 representative candidate CYPs.
Cis-acting elements in promoters determine the biological function of genes. According to Figure 4 and Table 2, GA-responsive elements were not limited to CYP88A, CYP701A, and CYP714 subfamilies; CYPs in the CYP76 and CYP71 subfamilies could also respond to the GA pathway. Additionally, candidate CYPs generally responded to abscisic acid (ABA) and anaerobic induction. ABA is involved in stress response, while anaerobic induction is associated with flooding damage. Except for one CYP in the CYP701A subfamily, all other subfamilies had CYPs participating in auxin signaling and environmental stresses such as LT. MeJA, related to damage, induces chemical defense. In addition to two CYPs in the CYP88A subfamily, the other four subfamilies included 49 candidate CYPs with promoters containing MeJA-responsive elements. Two CYP71 subfamily CYPs (protein IDs: XP_051121134 and XP_051133624) had the highest number of MeJA-responsive elements, each with seven. Another two CYP71 subfamily candidate CYPs (protein IDs: XP_051133617 and XP_051146596) had promoters containing flavonoid biosynthesis elements.
Table 2.
Promoter cis-elements of candidate CYPs involved in the diterpenoid pathway in Andrographis paniculata.
Research on wheat indicates that gene function is determined not only by exons and non-coding regions but also by introns, which significantly affect protein efficacy [21]. Our structural analysis showed that one candidate CYP in the CYP71 subfamily (protein ID: XP_051152769) contained an unusually long intron of 40,751 bp. According to Table S3, this long intron included multiple functionally decisive cis-acting elements involved in responses to light, LT, GA signaling, dehydration, and phosphate starvation. The lengths of other candidate CYPs were all within 10,000 bp.
2.5. Structural Analysis of Representative Diterpenoid Pathway CYPs
The promoter is divided into core, proximal, and distal regions. The core promoter contains the transcriptional start site (TSS), while the proximal promoter is located near the TSS. Both are primary binding areas for specific transcription factors (TFs). Conversely, the influence of TFs bound to the distal promoter is relatively weaker. In this study, the region located at the upstream one-third of the promoter from the start codon was designated as the core and proximal promoter. We selected 10 representative candidate CYPs from each subfamily whose cis-acting elements were predominantly located within the core and proximal promoters. These were marked by white circles in Figure 4. The protein IDs of