Abstract
Dwarf bamboo (Fargesia denudata) is a crucial food source for the giant pandas. With its shallow root system and rapid growth, dwarf bamboo is highly sensitive to drought stress and nitrogen deposition, both major concerns of global climate change affecting plant growth and rhizosphere environments. However, few reports address the response mechanisms of the dwarf bamboo rhizosphere environment to these two factors. Therefore, this study investigated the effects of drought stress and nitrogen deposition on the physicochemical properties and microbial community composition of the arrow bamboo rhizosphere soil, using metagenomic sequencing to analyze functional genes involved in carbon and nitrogen cycles. Both drought stress and nitrogen deposition significantly altered the soil nutrient content, but their combination had no significant impact on these indicators. Nitrogen deposition increased the relative abundance of the microbial functional gene nrfA, while decreasing the abundances of nirK, nosZ, norB, and nifH. Drought stress inhibited the functional genes of key microbial enzymes involved in starch and sucrose metabolism, but promoted those involved in galactose metabolism, inositol phosphate metabolism, and hemicellulose degradation. NO3−-N showed the highest correlation with N-cycling functional genes (p < 0.01). Total C and total N had the greatest impact on the relative abundance of key enzyme functional genes involved in carbon degradation. This research provides theoretical and technical references for the sustainable management and conservation of dwarf bamboo forests in giant panda habitats under global climate change.
1. Introduction
In the context of global change, climate change factors do not operate in isolation; they occur simultaneously, leading to complex coupling effects. Studies have revealed significant variations in the impact on soil physicochemical properties, functions and microbial biodiversity when multiple climate change factors are combined [1,2]. This results in synergistic, antagonistic, or additive effects among these factors. Water availability and nitrogen, crucial factors for plant growth under climate change, are profoundly influenced. During drought stress, nitrogen deposition significantly alleviates the adverse impacts of drought stress on plants by increasing leaf PSII activity and leaf photosynthesis [3,4,5]. Additionally, anthropogenic nitrogen addition severely affects the ecosystem functioning of soil microbial communities in semi-arid temperate grasslands [6,7]. Despite these findings, there is a substantial gap in our understanding of the coupled effects of drought stress and nitrogen deposition on plants and their rhizospheric environment. Consequently, conducting further in-depth studies in this area is imperative.
Meanwhile, the intensification, frequency, and impact range of extreme weather and climate events, such as drought, are escalating due to global climate change [8]. Drought not only disrupts the coordinated development of the social economy but also degrades the ecological environment, leading to ecological security concerns [9]. Recent environmental pressures and global warming have caused arid regions to expand into humid areas in southern China, threatening nearly half of the total land area. Drought stress is thus emerging as a significant and urgent challenge for sustainable development in China [10]. Additionally, global atmospheric nitrogen deposition has seen a significant annual increase. With rapid economic development, China has become the world’s leading producer and emitter of reactive nitrogen, making it the primary nitrogen deposition region in Asia [11]. Consequently, the ecological impacts of both drought and nitrogen deposition, along with their intricate interconnections, have become hotspots in the global climate change discourse. Many researchers have delved into this topic, aiming to comprehend the complexities of these interactions [4,6,12]. Despite the progress, there is still much to explore to enhance our understanding of these intertwined phenomena and address the challenges they present.
The rhizosphere, located within a few millimeters from the root surface, differs from the surrounding soil in its physical, chemical, and biological properties. It serves as the primary gateway for nutrients or harmful substances to enter the plant [13] and acts as a vital nexus connecting plants, soil, and microorganisms. This microenvironment plays a crucial role in material cycling and capacity flow within ecosystems [14,15]. Rhizosphere changes are effective adaptations by plants to extreme environments, making it a significant domain for soil-root microbial interactions. Particularly in the context of global climate change, the rhizosphere has emerged as a focal point for research in plant physiology, soil science, microbiology, and ecology [16,17,18].
