
Characterization and Expression of the Cytochrome P450 Genes in Daphnia magna Exposed to Cerium Oxide Nanoparticles

Abstract
:1. Introduction
2. Results
2.1. Identification of CYP Genes Family Members in Daphnia magna
2.2. Chromosomal Distribution of CYPs in Daphnia magna
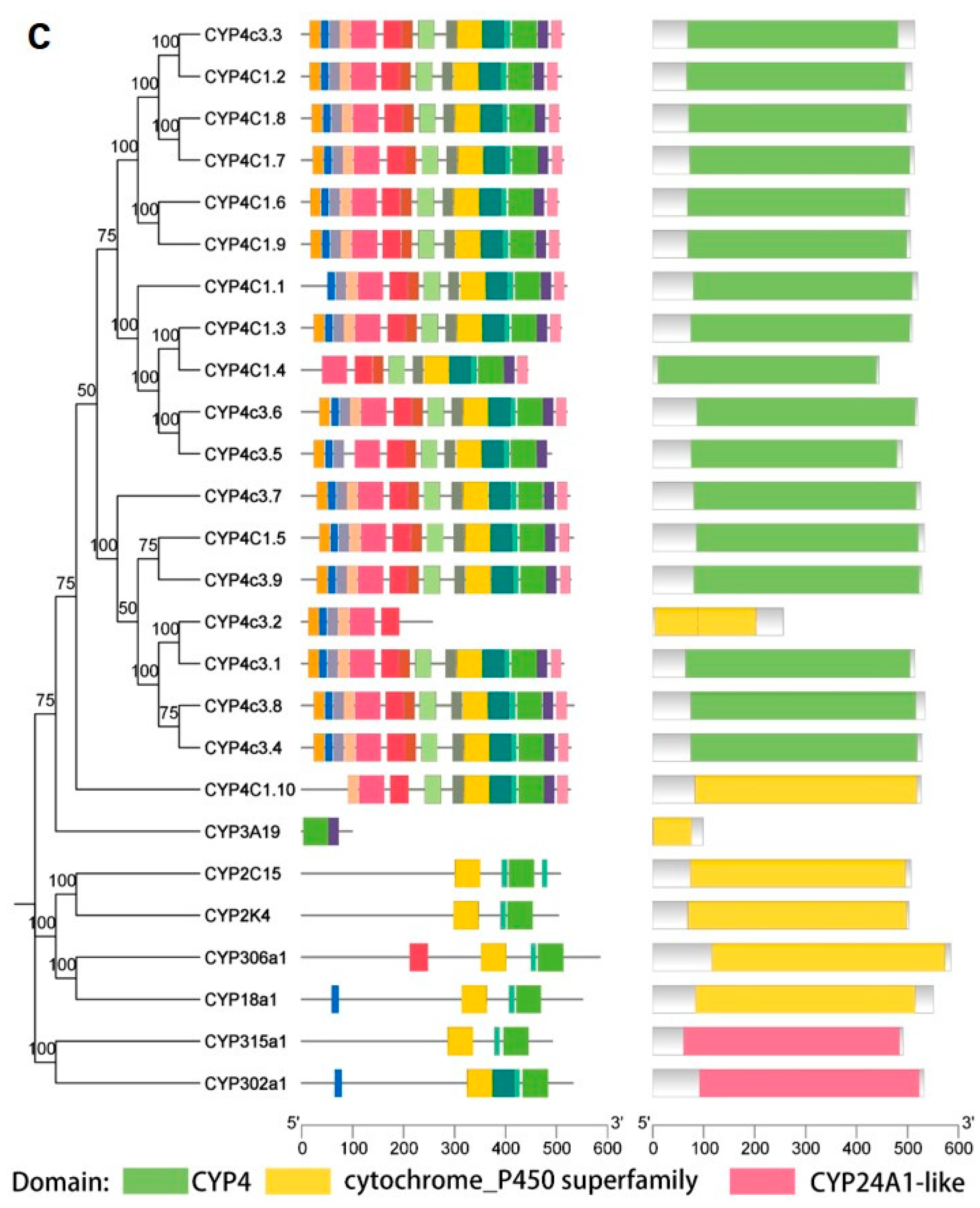
2.3. Phylogenetic Analysis of CYPs
2.4. Motifs and Structures of CYPs in Daphnia magna
2.5. Gene Expression Analysis of CYP Genes in Daphnia magna Exposed to nCeO2
3. Discussion
4. Materials and Methods
4.1. Experimental Materials and Toxicity Treatment
4.2. Identification of CYPs in Daphnia magna
4.3. Transcriptomic Analysis of CYPs in Daphnia magna Exposed to nCeO2
4.4. Chromosomal Distribution
4.5. Phylogenetic Tree
4.6. Structure and Motifs of CYP Genes in Daphnia magna
4.7. Quantitative Real-Time PCR (qPCR) Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Milenković, I.; Radotić, K.; Despotović, J.; Lončarević, B.; Lješević, M.; Spasić, S.Z.; Nikolić, A.; Beškoski, V.P. Toxicity investigation of CeO2 nanoparticles coated with glucose and exopolysaccharides levan and pullulan on the bacterium Vibrio fischeri and aquatic organisms Daphnia magna and Danio rerio. Aquat. Toxicol. 2021, 236, 105867. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Robertson, S.; Weber, R.J.M.; Buckley, A.; Warren, J.; Hodgson, A.; Rappoport, J.Z.; Ignatyev, K.; Meldrum, K.; Römer, I.; et al. Pulmonary toxicity of inhaled nano-sized cerium oxide aerosols in Sprague–Dawley rats. Nanotoxicology 2019, 13, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ma, Y.; Jiao, C.; Liu, M.; Luo, W.; Dong, C.; Fan, S.; He, X.; Yang, F.; Zhang, Z. Comparative toxicity of rod-shaped nano-CeO2 and nano-CePO4 to lettuce. Metallomics 2021, 13, mfab033. [Google Scholar] [CrossRef] [PubMed]
- Mauro, M.; Crosera, M.; Monai, M.; Montini, T.; Fornasiero, P.; Bovenzi, M.; Adami, G.; Turco, G.; Larese Filon, F. Cerium Oxide Nanoparticles Absorption through Intact and Damaged Human Skin. Molecules 2019, 24, 3759. [Google Scholar] [CrossRef]
- Villa, S.; Maggioni, D.; Hamza, H.; Di Nica, V.; Magni, S.; Morosetti, B.; Parenti, C.C.; Finizio, A.; Binelli, A.; Della Torre, C. Natural molecule coatings modify the fate of cerium dioxide nanoparticles in water and their ecotoxicity to Daphnia magna. Environ. Pollut. 2020, 257, 113597. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, Z.; Zhai, Y.; Zhang, F.; Vijver, M.G.; Peijnenburg, W.J.G.M. Effects of humic substances on the aqueous stability of cerium dioxide nanoparticles and their toxicity to aquatic organisms. Sci. Total Environ. 2021, 781, 146583. [Google Scholar] [CrossRef]