the 10 representative candidate CYPs were as follows: XP_051121709, XP_051151156, and XP_051151369 of the CYP714 subfamily; XP_051123242 and XP_051144151 of the CYP88A subfamily; XP_051135400 of the CYP701A subfamily; XP_051150854 of the CYP76 subfamily; and XP_051136915 (CYP71B), XP_051137327 (CYP71D), and XP_051133994 (CYP99) of the CYP71 subfamily. The spatial structures of representative candidate CYPs in the GA pathway subfamilies were relatively conserved. Within the CYP71 subfamily, structural discrepancies existed between CYP71D and the other three members, indicating functional specificity (Figure 5).

Figure 5.
3D and 2D structures of representative candidate CYPs.
2.6. Protein Interactions of LT-Responsive Representative CYPs
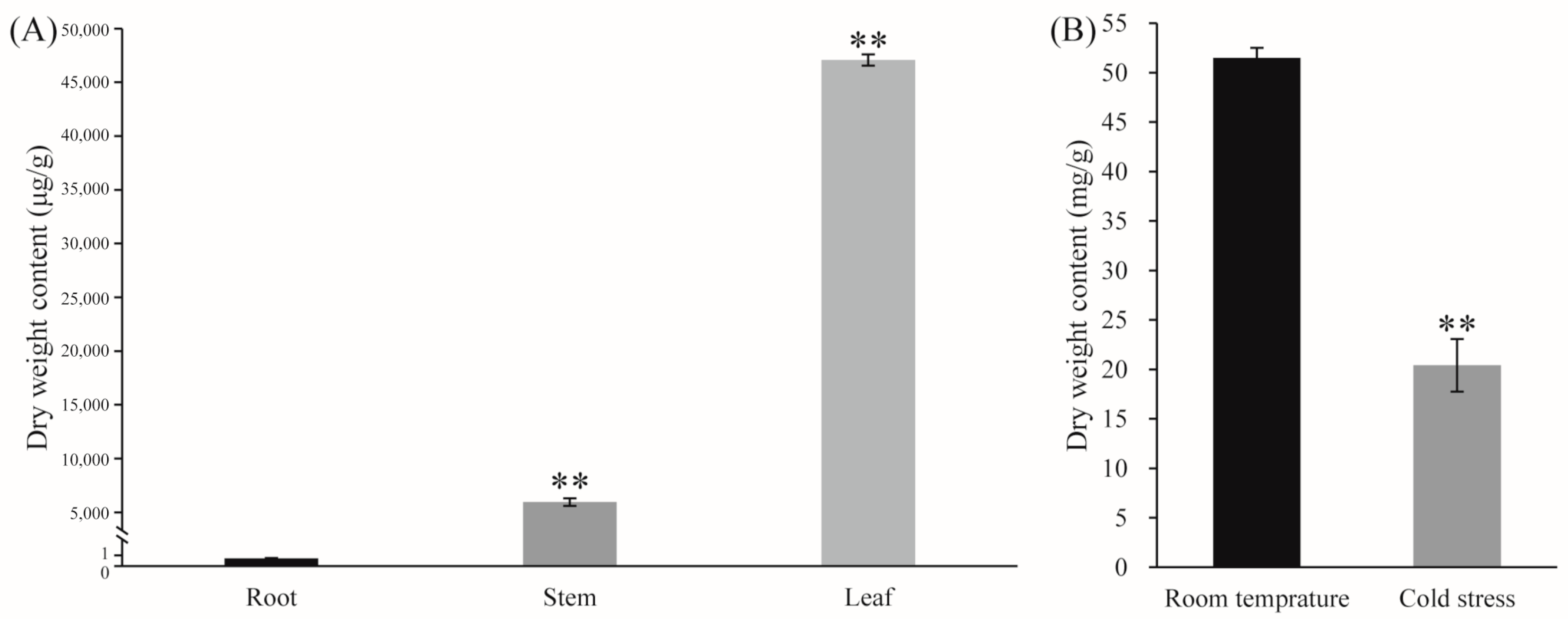
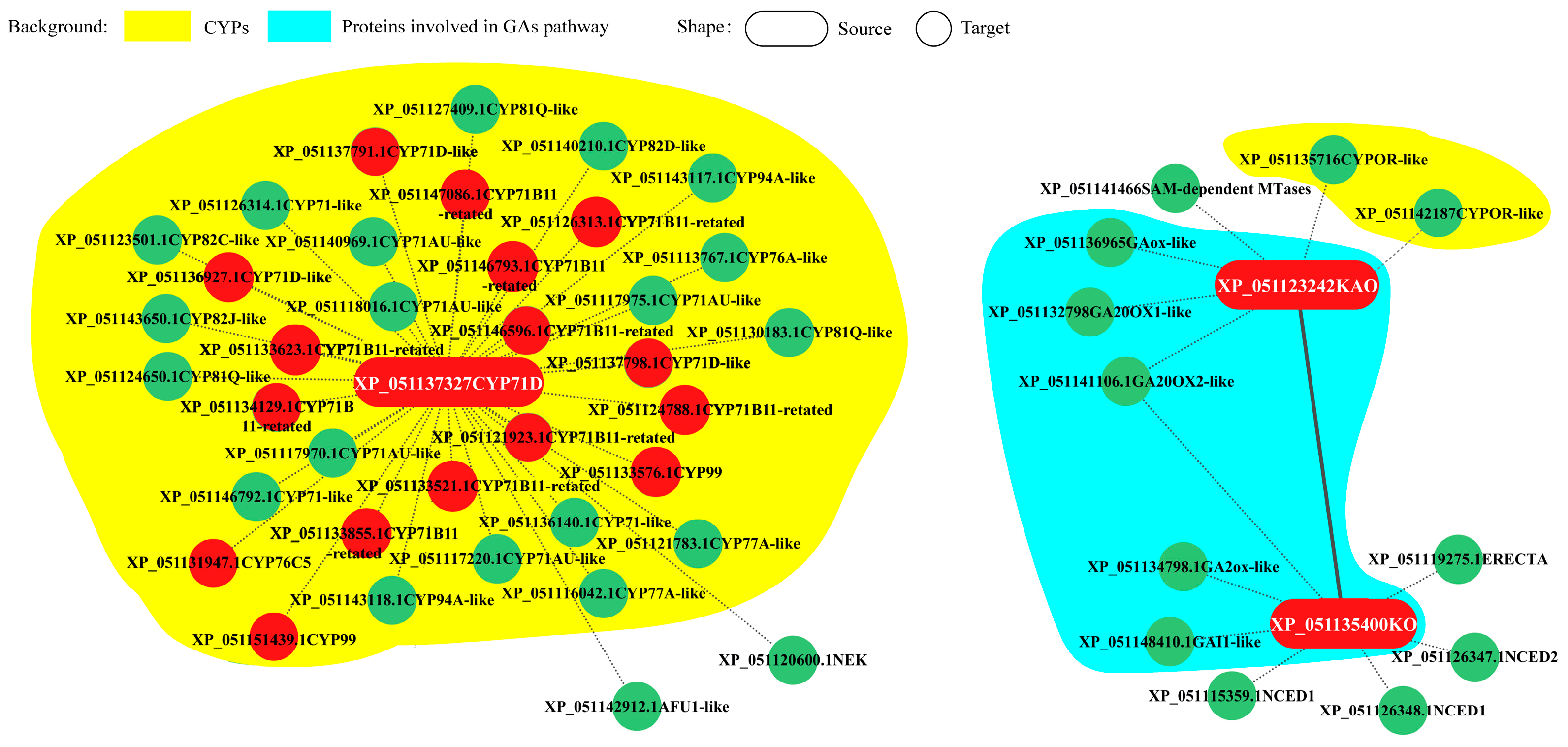
Andrographolide accumulated in the leaves (Figure 6A). Under LT conditions, the leaves of Andrographis paniculata turned red, with andrographolide content significantly decreasing (Figure 6B). To investigate the LT response of representative CYPs, Andrographis paniculata seedlings were subjected to different LT treatments: room temperature (green leaves, CK), mild stress (slight red leaves, SR), moderate stress (medium red leaves, MR), and severe stress (all red leaves, AR). Gene expression profiling revealed that three representative CYPs [protein IDs: XP_051123242 (KAO), XP_051135400 (KO), and XP_051137327 (CYP71D)] showed significant expression changes under LT conditions. These CYPs interacted with other differentially expressed genes (Figure 7). CYP71D had the most interacting proteins (37 proteins). Among them, 35 were CYPs (highlighted in yellow background), and 16 were diterpenoid pathway CYPs (marked with red circles). In the GA pathway, KAO and KO interacted with each other, and both also interacted with XP_051141106 (GA20OX2-like). Additionally, KAO and KO each independently interacted with two other GA pathway proteins (highlighted in blue background). KAO also interacted with two Cytochrome P450 oxidoreductases (CYPOR-like) (Figure 7).
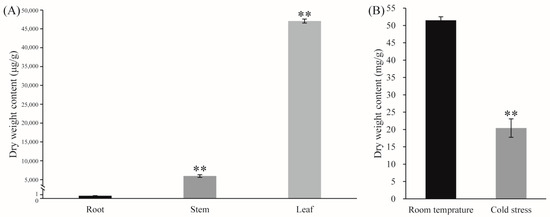
Figure 6.
(A) Tissue-specific accumulation of andrographolide; (B) andrographolide content in leaves under LT treatment Note: The data were represented as the averages of three independent biological experiments ± SD, and asterisks indicated a significant difference compared to the corresponding controls.
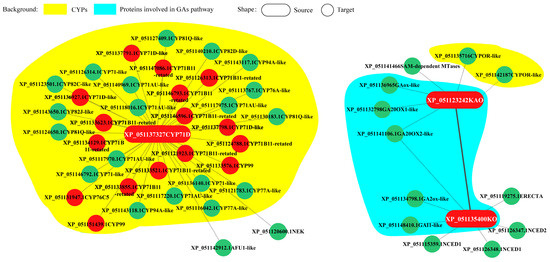
Figure 7.
Protein interaction of KAO, KO, and CYP71D Note: red—diterpenoid pathway CYPs; green—other interacting proteins.