As a national treasure and the primary food source for giant pandas, dwarf bamboo (Fargesia denudata) mainly grows in the southwest alpine areas of China, offering essential ecological functions such as water conservation, soil stability, and nutrient balance. As a semi-woody plant with shallow roots, dwarf bamboo has a high water demand and is easily susceptible to drought conditions [19,20]. Drought can result in reduced bamboo shoots, shedding of bamboo leaves, and even premature flowering and death of dwarf bamboo [21]. Furthermore, due to its rapid growth, dwarf bamboo is also remarkably sensitive to nitrogen deposition [22]. Current research has explored the physiological responses to drought stress, nitrogen deposition, and their combined effects on bamboo [20]. However, there are limited studies on the response mechanisms of dwarf bamboo to drought stress and nitrogen deposition from the perspective of the rhizosphere environment. This research is vital in providing a scientific foundation and technical guidance for the management and protection of dwarf bamboo, which is essential for the habitat of giant pandas under future climate change.
This study focuses on the rhizosphere soil of dwarf bamboo, investigating the effects of drought stress and nitrogen deposition on the physicochemical properties, microbial community structure, and functional genes related to carbon and nitrogen cycling in the rhizosphere soil, utilizing techniques such as metagenomics. The aim is to further explore the mutual relationship between microbial and physicochemical properties in the rhizosphere soil of dwarf bamboo under the influence of drought stress and nitrogen deposition. The objective is to reveal the ecological adaptability mechanisms of the rhizosphere microenvironment of dwarf bamboo to drought stress and nitrogen deposition. This research provides a theoretical foundation and technical reference for the sustainable management and conservation of dwarf bamboo forests in the giant panda habitat under future climate change.
2. Results
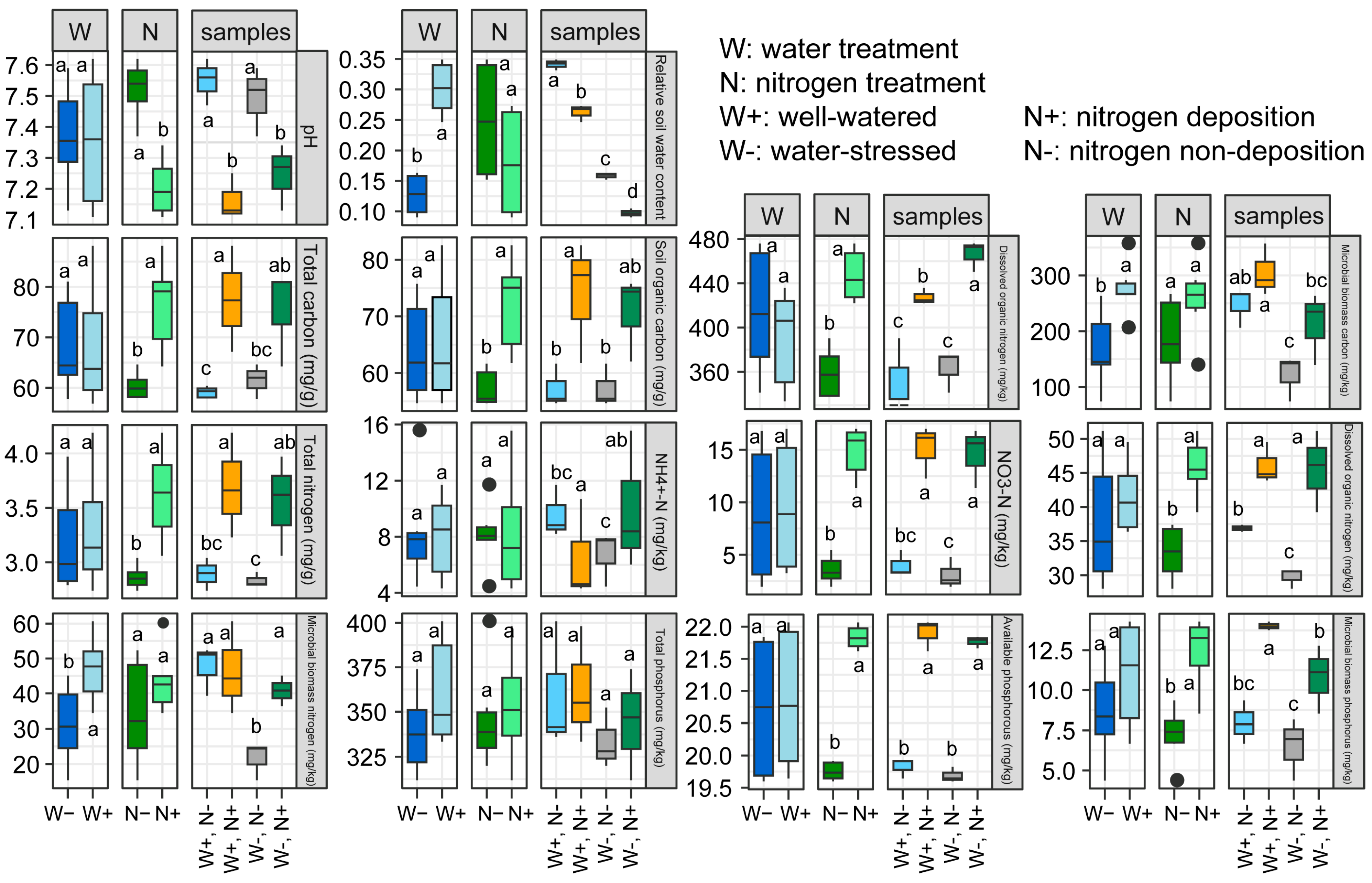
2.1. Effects of Nitrogen (N) Deposition and Drought Stress on Soil Physicochemical Index
Soil nutrients are essential nutrients provided by the soil for plant growth. The dynamics of rhizosphere soil nutrients not only reflect the relationship between root-soil nutrient supply and demand but also indicate the influence of environmental conditions, exhibiting a high sensitivity to environmental changes [23]. Under both nitrogen deposition and non-deposition conditions, drought stress did not significantly affect the pH of rhizosphere soil (Figure 1). However, under well-watered and drought-stress conditions, nitrogen deposition significantly reduced soil pH. Regarding soil carbon, drought stress did not significantly alter the levels of total nitrogen (TN), soil organic carbon (SOC), and dissolved organic carbon (DOC) in the rhizosphere soil. Nevertheless, it notably decreased the microbial biomass carbon (MBC) content, while nitrogen deposition notably increased the concentrations of total carbon (TC), SOC, and DOC in the rhizosphere soil. Concerning soil nitrogen, drought stress showed no significant impact on the levels of total phosphorus (TP), ammonium nitrogen (NH4+-N), nitrate nitrogen (NO3−-N), and dissolved organic nitrogen (DON) in the rhizosphere soil, except for a significant reduction in microbial biomass nitrogen (MBN) content. As for soil phosphorus, nitrogen deposition markedly augmented the concentrations of available phosphorus (AP) and microbial biomass phosphorus (MBP) in the rhizosphere soil. However, no significant effects were observed on all phosphorus indices in the rhizosphere soil of dwarf bamboo under different watering conditions (Figure 1).
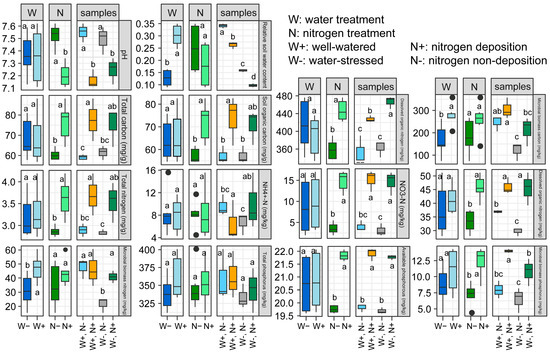
Figure 1.
Effects of drought stress and nitrogen deposition on the rhizosphere soil physical-chemical properties of Fargesia denudata under non-nitrogen (N), deposition (−N), and N deposition (+N) with and without drought stress. Different letters indicate the significant differences between means, lower case indicates p < 0.05.
2.2. Effects of Nitrogen (N) Deposition and Drought Stress on Microbial Communities and Functions
After sequencing and read-processing, 3,348,077 effective reads were obtained with a total sequence length of 1,448,960,064 and an average sequence length of 431.49 bp (Table 1). Alpha diversity analysis showed that drought stress and nitrogen deposition did not significantly affect the Chao1 and ACE (p > 0.05), indicating that these treatments had no significant impact on the richness of the microbial community under rhizosphere soil (Table 2). However, both drought stress and nitrogen deposition significantly influenced the microbial community diversity, Shannon, and Simpson index, in rhizosphere soil (p < 0.05) (Table 2).

Table 1.
Metagenome sequencing data for different treatments.
Table 2.
Diversity index in rhizosphere soil of Fargesia under different treatments.
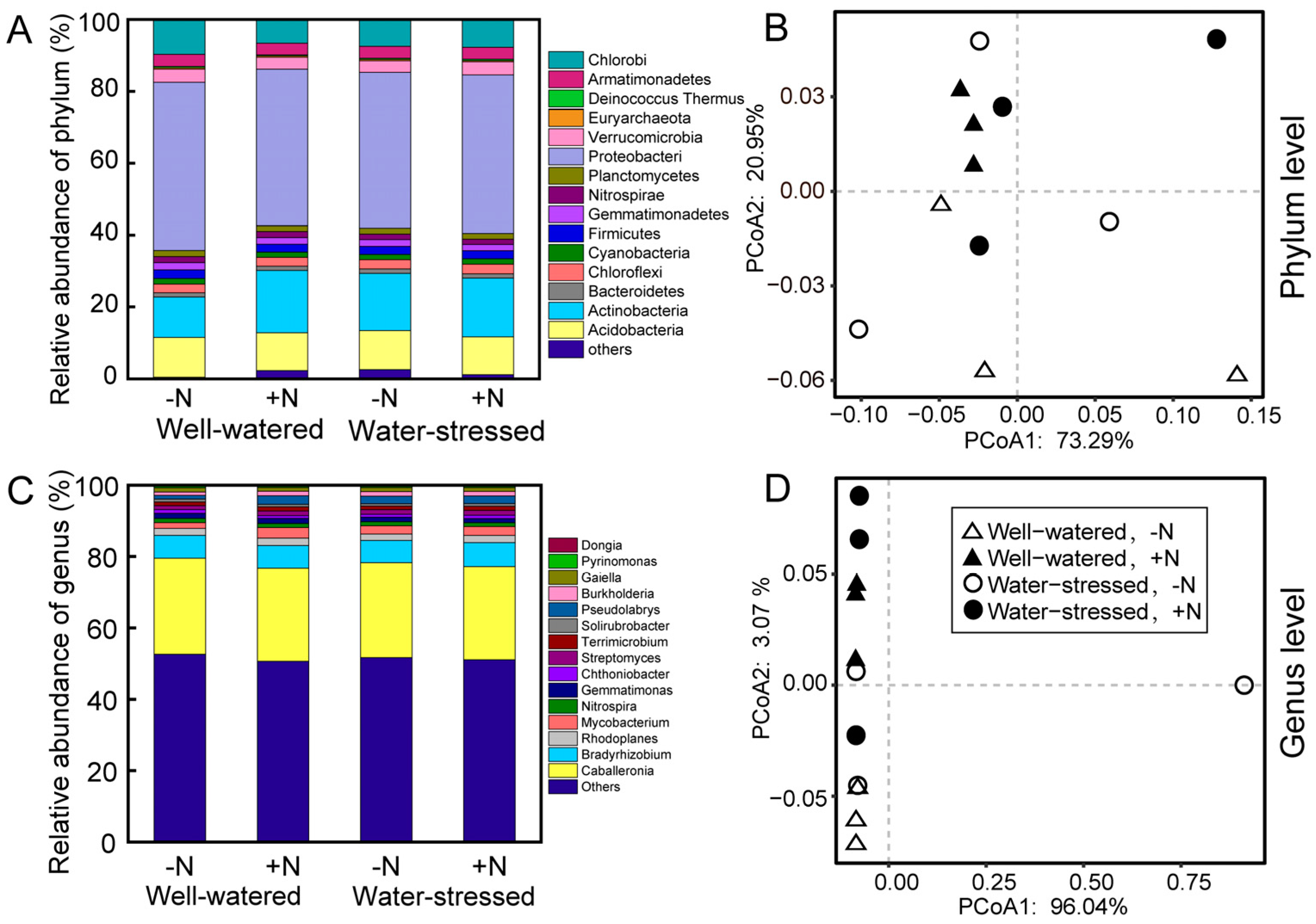
The distribution and abundance of bacteria in soils at the phylum and genus levels were analyzed (Figure 2). On the phylum level, Proteobacteria, Actinobacteria, and Acidobacteria had the highest relative abundance among all samples (Figure 2A). Drought stress had no significant effect on the relative abundance of these phyla. While nitrogen deposition only significantly increased the relative abundance of Actinobacteria, increasing by 16.05% (Table S1). The analysis of PCoA and PERMANOVA also showed that both N deposition and drought stress, as well as their combination, have no effects on the microbial community structure in the rhizosphere soil at the phylum level (Table S2).