- Della Torre, C.; Maggioni, D.; Nigro, L.; Farè, F.; Hamza, H.; Protano, G.; Magni, S.; Fontana, M.; Riccardi, N.; Chiara, M.; et al. Alginate coating modifies the biological effects of cerium oxide nanoparticles to the freshwater bivalve Dreissena polymorpha. Sci. Total Environ. 2021, 773, 145612. [Google Scholar] [CrossRef]
- Correia, A.T.; Rodrigues, S.; Ferreira-Martins, D.; Nunes, A.C.; Ribeiro, M.I.; Antunes, S.C. Multi-biomarker approach to assess the acute effects of cerium dioxide nanoparticles in gills, liver and kidney of Oncorhynchus mykiss. Comp. Biochem. Physiol. C 2020, 238, 108842. [Google Scholar] [CrossRef] [PubMed]
- Jemec, A.; Djinović, P.; Tišler, T.; Pintar, A. Effects of four CeO2 nanocrystalline catalysts on early-life stages of zebrafish Danio rerio and crustacean Daphnia magna. J. Hazard. Mater. 2012, 219–220, 213–220. [Google Scholar] [CrossRef]
- Xia, J.; Zhao, H.Z.; Lu, G.H. Effects of selected metal oxide nanoparticles on multiple biomarkers in Carassius auratus. Biomed. Environ. Sci. BES 2013, 26, 742–749. [Google Scholar]
- Mahaye, N.; Musee, N. Evaluation of Apical and Molecular Effects of Algae Pseudokirchneriella subcapitata to Cerium Oxide Nanoparticles. Toxics 2023, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Li, X.; Hei, L.; Chen, Y.; Dong, Y.; Zhang, S.; Ma, S.; Xu, J.; Pang, Q.; Lynch, I.; et al. Toxicity of ceria nanoparticles to the regeneration of freshwater planarian Dugesia japonica: The role of biotransformation. Sci. Total Environ. 2023, 857, 159590. [Google Scholar] [CrossRef] [PubMed]
- Issac, M.N.; Kandasubramanian, B. Effect of microplastics in water and aquatic systems. Environ. Sci. Pollut. Res. 2021, 28, 19544–19562. [Google Scholar] [CrossRef]
- Gaiser, B.K.; Biswas, A.; Rosenkranz, P.; Jepson, M.A.; Lead, J.R.; Stone, V.; Tyler, C.R.; Fernandes, T.F. Effects of silver and cerium dioxide micro- and nano-sized particles on Daphnia magna. J. Environ. Monit. 2011, 13, 1227–1235. [Google Scholar] [CrossRef]
- Savuca, A.; Nicoara, M.N.; Ciobica, A.; Gorgan, D.L.; Ureche, D.; Balmus, I.M. Current Aspects on the Plastic Nano- and Microparticles Toxicity in Zebrafish—Focus on the Correlation between Oxidative Stress Responses and Neurodevelopment. Animals 2023, 13, 1810. [Google Scholar] [CrossRef] [PubMed]