2.7. LT Expression Profiling of KO, KAO, and CYP71D-Interacting Genes
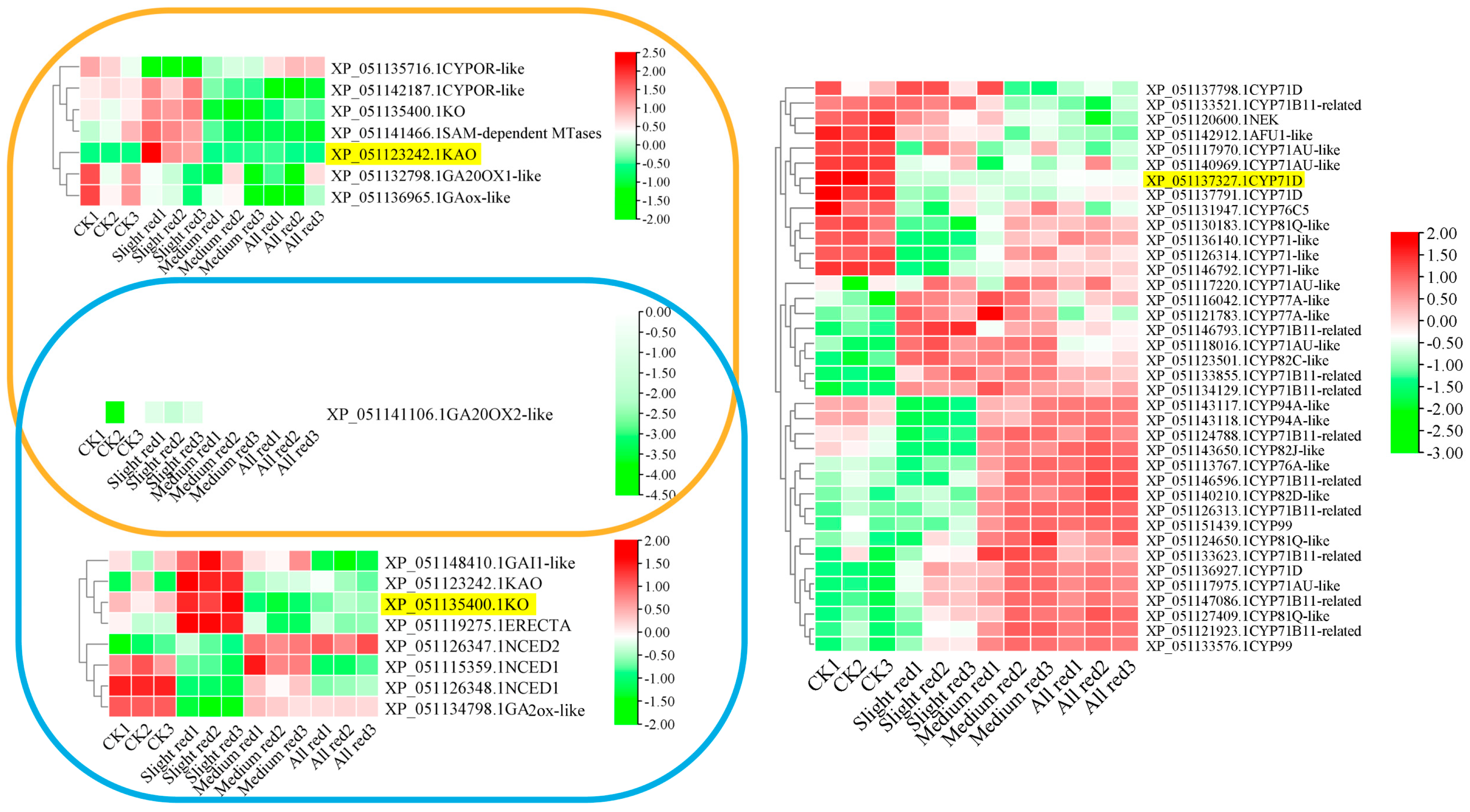
The accumulations of KAO and KO were significantly upregulated under mild LT stress but decreased under moderate to severe LT stress (Figure 8). The LT expression pattern of KAO was opposite to that of its interacting partners, GA20OX1-like and GAox-like. Similarly, KO exhibited a negative correlation with GA2ox-like, which interacted with it under the LT condition. The LT-related expression of GA20OX2-like did not show a clear trend relative to KAO and KO. The expression of CYP71D was significantly reduced under all LT treatments, indicating it was a negative regulator of the LT response. Conversely, transcripts of 25 CYPs interacting with CYP71D accumulated under varying degrees of LT stress. These CYPs served as positive response factors at the corresponding stages.
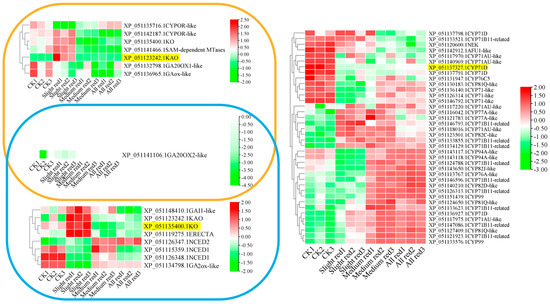
Figure 8.
LT−related expression profiles of KAO, KO, and CYP71D and transcripts of their interacting proteins. Note: yellow IDs—protein IDs of representative CYPs.
3. Discussion
CYPs play a vital role in diterpenoid metabolism in plants [22]. In this study, we identified and classified diterpenoid-modifying CYPs in Andrographis paniculata. A total of 345 CYPs were characterized across the genome, with 328 belonging to 42 known subfamilies. Seventeen CYPs were identified as unreported members of subfamilies specifically expanded in Andrographis paniculata. Further characterization of their catalytic functions is required. Subfamilies CYP736 and CYP749, absent in Arabidopsis but present in some other plants, were also found in Andrographis paniculata. CYP736A300 in oregano (Origanum vulgare; Lamiaceae) catalyzes the conversion of phenolic monoterpenes such as thymol and carvacrol [23]. CYP749A16 enhances cotton (Gossypium hirsutum) resistance to the sulfonylurea herbicide trifloxysulfuron-sodium [24]. The functions of CYP subfamilies across different plants show correlations [25]. These findings suggest that the CYP736 subfamily in Andrographis paniculata may play a role in phenolic monoterpene conversion. Additionally, its CYP749 subfamily could be associated with herbicide resistance.