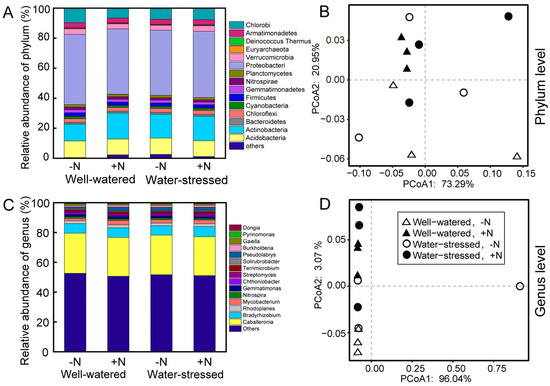
Figure 2.
Relative abundance (A,C) and PCoA (B,D) of species at phylum and genus levels of the microbial community in rhizosphere soil of Fargesia denudata under different treatments.
On the genus level, Caballeronia, Bradyrhizobium, Rhodoplanes, and Mycobacterium were the highest predominant genus among all samples (Figure 2C). Under normal water conditions, nitrogen deposition significantly increased the relative abundance of the genus Bradyrhizobium in the rhizosphere soil of dwarf bamboo, whereas nitrogen deposition increased the relative abundance of the genus Rhizobium under drought stress (Table S3). In the absence of N deposition, drought stress significantly increased the relative abundance of the genus Bradyrhizobium, whereas drought stress significantly increased the relative abundance of the genus Rhizobium under N deposition (Table S3).
Based on the KEGG pathway level-3 enrichment analysis, the relative abundance of microbial functional genes involved in nitrogen and carbon cycling was obtained (Figure S1). Most genes in N and C cycling were enriched in groups without N deposition, while the related genes decreased after N deposition. At the nitrogen metabolism level, we further analyzed the relative abundance changes of microbial functional genes involved in assimilatory nitrate reduction (nirA), dissimilatory nitrate reduction (nrfA), nitrification (amoA), denitrification (narG, napA, nirK, nirS, norB and nosZ), and nitrogen fixation (nifH) in the nitrogen cycle (Table 3). The relative abundance of functional genes related to N cycle, nirK, nosZ, nifH and norB, was significantly decreased after N deposition, while nrfA showed an upward trend in N deposition treatments. However, drought stress and nitrogen deposition did not significantly affect the relative abundance of functional genes, such as nirA, nirS, narG, napA, and amoA (Table 3).
Table 3.
Relative abundance of main microbial functional genes involved in the nitrogen cycle in rhizosphere soil of Fargesia denudata under different treatments.
At the carbon metabolism level, drought stress significantly affected the relative abundance of functional genes involved in inositol phosphate metabolism, whereas nitrogen deposition had a highly significant impact on the relative abundance of functional genes involved in C5-branched dibasic acid metabolism, citrate cycle, fructose, and mannose metabolism, glycolysis/gluconeogenesis, glyoxylate, and dicarboxylate metabolism, pentose and glucuronate interconversions, propanoate metabolism, pyruvate metabolism, carbon fixation in photosynthetic organisms, and carbon fixation pathways in prokaryotes (Table 4). The relative abundance of functional genes involved in galactose metabolism and starch and sucrose metabolism was markedly changed under the coupled nitrogen–drought stress treatment (Table 4).
Table 4.
Relative abundance of functional genes involved in carbon metabolism in rhizosphere soil of Fargesia denudata under different treatments.
At the carbon-degrading enzymes level, drought stress significantly affected the relative abundance of functional genes of hemicellulose-degrading enzymes (p < 0.05) (Table 5). Nitrogen deposition had a highly significant impact on the relative abundance of functional genes of cellobiose transporters, hemicellulose degradation, and sugar transporters (p < 0.05). The coupling effect of drought stress and nitrogen deposition did not significantly affect these functional genes involved in carbon degradation (p > 0.05) (Table 5).
Table 5.
Relative abundance of functional genes involved in carbon degradation in rhizosphere soil of Fargesia denudata under different treatments.
2.3. Correlation between N Cycling-Related Microorganisms and Soil Physicochemical Index
There was a correlation observed between the soil physicochemical properties and the relative abundance of microbial functional genes (Figure 3). These genes are involved in assimilatory nitrate reduction (nirA), dissimilatory nitrate reduction (nrfA), denitrification pathways (narG, napA, nirK, nirS, norB, nosZ), and nitrogen fixation pathways (nifH). However, no significant correlation was found with microbial functional genes involved in amoA (Figure 3). Among these genes, those involved in nifH exhibited the strongest correlation with the soil’s physicochemical properties (p < 0.01). Conversely, the correlation between the physicochemical properties and functional genes involved in nirA was the weakest, only demonstrating a significant negative correlation with NO3−-N (p < 0.05). Additionally, NO3−-N showed the highest correlation with functional genes associated with N-cycling (p < 0.01). Conversely, NH4+-N, MBC, and MBN exhibited no significant correlation with functional genes involved in N-cycling (p > 0.05).
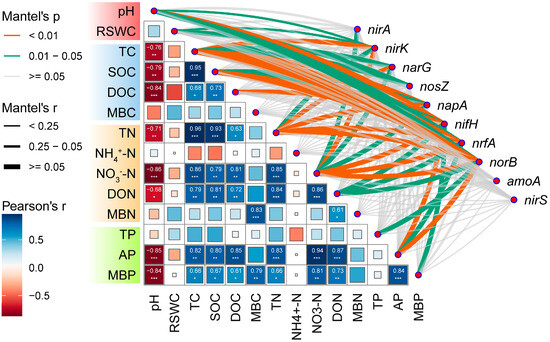
Figure 3.
Correlation analysis of main microbial functional genes involved in nitrogen cycle with soil physical and chemical properties in rhizosphere soil of Fargesia denudata. *** p < 0.001; ** p < 0.01; * p < 0.05.