- Sugantharaj David, E.M.D.; Madurantakam Royam, M.; Rajamani Sekar, S.K.; Manivannan, B.; Jalaja Soman, S.; Mukherjee, A.; Natarajan, C. Toxicity, uptake, and accumulation of nano and bulk cerium oxide particles in Artemia salina. Environ. Sci. Pollut. Res. 2017, 24, 24187–24200. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, X.; Xing, Z.; Xu, P.; Sun, J. Nano-Cerium Oxide Promotes Proliferation of Hepatoma Cells and Regulates mRNA Expression of Apoptosis-Related Genes Bcl-2 and Bax, as Detected Through Real-Time Fluorescent Quantitative Polymerase Chain Reaction. J. Nanosci. Nanotechnol. 2020, 20, 7457–7463. [Google Scholar] [CrossRef]
- Yao, M.; Ji, X.; Zhang, Y.; Mao, Z.; Chi, X. miR-99 family is potential target to reverse cerium dioxide nanoparticle-induced placental cell dysfunction. Ann. Transl. Med. 2022, 10, 402. [Google Scholar] [CrossRef]
- Sökmen, T.Ö.; Sulukan, E.; Türkoğlu, M.; Baran, A.; Özkaraca, M.; Ceyhun, S.B. Polystyrene nanoplastics (20 nm) are able to bioaccumulate and cause oxidative DNA damages in the brain tissue of zebrafish embryo (Danio rerio). Neurotoxicology 2020, 77, 51–59. [Google Scholar] [CrossRef]
- Morel, E.; Dozois, J.; Slaveykova, V.I.; Wilkinson, K.J. Distinguishing the effects of Ce nanoparticles from their dissolution products: Identification of transcriptomic biomarkers that are specific for ionic Ce in Chlamydomonas reinhardtii. Metallomics 2021, 13, mfaa005. [Google Scholar] [CrossRef]
- Zhang, Z.; Wen, Z.; Li, K.; Xu, W.; Liang, N.; Yu, X.; Li, C.; Chu, D.; Guo, L. Cytochrome P450 Gene, CYP6CX3, Is Involved in the Resistance to Cyantraniliprole in Bemisia tabaci. J. Agric. Food Chem. 2022, 70, 12398–12407. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Lee, K.W. Identification and response of cytochrome P450 genes in the brackish water flea Diaphanosoma celebensis after exposure to benzo[α]pyrene and heavy metals. Mol. Biol. Rep. 2021, 48, 657–664. [Google Scholar] [CrossRef]
- Zhang, L.; Xie, Z.; Liu, Z.; Zhou, S.; Ma, L.; Liu, W.; Huang, J.-W.; Ko, T.-P.; Li, X.; Hu, Y.; et al. Structural insight into the electron transfer pathway of a self-sufficient P450 monooxygenase. Nat. Commun. 2020, 11, 2676. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Ma, J.; Li, M.; Zhang, Y.; Jiang, B.; Zhao, X.; Huai, C.; Shen, L.; Zhang, N.; He, L.; et al. Cytochrome P450 Enzymes and Drug Metabolism in Humans. Int. J. Mol. Sci. 2021, 22, 12808. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Park, J.C.; Hagiwara, A.; Park, H.G.; Lee, J.