Through phylogenetic analysis and conserved domain alignment, 65 candidate diterpenoid pathway CYPs (hereinafter called candidate CYPs) were identified in Andrographis paniculata. They were further analyzed at the protein, RNA, and DNA levels. Nearly 97% of the candidate CYPs had peptide chains around 500 aa, indicating structural and functional similarities. Previous research shows that GGPP is primarily synthesized via the plastidial MEP pathway [26]. In this study, 88% of the candidate CYPs were localized to the chloroplast, with 12% being membrane-associated. Most candidate CYPs (77%) were transmembrane proteins, with transmembrane domains near the N-terminus, consistent with previous findings [27]. Signal peptides, leader peptides, and signal sequences are responsible for protein sorting and translocation. Signal peptides facilitate co-translational translocation, sorting proteins into the endomembrane system. Leader peptides and signal sequences function in post-translational translocation. They are responsible for sorting proteins into specific organelles, including mitochondria, chloroplasts, and peroxisomes [28]. No reliable signal peptides were predicted for the candidate CYPs. This suggested that their transmembrane localization relied solely on post-translational leader peptides or signal sequences.
The motif distribution of candidate CYPs exhibited subfamily specificity. These CYPs were derived from the CYP71, CYP76, CYP88A, CYP701A, and CYP714 subfamilies. The CYP714 subfamily had the fewest motifs, while the CYP71 subfamily had the most, including CYP71B and CYP71D. The CYP71D subfamily has been reported to contribute to diterpenoid biosynthesis in tobacco (Nicotiana tabacum) [29], tomato (Solanum lycopersicum), and Euphorbiaceae plants [22]. Lamiaceae subfamilies CYP71AU [30], CYP71BE [31], CYP71D [32], and Lamiaceae-specific subfamilies CYP76AH [33], CYP76BK [34], and CYP76AK [35] all participate in diterpenoid modification. CYP76 is closely associated with the synthesis of specialized labdane-type diterpenoids [36]. In Andrographis paniculata (Acanthaceae), the function of candidate CYP XP_051150854 was annotated as CYP76AH, suggesting similar diterpenoid modification pathways across different families within Lamiales. It is essential to conduct an in-depth analysis of XP_051150854’s role in the labdane-type diterpenoid andrographolide metabolic process. In contrast, the CYP71AU and CYP71BE subfamilies, found in both Andrographis paniculata and Asteraceae plants, have fewer specific expansion members [22]. Their broader roles in diterpenoid biosynthesis remain uncertain. The CYP99A subfamily (clustered with CYP71B in this study) appears more inclined towards modifying Poaceae diterpenoids [37]. In addition, the oxidation of ent-kaurene is critical for hormone synthesis in terrestrial plants. Most members of the CYP701A subfamily are functioning as ent-kaurene oxidases [38]. The ring-contraction reaction catalyzed by the CYP88A subfamily is essential for GA synthesis in vascular plants [39]. Members of the CYP714 subfamily deactivate GAs by catalyzing their hydroxylation. This results in dwarf and stress-resistant phenotypes in plants overexpressing CYP714 [40].
The promoter cis-acting elements of the candidate CYPs were broadly connected with processes involving GAs, MeJA, auxin, ABA, and abiotic stress. This indicated that they were key factors in growth, development, and chemical defense. The extensive CYP71 subfamily not only modifies diterpenoids but also includes members involved in the modification of other terpenoids or flavonoids [41]. The promoters of two CYP71 subfamily candidate CYPs (protein IDs: XP_051133617 and XP_051146596) contained flavonoid biosynthesis elements. Gene structure analysis revealed that one candidate CYP (protein ID: XP_051152769) contained an exceptionally long intron exceeding 40,000 bp. It harbored essential elements influencing transcription and protein function. These included light-responsive, hormone (ABA and GA) signaling, bZIP and WRKY binding sites, and abiotic stress-responsive elements. Based on the arrangement of elements within the core and proximal promoters, 10 representative CYPs from each subfamily were selected. The spatial structures of proteins from members of three GA pathway subfamilies were conserved. Additionally, the structures of members in the CYP71B, CYP76AH, and CYP99 subfamilies within the CYP71 family were similar. But they differed from the structure of the CYP71D subfamily member XP_051137327. This reflects the considerable functional divergence within the CYP71D subfamily [32].
LTs restrict the yield of Andrographis paniculata. In this study, prolonged LT stress significantly reduced andrographolide levels in the leaves. LT may limit plant growth by affecting the activity of diterpenoid pathway CYPs. It could also reduce andrographolide synthase activity or accelerate andrographolide degradation. Among the 10 representative CYPs, three [protein IDs: XP_051123242 (CYP88A, KAO), XP_051135400 (CYP701A, KO), and XP_051137327 (CYP71D)] responded significantly to LT stress. Studies have shown that GAs serve as a crucial factor in enhancing plant LT tolerance [42]. Treatment with 0.5 mmol/L GAs effectively maintains cell wall integrity in tomato fruit, improving postharvest LT resistance [43]. KAO and KO in the GA biosynthesis pathway exhibited upregulation under mild LT stress. This was followed by downregulation during moderate and severe stress. In contrast, the expression of CYP71D was consistently downregulated under LT treatment. These results indicated that KAO and KO functioned as positive regulators in the early LT response. CYP71D acted as a negative response factor throughout all stages of LT stress. Meanwhile, under LT conditions, KAO and KO interacted with each other, other proteins in the GA pathway, and a GA20OX2-like protein. However, GA20OX2-like showed minimal accumulation under LT treatment, with no clear trend relative to KAO and KO. This suggested that KAO, KO, and GA20OX2-like might have formed a trimeric complex under LT stress, but the role of GA20OX2-like was likely non-essential. KAO and KO collaborated to assist Andrographis paniculata in responding to early LT stress within the GA biosynthesis pathway. However, both might have had only a modest effect on rescuing andrographolide accumulation. By the later stages of LT stress, their transcript accumulation declined. The plant’s biological functions could have been severely impaired, leading to a significant decrease in andrographolide content. CYP71D interacted with 37 proteins, 95% of which were CYPs, with 46% belonging to the diterpenoid pathway CYPs identified in this study. This suggested that CYP71D might function as an auxiliary protein for CYPs. LT stress consistently suppressed its transcriptional activity. It might have been directly or indirectly involved in the synthesis of andrographolide under LT stress. In conclusion, plant CYPs make critical contributions to the chemical diversity of diterpenoids [44]. The identification and characterization of these CYPs in Andrographis paniculata can improve its environmental adaptability and enhance the pharmaceutical applications of andrographolide. Refs. [45,46] are cited in Supplementary Materials.