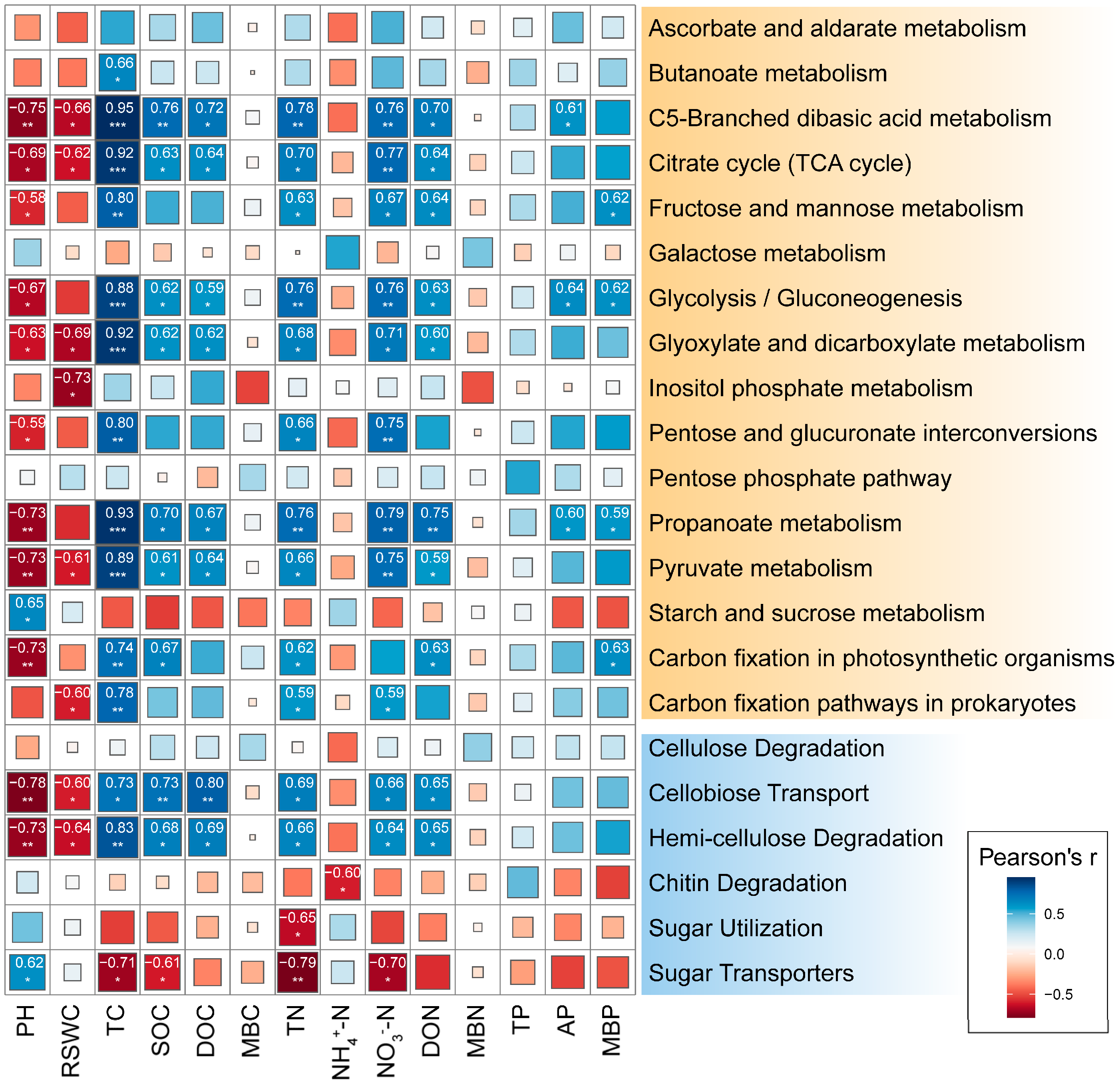
2.4. Correlation between C Cycling-Related Microorganisms and Soil Physicochemical Index
The relative abundance of microbial functional genes involved in C5-branched dibasic acid metabolism, citrate cycle, glycolysis/gluconeogenesis, glyoxylate, and dicarboxylate metabolism, propanoate metabolism, pyruvate metabolism showed significant positive correlations with TC, SOC, DOC, TN, NO3−-N, and DON while showing negative correlations with pH and RSWC (Figure 4). However, MBC, MBN, and TP exhibited no significant correlation with the relative abundance of functional genes involved in C cycling. The relative abundance of the gene encoding for cellobiose transporter and hemicellulose degradation was significantly positively correlated with soil TC, SOC, DOC, TN, NO3−-N, and DON while showing a significant negative correlation with soil pH and RSWC. Cellulose degradation was not significantly correlated with soil physicochemical index. Chitin degradation and sugar utilization had a significant negative correlation with NH4+-N and TN, respectively. Sugar transporters were negatively correlated with four soil indices, including TC, SOC, TN, and NO3−-N. Overall, among the soil physicochemical indicators, TC and TN had the greatest impact on the relative abundance of key enzyme functional genes involved in carbon degradation.
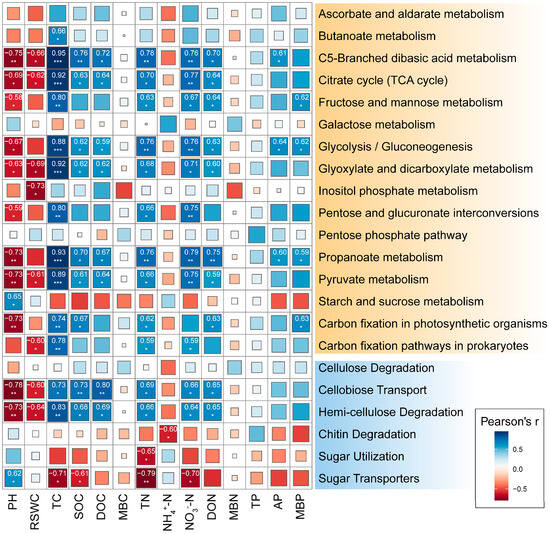
Figure 4.
Correlation analysis of the main microbial functional genes involved in carbon cycle with soil physical and chemical properties in rhizosphere soil of Fargesia denudata. *** p < 0.001; ** p < 0.01; * p < 0.05.
3. Discussion
3.1. Response of Soil Physicochemical Index to Drought Stress and Nitrogen Deposition
Research on rhizosphere soil nutrients typically focuses on essential elements such as soil carbon, nitrogen, and phosphorus [24]. Drought stress and nitrogen deposition are significant environmental factors driving changes in rhizosphere soil carbon, nitrogen, and phosphorus [25]. Drought can alter soil nutrient dynamics, leading to reduced effectiveness [26]. Our study demonstrated that drought stress decreases MBC, MBN, and MBP in rhizosphere soil. This decline may be due to increased nutrient uptake by drought-stressed dwarf bamboo or reduced nutrient transformations. While appropriate N deposition can enhance N mineralization and organic matter decomposition, excessive N can disrupt soil nutrient balance and lead to acidification [27,28]. Our findings revealed that N deposition significantly increased TC, SOC, DOC, TN, NO3−-N, DON, AP, and MBP levels in dwarf bamboo rhizosphere soil, promoting nutrient accumulation (Figure 1) [27]. Additionally, N deposition may enhance available N content and photosynthetic assimilation, improving drought resistance [29]. These results suggest that N deposition enhances rhizosphere soil nutrient content in drought-stressed dwarf bamboo, augmenting its adaptation to dry conditions.
3.2. Response of Soil Microbial Community Structure and Diversity to Drought Stress and Nitrogen Deposition
Soil microbial community structure and diversity serve as important indicators of soil quality, being highly sensitive to external environmental conditions [30]. Studies have found that soil properties, vegetation types, and climatic conditions significantly influence soil microbial diversity [31,32]. Here, metagenomic sequencing technology was used to investigate the microbial community structure and diversity in the rhizosphere of dwarf bamboo under drought stress and nitrogen deposition. Results revealed that Proteobacteria, Actinobacteria, and Acidobacteria were the predominant phyla (Figure 2), which is consistent with the previous findings [33,34]. Drought stress and nitrogen deposition significantly increased the relative abundance of the genus Bradyrhizobium in the rhizosphere soil of arrow bamboo (Table S3). Bradyrhizobium, belonging to the phylum Proteobacteria, is known for nitrogen fixation and phosphorus solubilization, benefiting plant growth [35,36]. Thus, nitrogen deposition and its coupling with drought stress enhance nitrogen fixation and phosphorus solubilization abilities of rhizosphere soil microbes, promoting nutrient absorption and utilization by arrow bamboo, consequently enhancing its growth. Previous studies have found that water and nitrogen significantly alter soil microbial community structure, but their coupling mostly exhibits antagonistic effects [37,38]. Similarly, our study also demonstrates that both drought stress and nitrogen deposition affected soil microbial community diversity in the dwarf bamboo rhizosphere, with their combination also showing antagonistic effects. This means that the combined effect of both stressors did not amplify changes in microbial community structure as might be expected, but rather dampened or neutralized each other’s influence in certain microbial populations. This could be due to the fact that nitrogen deposition may alleviate some of the limitations caused by drought stress, or that drought conditions might restrict the effectiveness of nitrogen’s influence on microbial activities. These antagonistic interactions have important implications for understanding how complex environmental factors simultaneously influence soil microbial dynamics and plant-microbe interactions, particularly in changing climate conditions.