-S. Identification of the full 26 cytochrome P450 (CYP) genes and analysis of their expression in response to benzo[α]pyrene in the marine rotifer Brachionus rotundiformis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 29, 185–192. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, W.; Zhang, Y.; Cui, Y.; Xu, S.; Li, S. Bisphenol A regulates cytochrome P450 1B1 through miR-27b-3p and induces carp lymphocyte oxidative stress leading to apoptosis. Fish Shellfish Immunol. 2020, 102, 489–498. [Google Scholar] [CrossRef]
- Risalde, M.A.; Molina, A.M.; Lora, A.J.; Ayala, N.; Gómez-Villamandos, J.C.; Moyano, M.R. Immunohistochemical expression of aromatase cyp19a1a and cyp19a1b in the ovary and brain of zebrafish (Danio rerio) exposed to different concentrations of bisphenol A. Aquat. Toxicol. 2021, 237, 105876. [Google Scholar] [CrossRef]
- Rusni, S.; Sassa, M.; Takagi, T.; Kinoshita, M.; Takehana, Y.; Inoue, K. Establishment of cytochrome P450 1a gene-knockout Javanese medaka, Oryzias javanicus, which distinguishes toxicity modes of the polycyclic aromatic hydrocarbons, pyrene and phenanthrene. Mar. Pollut. Bull. 2022, 178, 113578. [Google Scholar] [CrossRef]
- Liu, C.; Li, J.; Qi, X.; Wang, L.; Sun, D.; Zhang, J.; Zhang, K.; Li, J.; Li, Y.; Wen, H. Cytochrome P450 superfamily in spotted sea bass: Genome-wide identification and expression profiles under trichlorfon and environmental stresses. Comp. Biochem. Physiol. D 2023, 46, 101078. [Google Scholar] [CrossRef]
- Woo, S.J.; Chung, J.K. Cytochrome P450 1 enzymes in black rockfish, Sebastes schlegelii: Molecular characterization and expression patterns after exposure to benzo[a]pyrene. Aquat. Toxicol. 2020, 226, 105566. [Google Scholar] [CrossRef]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.V.; Jugulam, M. Role of Cytochrome P450 Enzymes in Plant Stress Response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef]
- Baldwin, W.S.; Marko, P.B.; Nelson, D.R. The cytochrome P450 (CYP) gene superfamily in Daphnia pulex. BMC Genom. 2009, 10, 169. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.L.; Fitzgibbon, Q.P.; Smith, G.G.; Elizur, A.; Ventura, T. Ontogeny of the Cytochrome P450 Superfamily in the Ornate Spiny Lobster (Panulirus ornatus). Int. J. Mol. Sci. 2024, 25, 1070. [Google Scholar] [CrossRef] [PubMed]
- Loerracher, A.-K.; Braunbeck, T. Cytochrome P450-dependent biotransformation capacities in embryonic, juvenile and adult stages of zebrafish (Danio rerio)—A state-of-the-art review. Arch. Toxicol. 2021, 95, 2299–2334. [Google Scholar] [CrossRef] [PubMed]