4. Materials and Methods
4.1. Identification of Total CYPs in Andrographis Paniculata
The Andrographis paniculata genome (GenBank: GCA_009805555.1) and protein sequences were downloaded from NCBI (https://www.ncbi.nlm.nih.gov/). A Cytochrome P450 protein HMM model (PF00067) from Pfam 37.0 (http://pfam.xfam.org/, accessed on 5 December 2023) was used to identify CYPs via TBtools−II v2.067 (TBtools) Simple HMM search. Duplicate proteins were removed. Confirmed CYP sequences underwent NCBI Batch CD-search, NCBI BlastP (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome, accessed on 6 December 2023), and InterPro 98.0 database checks.
4.2. Construction of Phylogenetic Trees
CYPs from the diterpene pathway of terrestrial plants, excluding those belonging to gymnosperm-specific subfamilies, were selected as per reference [8], detailed in Table S4. Arabidopsis CYPs were sourced from TAIR (https://www.arabidopsis.org/, accessed on 4 January 2024). CYPs with missing sequences, duplicate sequences, and those duplicating terrestrial plant diterpene pathway CYPs were removed. Using MEGA 11, the sequences of Andrographis paniculata CYPs, terrestrial plant diterpene pathway CYPs, and Arabidopsis CYPs were aligned with MUSCLE. Two overly short sequences (Arabidopsis AT5G35920 and Andrographis paniculata XP_051128743) were removed. The remaining CYPs were submitted to PhyloSuite_v1.2.3 for Gblock analysis to extract conserved protein sequences. A phylogenetic tree of conserved sequences was constructed using TBtools’ One Step Build an ML Tree. The Andrographis paniculata CYPs that clustered with terrestrial plant diterpene pathway CYPs were selected as candidates. A phylogenetic tree of full-length protein sequences was constructed for these candidates and terrestrial plant diterpene pathway CYPs. The standalone phylogenetic analysis of candidate diterpenoid pathway CYPs in Andrographis paniculata also utilized full-length protein sequences. Final trees were refined using iTOL v6 (https://itol.embl.de/, accessed on 12 January 2024) and Adobe Illustrator 2020 (AI).
4.3. Identification of Conserved Domains in Andrographis Paniculata CYPs
Conserved domains of plant CYPs include the I-helix (A/G-G-X-E/D-T-T/S), K-helix [K-E-T-L-R (E and R conserved) and P-E-R-F (P and R conserved)], and FXXGXRXCXG. FXXGXRXCXG is a characteristic signature for plant CYPs. The Andrographis paniculata CYP protein sequences were submitted to WebLogo (https://weblogo.berkeley.edu/logo.cgi, accessed on 3 February 2024). The analysis showed that all CYPs possessed the I-helix and K-helix domains. But, some lacked FXXGXRXCXG. This was possibly due to the poor genome sequencing or assembly quality of Andrographis paniculata. Using DNAMAN v9, the protein sequences of Andrographis paniculata CYPs were aligned, and 22 CYPs that lacked a complete FXXGXRXCXG were removed. The remaining full-length sequences were resubmitted to WebLogo. The generated images were enhanced using AI.
4.4. Analysis of Gene Structure
The full-length protein sequences of candidate CYPs were aligned using MUSCLE. They were submitted to TBtools One Step Build an ML Tree, saving the Newick data. Motifs were analyzed on MEME (https://meme-suite.org/meme/tools/meme, accessed on 10 February 2024) with classic mode. The upstream 2500 bp from the start codon was extracted as a promoter. The promoters were submitted to PlantCare (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 16 February 2024) to screen for cis-acting elements. Gene lengths, positions of exons, introns, and untranslated regions were extracted. The Newick data, cis-acting element data, and gene component data were submitted to TBtools Gene Structure View (Advanced). AI was used to plot the gene structure diagrams. Functional elements of the unusually long intron were analyzed using PLACE (https://www.dna.affrc.go.jp/PLACE/?action=newplace, accessed on 2 May 2024) [21].
4.5. Analysis of Protein Characteristics
Predictive analyses included molecular mass (Expasy, https://www.expasy.org/, accessed on 13 May 2024), subcellular localization (WoLF PSORT, https://wolfpsort.hgc.jp/, accessed on 13 May 2024), transmembrane domains (TMHMM-2.0, https://services.healthtech.dtu.dk/services/TMHMM-2.0/, accessed on 17 May 2024), signal peptides (SignalP 5.0, https://services.healthtech.dtu.dk/services/SignalP-5.0/, accessed on 17 May 2024), 2D structure (PSIPRED, http://bioinf.cs.ucl.ac.uk/psipred/), and 3D structure (SWISS-MODEL, https://swissmodel.expasy.org/, accessed on 17 May 2024). Protein sequences were submitted to the STRING database (https://cn.string-db.org/, accessed on 17 May 2024). The interaction networks were constructed using Cytoscape. Interactions that had a combined score greater than 500 were retained. The image was annotated with AI.
4.6. Quantification of Andrographolide Content
The seeds of Andrographis paniculata, which are already widely cultivated, were sourced from Raoping County, Chaozhou City, Guangdong Province. They were cultivated at the Guangdong Provincial Key Laboratory of Crop Genetic Improvement, Crop Research Institute, Guangdong Academy of Agricultural Sciences.
4.6.1. Growth Conditions
The seeds were sterilized with 75% ethanol and 10% sodium hypochlorite, then sown in sterile peat soil and incubated at 28 °C with long-day conditions (16 h light/8 h dark) and 75% humidity. The plants were grown in pots, with Hoagland nutrient solution added every 3–5 days to maintain soil moisture. After 60 days, the seedlings were about 20 cm tall and had four pairs of fully expanded leaves, excluding the cotyledons. The roots, stems, and third leaves from the top were harvested for andrographolide content analysis. Additional seedlings were subjected to LT treatment.
4.6.2. LT Treatment Conditions
The seedlings were maintained at 20 °C for 8 weeks. LT damage to the leaves was assessed by observing reddening. The third leaves from the top were collected for andrographolide analysis at 0 weeks (CK, green leaves) and 8 weeks (AR, all red leaves) of LT treatment. Three independent biological replicates were used for each condition, each from at least 2 seedlings.