3.3. The Effect of N Deposition on Microbial Functional Genes for Soil C-N Cycling
As major climate factors of global climate change, drought stress, and nitrogen deposition can alter soil carbon and nitrogen transformation processes [39]. Many scholars have conducted research on the effects of drought stress on microbial functional genes involved in nitrogen cycling [40,41,42]. However, these studies typically focused on individual environmental factors. With the exacerbation of global climate change, multiple environmental factors often occur simultaneously or coexist. Therefore, we investigated the impacts of drought and nitrogen deposition on soil microbial functional genes involved in nitrogen cycling in the rhizosphere soil of arrow bamboo. We found that drought stress alone or coupled with nitrogen deposition had little effect on most microbial functional genes involved in nitrogen cycling, while nitrogen deposition significantly influenced many of these genes (Table 3). Specifically, nitrogen deposition markedly increased the relative abundance of the functional gene nrfA involved in dissimilatory nitrate reduction, while decreasing the relative abundance of nirK, nosZ, and norB genes involved in denitrification (Table 3). This indicates that nitrogen deposition suppresses denitrification, reducing nitrogen loss and accelerating the conversion of NO2− to NH4+ in the rhizosphere soil [43]. Consequently, it was shown to enhance nitrogen utilization by arrow bamboo. The observed increase in nrfA and decrease in nirK genes are notable, as they highlight specific shifts in nitrogen cycling pathways under nitrogen deposition. nrfA encodes for a key enzyme in the dissimilatory nitrate reduction to ammonium (DNRA) pathway, while nirK is involved in the denitrification process, converting nitrite (NO2−) to nitric oxide (NO) [44]. The increase in nrfA suggests that nitrogen deposition enhances DNRA activity, leading to a greater retention of nitrogen in the form of ammonium (NH4+), which is more readily available for plant uptake. In contrast, the decrease in nirK indicates a suppression of denitrification, which reduces nitrogen loss from the system in the form of nitrogen gases (e.g., N2, N2O).
These shifts in microbial nitrogen cycling pathways can have significant ecological implications for the overall nitrogen budget of the ecosystem. The increase in DNRA activity promotes nitrogen conservation within the soil, as ammonium is retained rather than lost to the atmosphere through denitrification [45]. This can lead to a more efficient nitrogen cycle, where plants such as arrow bamboo benefit from enhanced nitrogen availability. However, the reduction in denitrification could alter the balance of nitrogen gases emitted from the soil, potentially decreasing the release of greenhouse gases such as nitrous oxide (N2O), a potent greenhouse gas [46]. Furthermore, the shift from denitrification to DNRA may influence the microbial community composition and soil chemical properties over time [47]. With more ammonium being retained in the soil, this could lead to increased ammonium toxicity or soil acidification if nitrogen deposition continues over the long term, potentially affecting soil health and plant-microbe interactions. However, nitrogen deposition had no significant effect on the relative abundance of functional genes involved in nitrification (Table 3), suggesting that nitrogen deposition may not consistently promote nitrogen cycling in the rhizosphere soil of arrow bamboo. This could be due to soil acidification and reduced microbial activity upon reaching certain nitrogen levels, leading to inhibition of nitrogen cycling [48,49]. Nitrogen deposition application reduced the activity and diversity of nitrogen-fixing microbes, inhibiting their nitrogen-fixing function [50,51,52]. Similarly, our study found that nitrogen deposition significantly decreased the relative abundance of the nitrogen fixation gene (nifH), suggesting that nitrogen deposition may reduce the activity of nitrogen-fixing microbes.
In the process of carbon cycling, both drought stress and nitrogen deposition can affect the soil microbes or their major functional genes involved in carbon cycling. Li, Xie [53] and Kuerban, Cong [54] found that nitrogen and water treatments altered the functionality of soil microbes, leading to changes in microbial metabolic activity and utilization of carbon sources. Some studies have also suggested that nitrogen deposition may stimulate the expression of microbial functional genes involved in carbon degradation, thereby increasing the decomposition of starch, chitin, and pectin in the soil [55,56,57]. This study also found that drought stress inhibited the relative abundance of functional genes of carbon cycling (cellulose-degrading enzymes), while nitrogen deposition significantly promoted the relative abundance of most functional genes involved in carbon cycling (cellulose transporter and cellulose-degrading enzymes) (Table 5). However, the coupling of these functional genes showed no significant effect. This indicated that drought stress inhibits the decomposition rate of organic matter, while nitrogen deposition promotes organic matter decomposition, which is consistent with the findings in the grasslands under nitrogen deposition conditions [7,58]. On a broader scale, these changes in carbon metabolism can significantly influence soil nutrient cycling. The increased microbial activity related to cellulose degradation and lignin decomposition under nitrogen deposition can enhance organic matter turnover, releasing carbon compounds that contribute to soil organic matter formation. This process improves carbon sequestration and helps maintain soil structure and fertility. Additionally, enhanced carbon cycling indirectly affects other nutrient cycles, particularly nitrogen and phosphorus, as microbes release and recycle these nutrients during organic matter decomposition. Thus, nitrogen deposition not only increases carbon degradation but also promotes nutrient availability for plant uptake, ultimately supporting ecosystem productivity. However, the suppression of microbial activity under drought conditions limits these processes, reducing nutrient cycling efficiency. However, there is still a limited and incomplete understanding of the effects of drought and nitrogen deposition on functional genes involved in carbon cycling in forest rhizosphere soil, necessitating further research in this area.
The combined effects of drought and nitrogen deposition were not as significant as anticipated. One possible explanation is that the microbial community may exhibit compensatory mechanisms, where nitrogen deposition might alleviate some of the limitations imposed by drought stress, or drought could limit the impact of nitrogen deposition. Additionally, the duration of the stress treatments in this study was relatively short so a longer duration might lead to more pronounced effects. Finally, soil characteristics such as moisture retention and pH could have played a role in modulating microbial responses, thus limiting the observable effects of the combined stresses. Future studies should consider longer stress durations and more variable environmental conditions to gain a better understanding of the interactions between these factors.
3.4. The Effect of Soil Physicochemical Index on Microbial Communities
Drought stress and nitrogen deposition have led to changes in soil physicochemical properties and nutrient distribution, which affect the diversity, structure, and function of soil microbial communities [59,60]. Therefore, we conducted the correlation analysis between the soil physicochemical properties and its microbial community to reveal the response of soil physicochemical properties and microbial communities to drought stress and nitrogen deposition. Previous studies have demonstrated that soil pH and nutrients are significant driving factors in the variation of soil microbial community structure [61,62]. Similarly, in this study a significant correlation was found between pH and nutrients and the functional genes involved in carbon and nitrogen cycling, highlighting soil pH and nutrients as crucial factors affecting the structure and function of soil microbial communities in Fargesia (Figure 3 and Figure 4). The concentration of NO3−-N is the main factor influencing microbial functional genes involved in nitrogen cycling, whereas total carbon, total nitrogen, and nitrate nitrogen content are the main factors influencing microbial functional genes involved in carbon cycling. We speculate that nitrogen deposition might primarily affect the microbial communities involved in soil carbon and nitrogen cycling by increasing the content of total carbon, total nitrogen, and nitrate nitrogen, while drought stress had no significant effect on the soil nutrient content, thus exerting a relatively smaller impact on the microbial communities involved in soil carbon and nitrogen cycling.
However, the combined effects of drought and nitrogen deposition were not as significant as anticipated. One possible explanation is that the microbial community may exhibit compensatory mechanisms, where nitrogen deposition might alleviate some of the limitations imposed by drought stress, or drought could limit the impact of nitrogen deposition. Additionally, the duration of the stress treatments in this study was relatively short, and a longer duration might lead to more pronounced effects. Finally, soil characteristics such as moisture retention and pH could have played a role in modulating microbial responses, limiting the observable effects of the combined stresses. Future studies should consider longer stress durations and more variable environmental conditions to gain a better understanding of the interactions between these factors.
4. Materials and Methods
4.1. Research Area and Experiment Design
The experiment site was located at the Maoxian Ecological Research Station, Chinese Academy of Sciences (103°53′58″ E, 31°41′07″ N, 1820 ma.s.l.) in southwestern China. The healthy and uniform dwarf bamboo plants (2 years old) were collected from the nursery at Wangliang National Nature Reserve (103°55′ E, 32°49′ N) and transplanted into 50 L plastic pots with 70 kg of homogenized soil. One standard plant with 4–5 ramets was cultured in each pot. All plants were grown in a semi-controlled solar greenhouse with ambient conditions of 8–33 °C, relative humidity of 40–85%, and watered every 3 days.
Seven months after transplantation, the experimental design was applied with two factors (water regime and N deposition). The N deposition treatments were performed, including without N deposition [0 g N m−2 year−1 (−N): no addition of NH4NO3 solutions to each pot] and with N deposition [10 g N m−2 year−1 (+N), addition of 200 mL NH4NO3 solutions (6.18 mM N) to each pot weekly]. The amount of N addition level was determined according to the atmospheric N accumulation rate in the study area (3.9 g N m−2 year−1) [63], this increased level was used to simulate elevated nitrogen deposition levels commonly observed in rapidly developing industrial regions. After two months of N deposition treatments, different water treatments were performed. Well-watered (W+): soil was maintained at 80–85% relative soil water content (RSWC), simulating conditions of adequate water availability. Water-stressed (W−): soil was maintained at 30–35% RSWC, simulating drought conditions by withholding water for 30 days. The chosen RSWC levels for well-watered and water-stressed treatments were based on prior studies that have identified these thresholds as critical for plant physiological responses under drought conditions in similar environments [64,65]. RSWC in each treatment was controlled by the weight method, where soil moisture was adjusted accordingly to maintain target levels throughout the experiment. Each treatment had three replications with nine standard plants per replication. At the end of the experiment, the rhizosphere soil was collected from the plant and placed into plastic bags for metagenome analysis and the physiochemistry properties. Air-dried soil samples were filtered through a 1-mm sieve.
4.2. Soil Physiochemistry Properties
To characterize the soil under different treatments, we analyzed 13 soil physical-chemical properties. Soil pH was measured in soil suspension with distilled water (1:2.5 w/v) using a pH meter (FE20, Mettler Toledo, Shanghai, China). Total carbon (TC) and total nitrogen (TN) of the soil were measured using a Vario MACRO cube Elemental analyzer (Elementar, Hanau, Germany). Dissolved organic carbon (DOC), soil organic carbon (SOC), and dissolved organic nitrogen (DON) were measured using a total carbon analyzer (Multi N/C 2100, Jena Analytic, Langewiesen, Germany). The concentrations of NH4+-N and NO3−-N were determined using SEAL AA3 continuous flow analysis (SEAL Analytical, Norderstedt, Germany). The total phosphorus (TP) of the soil was determined using H2SO4-HClO4 digestion. Available phosphorous (AP) was determined by the molybdenum-antimony-scandium colorimetry method. Soil microbial biomass phosphorus (MBP), microbial biomass carbon (MBC), and microbial biomass nitrogen (MBN) were measured using the chloroform fumigation extraction method.
4.3. Metagenome Sequencing
One gram of the soil sample was used for microbial DNA extraction (Power Soil DNA Isolation Kit, Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The extracted DNA was detected by 2% agarose gel electrophoresis, and the purity of DNA was quantitatively analyzed with NanoDrop 2000 (Thermo Scientific, Wilmington, NC, USA). To construct the paired-end library, 300 bp DNA fragments were randomly sheared using Covaris M220 (Covaris Inc., Woburn, MA, USA) and end-repair, dA-tailing, and adapter ligation procedures were performed using NEBNext Rapid DNA-Seq (Bioo Scientific, Austin, TX, USA). Sequencing was performed on an Illumina Hiseq Xten (Illumina Inc., San Diego, CA, USA) at Majorbio Biotech Co., Ltd. (Shanghai, China).
Illumina raw reads were cleaned and trimmed using fastp v0.23.2 to remove low-quality reads [66]. Clean data were then assembled to obtain contigs based on the succinct de Bruijn graph method by MEGAHIT software (version 1.2.9), and then the contigs were used to predicate the open reading frames (ORFs) using MATAGENE [67]. Then, the predicted ORFs (>100 bp) were translated to protein sequences and clustered at 95% sequence identity and 90% coverage using CD-HIT v4.6 (http://www.bioinformatics.org/cd-hit/ (accessed on 10 September 2019)), and the longest genes of every cluster were selected as the representative sequence to construct a nonredundant gene catalog. The high quality of each sample read was blasted with a nonredundant gene set at 95% sequence identity to obtain the abundance of genes in each sample using SOAPaligner software (version 2.21). Gene taxonomic classification and functional annotation were performed against the NCBI-NR database, COG (Clusters of orthologous and Groups), and KEGG (Kyoto Encyclopedia of Genes and Genomes) databases, respectively, using a BLASTP search (BLAST Version 2.2.28+) [68].
4.4. Statistical Analysis
All data were analyzed by one-way analysis of variance (ANOVA), and multiple comparisons among treatments were performed using the least significant difference (LSD) method. Then, two-way ANOVA was used to evaluate the effects of water regime, nitrogen deposition, and their interaction on all dependent variables. Principal Coordinates Analysis (PCoA) based on Euclidean distance was used to assess the microbial community structure in the rhizosphere soil. Finally, Pearson correlation analysis was applied to evaluate the correlation between rhizosphere soil physicochemical properties and microbial community. All data analyses were performed using Excel 2016, SPSS 24, and R 2.5.6 software. Graphs were generated using Origin 9.0 software and packages such as vegan and ggplot2 in R 2.5.6 were utilized. The raw data from metagenome sequencing was deposited in NCBI under the accession number PRJNA1120641.
5. Conclusions
This study highlights the significant impacts of drought stress and nitrogen deposition on the rhizosphere environment of dwarf bamboo (Fargesia denudata), a vital food source for giant pandas. This research demonstrates that both factors alter soil nutrient content and microbial community composition, although their combined effect is insignificant. Specifically, nitrogen deposition increases the relative abundance of the microbial functional gene nrfA and decreases the abundances of nirK, nosZ, norB, and nifH, indicating changes in nitrogen cycling processes. Drought stress influences microbial functional genes differently, inhibiting those related to starch and sucrose metabolism while promoting genes involved in galactose metabolism, inositol phosphate metabolism, and hemicellulose degradation. Additionally, this study identifies NO3−-N as highly correlated with N-cycling functional genes and finds that total C and total N significantly impact the relative abundance of key enzyme functional genes involved in carbon degradation. These findings enhance our understanding of the response of microbial functional genes involved in the soil carbon and nitrogen cycles to drought stress and nitrogen deposition, allowing for better predictions of the structure and function of soil microbial communities under global change.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms251910790/s1.