4.6.3. Detection of Andrographolide Using Liquid Chromatography–Tandem Mass Spectrometry
The andrographolide content was measured using liquid chromatography (Shim-pack UFLC SHIMADZU CBM30A, Shimadzu Corporation, Kyoto, Japan) with tandem mass spectrometry (Applied Biosystems 4500 QTRAP, Thermo Fisher Scientific, Waltham, Massachusetts, United States). The andrographolide standard has a CAS number of 5508-58-7 [47].
Chromatography was performed on a Waters ACQUITY UPLC HSS T3 C18 column (1.8 µm, 2.1 mm × 100 mm). The mobile phase included ultrapure water with 0.04% acetic acid (Phase A) and acetonitrile with 0.04% acetic acid (Phase B). The gradient was as follows: 0 min A/B 95:5, 10 min A/B 5:95, 11 min A/B 5:95, 11.1 min A/B 95:5, and 14 min A/B 95:5. The flow rate was 0.35 mL/min, with a column temperature of 40 °C and an injection volume of 3 μL.
Mass spectrometry settings were the following: ESI ion source, 550 °C temperature, 5500 V ion source voltage, 35 psi curtain gas, CAD -2, and optimized DP and CE.
4.6.4. Statistical Analysis
The mean and standard deviation (SD) of the three replicates were calculated. A Student’s t-test was conducted using SPSS 19.0 to determine significant differences at p < 0.05 (*) or p < 0.01 (**).
4.7. Gene Expression Analysis under LT Treatment
The third leaves from the top were collected at 0 weeks (CK), 2 weeks [mild stress, slight red leaves, SR], 4 weeks [moderate stress, medium red leaves, MR], and 8 weeks (AR) of LT treatment. The collected leaves were rapidly frozen in liquid nitrogen and stored at −80 °C. Three independent biological replicates were taken for each treatment, with each replicate consisting of leaves from at least two seedlings.
The total RNA extraction and transcriptome sequencing were conducted by Guangzhou Gene Denovo Biotechnology Co., Ltd. The FPKM values of the target genes in CK and treatment groups were log2-transformed (Table S5). They were submitted to TBtools HeatMap for generating heat maps of gene expression levels. The sequence data reported in this study were archived in the Sequence Read Archive (SRA) with the accession number PRJNA1167344.
The images were assembled with AI.
5. Conclusions
We systematically identified and classified 345 CYPs in Andrographis paniculata. Of these, 328 belonged to 42 known subfamilies, while 17 were possibly unique to Andrographis paniculata. Subfamilies CYP736 and CYP749, absent in Arabidopsis but present in some other plants, were also identified. Phylogenetic analysis and domain alignment revealed 65 candidate CYPs involved in diterpenoid pathways. These candidates were mostly transmembrane proteins localized to the chloroplast and had similar molecular sizes. Their motif distribution showed subfamily specificity. Promoter analysis indicated that these candidates were key factors in growth, development, and chemical defense. Based on the characteristics of cis-acting element arrangement and subfamily classification, 10 representative candidate CYPs were selected. The expression patterns under LT stress revealed that KAO (protein ID: XP_051123242) and KO (protein ID: XP_051135400) acted as positive regulators in the early LT response. CYP71D (protein ID: XP_051137327) functioned as a negative response factor throughout LT stress. CYP71D interacted with 37 proteins, 95% of which were CYPs, including 46% from the diterpenoid pathway. KAO and KO were found to interact with each other and with other proteins in the GA pathway, suggesting a coordinated role in response to LT stress. These findings enhance our understanding of the functional diversity of CYPs in Andrographis paniculata, particularly their roles in diterpenoid metabolism and LT responses. This knowledge provides a foundation for future studies aimed at improving the environmental adaptability of Andrographis paniculata and enhancing the pharmaceutical applications of andrographolide. The candidates supply direction for research on andrographolide and GAs in molecular biology, synthetic biology, and related fields.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms251910741/s1. Refs. [45,46] are cited in Supplementary Materials file.
Author Contributions
Conceptualization, M.S. and J.W.; Methodology, M.S., J.L. and S.X.; Software, M.S. and S.X.; Validation, M.S. and J.L.; Formal analysis, M.S.; Investigation, J.L. and Y.G.; Resources, J.L. and Y.G.; Data curation, M.S.; Writing—original draft, M.S.; Writing—review & editing, M.S.; Visualization, M.S. and J.L.; Supervision, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Guangdong Basic and Applied Basic Research Foundation (2023A1515010426), the Guangzhou Science and Technology Plan Project (2023A04J0777), the Crops Research Institute, Guangdong Academy of Agricultural Sciences, Key Laboratory of Crops Genetic Improvement of Guangdong Province (202308).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sun, M.-Y.; Xu, S.-Q.; Zhang, W.-T.; Gu, Y.; Mei, Y.; Li, J.-Y.; Zhou, F.; Wang, J.-H. Advances in scientific research on Andrographis paniculata. Chin. Agric. Sci. Bull. 2022, 38, 155–164. [Google Scholar]
- Lim, J.C.W.; Chan, T.K.; Ng, D.S.W.; Sagineedu, S.R.; Wong, W.S.F. Andrographolide and its analogues: Versatile bioactive molecules for combating inflammation and cancer. Clin. Exp. Pharmacol. Physiol. 2012, 39, 300–310. [Google Scholar] [CrossRef]