Author Contributions
J.X.: Data curation, writing—original draft preparation, visualization, investigation; N.Z.: Formal analysis; J.L.: Data Curation; Y.Z.: Formal analysis; T.C.: Writing—reviewing and editing; Y.W.: Conceptualization, methodology, software, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
The Second Tibetan Plateau Scientific Expedition and Research Program (STEP), Grant: 2019QZKK0303; National Natural Science Foundation of China, Grant/Award Numbers: 31470621.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article and Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yang, S.; Zheng, Q.; Yuan, M.; Shi, Z.; Chiariello, N.R.; Docherty, K.M.; Dong, S.; Field, C.B.; Gu, Y.; Gutknecht, J. Long-term elevated CO2 shifts composition of soil microbial communities in a Californian annual grassland, reducing growth and N utilization potentials. Sci. Total Environ. 2019, 652, 1474–1481. [Google Scholar] [CrossRef]
- Rillig, M.C.; Ryo, M.; Lehmann, A.; Aguilar-Trigueros, C.A.; Buchert, S.; Wulf, A.; Iwasaki, A.; Roy, J.; Yang, G. The role of multiple global change factors in driving soil functions and microbial biodiversity. Science 2019, 366, 886–890. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Dong, S.; Li, S.; Xiao, J.; Han, Y.; Yang, M.; Zhang, J.; Gao, X.; Xu, Y.; Li, Y. Effects of simulated N deposition on photosynthesis and productivity of key plants from different functional groups of alpine meadow on Qinghai-Tibetan plateau. Environ. Pollut. 2019, 251, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tariq, A.; Zeng, F.; Graciano, C.; Zhang, B. Nitrogen application mitigates drought-induced metabolic changes in Alhagi sparsifolia seedlings by regulating nutrient and biomass allocation patterns. Plant Physiol. Biochem. 2020, 155, 828–841. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, J.; Liu, M.; Meng, Z.; Liu, K.; Sui, N. Nitrogen increases drought tolerance in maize seedlings. Funct. Plant Biol. 2019, 46, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Peguero, G.; Folch, E.; Liu, L.; Ogaya, R.; Penuelas, J. Divergent effects of drought and nitrogen deposition on microbial and arthropod soil communities in a Mediterranean forest. Eur. J. Soil Biol. 2021, 103, 103275. [Google Scholar] [CrossRef]
- Wang, Z.; Na, R.; Koziol, L.; Schellenberg, M.P.; Li, X.; Ta, N.; Jin, K.; Wang, H. Response of bacterial communities and plant-mediated soil processes to nitrogen deposition and precipitation in a desert steppe. Plant Soil 2020, 448, 277–297. [Google Scholar] [CrossRef]
- Ummenhofer, C.C.; Meehl, G.A. Extreme weather and climate events with ecological relevance: A review. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160135. [Google Scholar] [CrossRef]
- Han, L.; Zhang, Q.; Ma, P.; Jia, J.; Wang, J. The spatial distribution characteristics of a comprehensive drought risk index in southwestern China and underlying causes. Theor. Appl. Climatol. 2016, 124, 517–528. [Google Scholar] [CrossRef]
- Liu, F.; Xiao, X.; Qin, Y.; Yan, H.; Huang, J.; Wu, X.; Zhang, Y.; Zou, Z.; Doughty, R.B. Large spatial variation and stagnation of cropland gross primary production increases the challenges of sustainable grain production and food security in China. Sci. Total Environ. 2022, 811, 151408. [Google Scholar] [CrossRef]
- Cui, S.; Shi, Y.; Groffman, P.M.; Schlesinger, W.H.; Zhu, Y.-G. Centennial-scale analysis of the creation and fate of reactive nitrogen in China (1910–2010). Proc. Natl. Acad. Sci. USA 2013, 110, 2052–2057. [Google Scholar] [CrossRef] [PubMed]
- Aronson, E.L.; Goulden, M.L.; Allison, S.D. Greenhouse gas fluxes under drought and nitrogen addition in a Southern California grassland. Soil Biol. Biochem. 2019, 131, 19–27. [Google Scholar] [CrossRef]
- McNear, D., Jr. The Rhizosphere-Roots Soil and Everything in Between. Nat. Educ. Knowl. 2013, 4, 1. [Google Scholar]
- Liu, S.; He, F.; Kuzyakov, Y.; Xiao, H.; Hoang, D.T.T.; Pu, S.; Razavi, B.S. Nutrients in the rhizosphere: A meta-analysis of content, availability, and influencing factors. Sci. Total Environ. 2022, 826, 153908. [Google Scholar] [CrossRef]
- Lau, J.A.; Lennon, J.T. Rapid responses of soil microorganisms improve plant fitness in novel environments. Proc. Natl. Acad. Sci. USA 2012, 109, 14058–14062. [Google Scholar] [CrossRef]
- Kuppe, C.W.; Schnepf, A.; von Lieres, E.; Watt, M.; Postma, J.A. Rhizosphere models: Their concepts and application to plant-soil ecosystems. Plant Soil 2022, 474, 17–55. [Google Scholar] [CrossRef]
- Ling, W.; Ma, B.; Zhang, W. Rhizosphere microbiology: Toward a clean and healthy soil environment. Front. Microbiol. 2022, 13, 991356. [Google Scholar] [CrossRef]
- Suman, J.; Rakshit, A.; Ogireddy, S.D.; Singh, S.; Gupta, C.; Chandrakala, J. Microbiome as a key player in sustainable agriculture and human health. Front. Soil Sci. 2022, 2, 821589. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Y.; Pan, K.; Zhu, T.; Li, W.; Zhang, L. Carbon and nitrogen metabolism in leaves and roots of dwarf bamboo (Fargesia denudata Yi) subjected to drought for two consecutive years during sprouting period. J. Plant Growth Regul. 2014, 33, 243–255. [Google Scholar] [CrossRef]
- Wu, S.; Tian, J.; Ren, T.; Wang, Y. Osmotic adjustment and antioxidant system regulated by nitrogen deposition improve photosynthetic and growth performance and alleviate oxidative damage in dwarf bamboo under drought stress. Front. Plant Sci. 2022, 13, 819071. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Huang, J. Reseach into Bomboo Species as Giant Panda’s Main Diet and its Progress. World Bamboo Ratt. 2005, 3, 1–6. [Google Scholar]
- Zhou, X.; Wang, J.; Zhang, H.; Wang, J.; Zhang, Y. Effect of elevated CO2 and nitrogen deposition on leaf nutrient quality of Fargesia rufa Yi. Acta Ecol. Sin. 2012, 32, 7644–7653. [Google Scholar] [CrossRef]
- Pieters, A.J.; Paul, M.J.; Lawlor, D.W. Low sink demand limits photosynthesis under Pi deficiency. J. Exp. Bot. 2001, 52, 1083–1091. [Google Scholar] [CrossRef] [PubMed]
- Berhe, A.A.; Barnes, R.T.; Six, J.; Marín-Spiotta, E. Role of soil erosion in biogeochemical cycling of essential elements: Carbon, nitrogen, and phosphorus. Annu. Rev. Earth Planet. Sci. 2018, 46, 521–548. [Google Scholar] [CrossRef]
- Xu, N.; Guo, W.; Liu, J.; Du, N.; Wang, R. Increased nitrogen deposition alleviated the adverse effects of drought stress on Quercus variabilis and Quercus mongolica seedlings. Acta Physiol. Plant. 2015, 37, 107. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Lu, H.; Shao, Y.; Liu, S.; Fu, S. Drought promotes soil phosphorus transformation and reduces phosphorus bioavailability in a temperate forest. Sci. Total Environ. 2020, 732, 139295. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, J.; Sayer, E.J.; Lambers, H.; Liu, Z.; Lu, X.; Li, Y.; Li, Y.; Li, H.; Wang, F. Nitrogen deposition enhances soil organic carbon and microbial residual carbon in a tropical forest. Plant Soil 2023, 484, 217–235. [Google Scholar] [CrossRef]
- Wang, Z.; Tao, T.; Wang, H.; Chen, J.; Small, G.E.; Johnson, D.; Chen, J.; Zhang, Y.; Zhu, Q.; Zhang, S. Forms of nitrogen inputs regulate the intensity of soil acidification. Glob. Change Biol. 2023, 29, 4044–4055. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Shi, Z.; Kang, B.; Tu, H.; Zhu, J.; Li, H. Nitrogen addition and drought affect nitrogen uptake patterns and biomass production of four urban greening tree species in North China. Sci. Total Environ. 2023, 893, 164893. [Google Scholar] [CrossRef]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Using soil bacterial communities to predict physico-chemical variables and soil quality. Microbiome 2020, 8, 79. [Google Scholar] [CrossRef]
- Xia, Q.; Rufty, T.; Shi, W. Soil microbial diversity and composition: Links to soil texture and associated properties. Soil Biol. Biochem. 2020, 149, 107953. [Google Scholar] [CrossRef]
- Tajik, S.; Ayoubi, S.; Lorenz, N. Soil microbial communities affected by vegetation, topography and soil properties in a forest ecosystem. Appl. Soil Ecol. 2020, 149, 103514. [Google Scholar] [CrossRef]
- Shu, X.; Zhang, K.; Zhang, Q.; Wang, W. Changes in the composition of rhizosphere bacterial communities in response to soil types and acid rain. J. Environ. Manag. 2023, 325, 116493. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Cai, J.; Yang, S.; Li, S.; Shao, X.; Fu, C.; Li, C.; Deng, Y.; Huang, J.; Ruan, Y. Partial substitution of chemical fertilizer with organic fertilizer and slow-release fertilizer benefits soil microbial diversity and pineapple fruit yield in the tropics. Appl. Soil Ecol. 2023, 189, 104974. [Google Scholar] [CrossRef]
- Ijaz, F.; Ijaz, M.F.; Javed, H.; Amin, H.A.; Zafar, H.; Hamza, A.; Saleem, M.U.; Mujeeb, F.; Ehsan, S.; Alvi, A. Co-inoculation of Bradyrhizobium and Phosphate Solubilizing Microbes on Growth Promotion of Groundnut Under Rain-fed Conditions. J. Appl. Res. Plant Sci. 2023, 4, 348–355. [Google Scholar] [CrossRef]
- Gogoi, N.; Baruah, K.K.; Meena, R.S. Grain legumes: Impact on soil health and agroecosystem. In Legumes for Soil Health and Sustainable Management; Springer: Singapore, 2018; pp. 511–539. [Google Scholar]
- Zhang, Z.; Tariq, A.; Zeng, F.; Graciano, C.; Sun, F.; Chai, X.; Ahmed, Z. Nitrogen and water addition regulate fungal community and microbial co-occurrence network complexity in the rhizosphere of Alhagi sparsifolia seedlings. Appl. Soil Ecol. 2021, 164, 103940. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Q.; Dong, Y.; Guo, Y. The influence of increased precipitation and nitrogen deposition on the litter decomposition and soil microbial community structure in a semiarid grassland. Sci. Total Environ. 2022, 844, 157115. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, J.; Zhang, Y.; Dong, L.; Shangguan, Z.; Deng, L. Interactive effects of nitrogen and water addition on soil microbial resource limitation in a temperate desert shrubland. Plant Soil 2022, 475, 361–378. [Google Scholar] [CrossRef]
- Deng, L.; Peng, C.; Kim, D.-G.; Li, J.; Liu, Y.; Hai, X.; Liu, Q.; Huang, C.; Shangguan, Z.; Kuzyakov, Y. Drought effects on soil carbon and nitrogen dynamics in global natural ecosystems. Earth-Sci. Rev. 2021, 214, 103501. [Google Scholar] [CrossRef]
- Gao, D.; Bai, E.; Li, M.; Zhao, C.; Yu, K.; Hagedorn, F. Responses of soil nitrogen and phosphorus cycling to drying and rewetting cycles: A meta-analysis. Soil Biol. Biochem. 2020, 148, 107896. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Xie, F. Effect of drought stress at reproductive stages on growth and nitrogen metabolism in soybean. Agronomy 2020, 10, 302. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Sun, R.; Hu, S.; Qiao, Z.; Wang, S.; Mi, X. NH4+-N/NO3−-N ratio controlling nitrogen transformation accompanied with NO2−-N accumulation in the oxic-anoxic transition zone. Environ. Res. 2020, 189, 109962. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Qin, Y.; Wang, Y.; Liu, S.; Yu, B.; Song, Y.; Wang, X.; Zhu, G. Dissimilatory nitrate/nitrite reduction to ammonium (DNRA) pathway dominates nitrate reduction processes in rhizosphere and non-rhizosphere of four fertilized farmland soil. Environ. Res. 2020, 186, 109612. [Google Scholar] [CrossRef]
- Bowen, J.L.; Spivak, A.C.; Bernhard, A.E.; Fulweiler, R.W.; Giblin, A.E. Salt marsh nitrogen cycling: Where land meets sea. Trends Microbiol. 2023, 32, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.; Felgate, H.; Watmough, N.; Thomson, A.; Baggs, E. Mitigating release of the potent greenhouse gas N2O from the nitrogen cycle–could enzymic regulation hold the key? Trends Biotechnol. 2009, 27, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, E.M.; Franklin, R.B. Resource effects on denitrification are mediated by community composition in tidal freshwater wetlands soils. Environ. Microbiol. 2015, 17, 1520–1532. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Zheng, M.; Jiang, L.; Luo, Y. Patterns and mechanisms of responses by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Guo, G.; Kong, W.; Liu, J.; Zhao, J.; Du, H.; Zhang, X.; Xia, P. Diversity and distribution of autotrophic microbial community along environmental gradients in grassland soils on the Tibetan Plateau. Appl. Microbiol. Biotechnol. 2015, 99, 8765–8776. [Google Scholar] [CrossRef]
- He, W.; Zhang, M.; Jin, G.; Sui, X.; Zhang, T.; Song, F. Effects of nitrogen deposition on nitrogen-mineralizing enzyme activity and soil microbial community structure in a Korean pine plantation. Microb. Ecol. 2021, 81, 410–424. [Google Scholar] [CrossRef]
- Guoyong, Y.; Yajuan, X.; Shijie, H.; Zhang, J.; Qinggui, W.; Changcheng, M. Long-time precipitation reduction and nitrogen deposition increase alter soil nitrogen dynamic by influencing soil bacterial communities and functional groups. Pedosphere 2020, 30, 363–377. [Google Scholar]
- Bahulikar, R.A.; Chaluvadi, S.R.; Torres-Jerez, I.; Mosali, J.; Bennetzen, J.L.; Udvardi, M. Nitrogen fertilization reduces nitrogen fixation activity of diverse diazotrophs in switchgrass roots. Phytobiomes J. 2021, 5, 80–87. [Google Scholar] [CrossRef]
- Li, W.; Xie, L.; Zhao, C.; Hu, X.; Yin, C. Nitrogen fertilization increases soil microbial biomass and alters microbial composition especially under low soil water availability. Microb. Ecol. 2023, 86, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Kuerban, M.; Cong, W.-F.; Jing, J.; Bezemer, T.M. Microbial soil legacies of crops under different water and nitrogen levels determine succeeding crop performance. Plant Soil 2023, 485, 167–180. [Google Scholar] [CrossRef]
- Hagh-Doust, N.; Mikryukov, V.; Anslan, S.; Bahram, M.; Puusepp, R.; Dulya, O.; Tedersoo, L. Effects of nitrogen deposition on carbon and nutrient cycling along a natural soil acidity gradient as revealed by metagenomics. New Phytol. 2023, 238, 2607–2620. [Google Scholar] [CrossRef] [PubMed]
- Liao, L.; Wang, J.; Dijkstra, F.A.; Lei, S.; Zhang, L.; Wang, X.; Liu, G.; Zhang, C. Nitrogen enrichment stimulates rhizosphere multi-element cycling genes via mediating plant biomass and root exudates. Soil Biol. Biochem. 2024, 190, 109306. [Google Scholar] [CrossRef]
- Yang, X.; Ni, K.; Shi, Y.; Yi, X.; Ji, L.; Wei, S.; Jiang, Y.; Zhang, Y.; Cai, Y.; Ma, Q. Metagenomics reveals N-induced changes in carbon-degrading genes and microbial communities of tea (Camellia sinensis L.) plantation soil under long-term fertilization. Sci. Total Environ. 2023, 856, 159231. [Google Scholar] [CrossRef]
- Khalili, B.; Ogunseitan, O.A.; Goulden, M.L.; Allison, S.D. Interactive effects of precipitation manipulation and nitrogen addition on soil properties in California grassland and shrubland. Appl. Soil Ecol. 2016, 107, 144–153. [Google Scholar] [CrossRef]
- Philippot, L.; Chenu, C.; Kappler, A.; Rillig, M.C.; Fierer, N. The interplay between microbial communities and soil properties. Nat. Rev. Microbiol. 2024, 22, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, J.; Ma, T.; Lv, W.; Zhang, Z.; Shen, Y.; Yang, Q.; Wang, X.; Li, J.; Xiang, Q. Effects of short-term drought, nitrogen application and their interactions on the composition and functional genes of soil microbial communities in alfalfa grassland on the Loess Plateau. Front. Sustain. Food Syst. 2023, 7, 1332683. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Gao, X.; Wang, Q.; Cui, M.; Zhang, D.; Guo, P. Unveiling the driving role of pH on community stability and function during lignocellulose degradation in paddy soil. Front. Microbiol. 2024, 15, 1338842. [Google Scholar] [CrossRef]
- Xiang, J.; Gu, J.; Wang, G.; Bol, R.; Yao, L.; Fang, Y.; Zhang, H. Soil pH controls the structure and diversity of bacterial communities along elevational gradients on Huangshan, China. Eur. J. Soil Biol. 2024, 120, 103586. [Google Scholar] [CrossRef]
- Lü, C.Q.; Tian, H.Q. Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data. J. Geophys. Res. Atmos. 2007, 112, D22S05. [Google Scholar] [CrossRef]
- Xu, Z.Z.; Zhou, G.S.; Shimizu, H. Are plant growth and photosynthesis limited by pre-drought following rewatering in grass? J. Exp. Bot. 2009, 60, 3737–3749. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Wang, Y.J.; Pan, K.W.; Wang, Q.W.; Liang, J.; Jin, Y.Q.; Tariq, A. The synergistic responses of different photoprotective pathways in dwarf bamboo (Fargesia rufa) to drought and subsequent rewatering. Front. Plant Sci. 2017, 8, 489. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Noguchi, H.; Park, J.; Takagi, T. MetaGene: Prokaryotic gene finding from environmental genome shotgun sequences. Nucleic Acids Res. 2006, 34, 5623–5630. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).