- Sun, M.-Y.; Xu, S.-Q.; Li, J.-Y.; Gu, Y.; Mei, Y.; Zhang, W.-T.; Zhou, F.; Wang, J.-H. Transcriptome analysis and discovery of genes related to andrographolide synthesis of Andrographis paniculata under UV-C treatment. J. South. Agric. 2022, 53, 618–627. [Google Scholar]
- Tan, H.-T.; Gong, D.-Y.; Li, B. Advances in the biosynthetic pathway of dendrobine and terpenoid CYP450 enzymes. J. Chongqing Technol. Bus. Univ. (Nat. Sci. Ed.) 2022, 39, 1–13. [Google Scholar]
- Li, D. Cloning and Function Analysis of CYP82Ds Genes from Camellia sinensis. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2021. [Google Scholar]
- Nelson, D.; Werck-Reichhart, D. A P450-centric view of plant evolution. Plant J. 2011, 66, 194–211. [Google Scholar] [CrossRef]
- Zheng, X.-Y.; Li, P.; Lu, X. Research advances in cytochrome P450-catalysed pharmaceutical terpenoid biosynthesis in plants. J. Exp. Bot. 2019, 70, 4619–4630. [Google Scholar] [CrossRef]
- Bathe, U.; Tissier, A. Cytochrome P450 enzymes: A driving force of plant diterpene diversity. Phytochemistry 2019, 161, 149–162. [Google Scholar] [CrossRef]
- Mitu, S.; Ogbourne, S.; Klein, A.; Tran, T.; Cummins, S. The CYP450 multigene family of Fontainea and insights into diterpenoid synthesis. BMC Plant Biol. 2021, 21, 191. [Google Scholar] [CrossRef]
- Hu, J.-D.; Qiu, S.; Wang, F.-Y.; Li, Q.; Xiang, C.-L.; Di, P.; Wu, Z.-D.; Jiang, R.; Li, J.-X.; Zeng, Z.; et al. Functional divergence of CYP76AKs shapes the chemodiversity of abietane-type diterpenoids in genus Salvia. Nat. Commun. 2023, 14, 4696. [Google Scholar] [CrossRef]
- Mizobuchi, R.; Sato, H.; Fukuoka, S.; Tanabata, T.; Tsushima, S.; Imbe, T.; Yano, M. Mapping a quantitative trait locus for resistance to bacterial grain rot in rice. Rice 2013, 6, 13. [Google Scholar] [CrossRef]
- King, A.J.; Brown, G.D.; Gilday, A.D.; Larson, T.R.; Graham, I.A. Production of bioactive diterpenoids in the Euphorbiaceae depends on evolutionarily conserved gene clusters. Plant Cell 2014, 26, 3286–3298. [Google Scholar] [CrossRef]
- Kuang, X.-J.; Sun, S.-J.; Wei, J.-H.; Li, Y.; Sun, C. Iso-Seq analysis of the Taxus cuspidata transcriptome reveals the complexity of Taxol biosynthesis. BMC Plant Biol. 2019, 19, 210. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Wu, Q.-Y.; Ge, Y.; Huang, Z.-Y.; Hong, R.; Li, A.-T.; Xu, J.-H.; Yu, H.-L. Hydroxylases involved in terpenoid biosynthesis: A review. Bioresour. Bioprocess. 2023, 10, 39. [Google Scholar] [CrossRef]
- Eilon, S.; Peter, H.; Tai-Ping, S. Highlights in gibberellin research: A tale of the dwarf and the slender. Plant Physiol. 2024, 195, 111. [Google Scholar]
- He, J.; Chen, Q.-W.; Xin, P.-Y.; Yuan, J.; Ma, Y.-H.; Wang, X.-M.; Xu, M.-M.; Chu, J.-F.; Peters, R.J.; Wang, G.-D. CYP72A enzymes catalyse 13-hydrolyzation of gibberellins. Nat. Plants. 2019, 5, 1057–1065. [Google Scholar] [CrossRef]
- Wang, J.-W.; Liu, Z.-Y.; She, H.-B.; Xu, Z.-S.; Zhang, H.-L.; Fang, Z.-W.; Qian, W. Genome-wide identification and characterization of U-Box gene family members and analysis of their expression patterns in Phaseolus vulgaris L. under cold stress. Int. J. Mol. Sci. 2024, 25, 7968. [Google Scholar] [CrossRef]
- Kumar, M.S.; Babu, P.R.; Rao, K.V.; Reddy, V.D. Organization and classification of cytochrome P450 genes in castor (Ricinus communis L.). Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2014, 84, 131–143. [Google Scholar] [CrossRef]
- Zhang, W.; Li, H.-X.; Li, Q.-H.; Wang, Z.-W.; Zeng, W.-W.; Yin, H.; Qi, K.-J.; Zou, Y.; Hu, J.; Huang, B.-S.; et al. Genome-wide identification, comparative analysis and functional roles in flavonoid biosynthesis of cytochrome P450 superfamily in pear (Pyrus spp.). BMC Genomic Data 2023, 24, 58. [Google Scholar] [CrossRef]
- Liu, X.-L.; Cao, F.-L.; Cai, J.-F.; Wang, H.-L. The molecular cloning and expression analysis of a CYP71 gene in Ginkgo biloba L. Not. Bot. Horti Agrobo. Cluj-Napoca 2016, 44, 77–84. [Google Scholar] [CrossRef]
- Xia, T.-F.; Yang, Y.-P.; Zheng, H.-Y.; Han, X.-Y.; Jin, H.-B.; Xiong, Z.-J.; Qian, W.-Q.; Xia, L.-Q.; Ji, X.; Li, G.-W.; et al. Efficient expression and function of a receptor-like kinase in wheat powdery mildew defense require an intron located MYB binding site. Plant Biotechnol. J. 2021, 19, 897–909. [Google Scholar] [CrossRef]
- Wang, Z.-B.; Nelson, D.R.; Zhang, J.; Wan, X.-Y.; Peters, R.J. Plant (di)terpenoid evolution: From pigments to hormones and beyond. Nat. Prod. Rep. 2023, 40, 452–469. [Google Scholar] [CrossRef]
- Mai, T.D.; Kim, H.M.; Park, S.Y.; Ma, S.H.; Do, J.H.; Choi, W.; Jang, H.M.; Hwang, H.B.; Song, E.G.; Shim, J.S.; et al. Metabolism of phenolic compounds catalyzed by Tomato CYP736A61. Enzyme Microb. Technol. 2024, 176, 110425. [Google Scholar] [CrossRef]
- Thyssen, G.N.; Naoumkina, M.; McCarty, J.C.; Jenkins, J.N.; Florane, C.; Li, P.; Fang, D.D. The P450 gene CYP749A16 is required for tolerance to the sulfonylurea herbicide trifloxysulfuron sodium in cotton (Gossypium hirsutum L.). BMC Plant Biol. 2018, 18, 186. [Google Scholar] [CrossRef]
- Nelson, D.R.; Ming, R.; Alam, M.; Schuler, M.A. Comparison of cytochrome P450 genes from six plant genomes. Trop. Plant Biol. 2008, 1, 216–235. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Sun, W.-T.; Zhang, X.-Z.; Wan, S.-T.; Wang, R.-W.; Li, C. Regulation on oxidation selectivity for β-amyrin by Class Ⅱ cytochrome P450 enzymes. Synth. Biol. J. 2021, 2, 804. [Google Scholar]
- Owji, H.; Nezafat, N.; Negahdaripour, M.; Hajiebrahimi, A.; Ghasemi, Y. A comprehensive review of signal peptides: Structure, roles, and applications. Eur. J. Cell Biol. 2018, 97, 422–441. [Google Scholar] [CrossRef]
- Wang, E.; Wang, R.; DeParasis, J.; Loughrin, J.H.; Gan, S.; Wagner, G.J. Suppression of a P450 hydroxylase gene in plant trichome glands enhances natural-product-based aphid resistance. Nat. Biotechnol. 2001, 19, 371–374. [Google Scholar] [CrossRef]
- Karunanithi, P.S.; Dhanota, P.; Addison, J.B.; Tong, S.; Fiehn, O.; Zerbe, P. Functional characterization of the cytochrome P450 monooxygenase CYP71AU87 indicates a role in marrubiin biosynthesis in the medicinal plant Marrubium vulgare. BMC Plant Biol. 2019, 19, 114. [Google Scholar] [CrossRef]
- Ignea, C.; Athanasakoglou, A.; Ioannou, E.; Georgantea, P.; Trikka, F.A.; Loupassaki, S.; Roussis, V.; Makris, A.M.; Kampranis, S.C. Carnosic acid biosynthesis elucidated by a synthetic biology platform. Proc. Natl. Acad. Sci. USA 2016, 113, 3681–3686. [Google Scholar] [CrossRef]
- Ma, Y.; Cui, G.; Chen, T.; Ma, X.; Wang, R.; Jin, B.; Yang, J.; Kang, L.; Tang, J.; Lai, C.-J.-S.; et al. Expansion within the CYP71D subfamily drives the heterocyclization of tanshinones synthesis in Salvia miltiorrhiza. Nat. Commun. 2021, 12, 685. [Google Scholar] [CrossRef] [PubMed]
- Pateraki, I.; Andersen-Ranberg, J.; Jensen, N.B.; Wubshet, S.G.; Heskes, A.M.; Forman, V.; Hallström, B.; Hamberger, B.; Motawia, M.S.; Olsen, C.E.; et al. Total biosynthesis of the cyclic AMP booster forskolin from Coleus forskohlii. Elife 2017, 6, e23001. [Google Scholar] [CrossRef] [PubMed]
- Heskes, A.M.; Sundram, T.C.M.; Boughton, B.A.; Jensen, N.B.; Hansen, N.L.; Crocoll, C.; Cozzi, F.; Rasmussen, S.; Hamberger, B.; Hamberger, B.; et al. Biosynthesis of bioactive diterpenoids in the medicinal plant Vitex agnus-castus. Plant J. 2018, 93, 943–958. [Google Scholar] [CrossRef]
- Guo, J.; Ma, X.-H.; Cai, Y.; Ma, Y.; Zhan, Z.-L.; Zhou, Y.-J.J.; Liu, W.J.; Guan, M.-X.; Yang, J.; Cui, G.-H.; et al. Cytochrome P450 promiscuity leads to a bifurcating biosynthetic pathway for tanshinones. New Phytol. 2016, 210, 525–534. [Google Scholar] [CrossRef]
- Chen, Z.-X. Prokaryotic Expression, Purification and Crystallography Studies on CYP76AH3 from Salvia miltiorrhiza and CYP725A4 from Taxus wallichiana. Ph.D. Thesis, Peking University Health Science Center, Beijing, China, 2022. [Google Scholar]
- Schmelz, E.A.; Huffaker, A.; Sims, J.W.; Christensen, S.A.; Lu, X.; Okada, K.; Peters, R.J. Biosynthesis, elicitation and roles of monocot terpenoid phytoalexins. New Phytol. 2014, 79, 659–678. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Weckwerth, P.R.; Poretsky, E.; Murphy, K.M.; Sims, J.; Saldivar, E.; Christensen, S.A.; Char, S.N.; Yang, B.; Tong, A.D.; et al. Genetic elucidation of complex biochemical traits mediating maize innate immunity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Peters, R.J. Gibberellin Phytohormone Metabolism. Isoprenoid Synthesis in Plants and Microorganisms: New Concepts and Experimental Approaches; Springer: Berlin/Heidelberg, Germany, 2013; pp. 233–249. [Google Scholar]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 1947–1952. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S. Triterpene structural diversification by plant cytochrome P450 enzymes. Front. Plant Sci. 2017, 8, 1886. [Google Scholar] [CrossRef]
- Huang, X.-F.; Qu, T.-T.; Ding, Y.; Lei, T.-T.; Zhang, J.; Tang, X.-M. Chilling resistance of gibberellin-deficiency tomato fruits. Food Ferment. Ind. 2020, 46, 8. [Google Scholar]
- Zhu, Z.; Ding, Y.; Zhao, J.-H.; Nie, Y.; Zhang, Y.; Sheng, J.-P.; Tang, X.-M. Effects of postharvest gibberellic acid treatment on chilling tolerance in cold-stored tomato (Solanum lycopersicum L.) Fruit. Food Bioprocess Technol. 2016, 9, 1202–1209. [Google Scholar] [CrossRef]
- Sun, W.; Leng, L.; Yin, Q.-G.; Xu, M.-M.; Huang, M.-K.; Xu, Z.-C.; Zhang, Y.-J.; Yao, H.; Wang, C.-X.; Xiong, C.; et al. The genome of the medicinal plant Andrographis paniculata provides insight into the biosynthesis of the bioactive diterpenoid neoandrographolide. Plant J. 2018, 97, 841–857. [Google Scholar] [CrossRef] [PubMed]
- Zi, J.-C.; Peters, R.J. Characterization of CYP76AH4 clarifies phenolic diterpenoid biosynthesis in the Lamiaceae. Org. Biomol. Chem. 2013, 11, 7650–7652. [Google Scholar] [CrossRef] [PubMed]
- Esposito, S.; Cardi, T.; Campanelli, G.; Sestili, S.; JoséDíez, M.; Soler, S.; Prohens, J.; Tripodi, P. ddRAD sequencing-based genotyping for population structure analysis in cultivated tomato provides new insights into the genomic diversity of Mediterranean’da serbo’type long shelf-life germplasm. Hortic. Res. 2020, 7, 134. [Google Scholar] [CrossRef] [PubMed]
- CAS 5508-58-7. Available online: https://www.merckmillipore.com/INTL/en/product/Andrographolide-CAS-5508-58-7-Calbiochem,EMD_BIO-172060?ReferrerURL=https%3A%2F%2Fwww.google.com%2F (accessed on 1 October 2024).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).