- Vizzini, A.; Bonura, A.; La Paglia, L.; Fiannaca, A.; La Rosa, M.; Urso, A.; Mauro, M.; Vazzana, M.; Arizza, V. Transcriptomic Analyses Reveal 2 and 4 Family Members of Cytochromes P450 (CYP) Involved in LPS Inflammatory Response in Pharynx of Ciona robusta. Int. J. Mol. Sci. 2021, 22, 11141. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Liu, Q.; Chen, D.; Liu, Y. Atrazine exposure induces necroptosis through the P450/ROS pathway and causes inflammation in the gill of common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2022, 131, 809–816. [Google Scholar] [CrossRef]
- Awali, S.; Abdulelah, S.A.; Crile, K.G.; Yacoo, K.E.; Almouseli, A.; Torres, V.C.; Dayfield, D.J.; Evans, K.R.; Belanger, R.M. Cytochrome P450 and Glutathione-S-Transferase Activity are Altered Following Environmentally Relevant Atrazine Exposures in Crayfish (Faxoniusvirilis). Bull. Environ. Contam. Toxicol. 2019, 103, 579–584. [Google Scholar] [CrossRef]
- Meng, Q.; Yeung, K.; Kwok, M.L.; Chung, C.T.; Hu, X.L.; Chan, K.M. Toxic effects and transcriptome analyses of zebrafish (Danio rerio) larvae exposed to benzophenones. Environ. Pollut. 2020, 265, 114857. [Google Scholar] [CrossRef]
- Zacchi, F.L.; de Lima, D.; Flores-Nunes, F.; Mattos, J.J.; Lüchmann, K.H.; de Miranda Gomes, C.H.A.; Bícego, M.C.; Taniguchi, S.; Sasaki, S.T.; Dias Bainy, A.C. Transcriptional changes in oysters Crassostrea brasiliana exposed to phenanthrene at different salinities. Aquat. Toxicol. 2017, 183, 94–103. [Google Scholar] [CrossRef]
- Zhang, J.; He, Y.; Yan, X.; Qu, C.; Li, J.; Zhao, S.; Wang, X.; Guo, B.; Liu, H.; Qi, P. Two novel CYP3A isoforms in marine mussel Mytilus coruscus: Identification and response to cadmium and benzo[a]pyrene. Aquat. Toxicol. 2019, 214, 105239. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, S.; Li, Y.; Wu, G.; Sun, Y.; Zhang, L.; Huang, Y.; Lyu, K.; Chen, Y.; Yang, Z. Combined effects of ZnO nanoparticles and toxic Microcystis on life-history traits of Daphnia magna. Chemosphere 2019, 233, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.J.; An, H.E.; Kim, C.B. Machine Learning Models for Identification and Prediction of Toxic Organic Compounds Using Daphnia magna Transcriptomic Profiles. Life 2022, 12, 1443. [Google Scholar] [CrossRef]
- Renzi, M.; Blašković, A. Ecotoxicity of nano-metal oxides: A case study on Daphnia magna. Ecotoxicology 2019, 28, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Andreani, T.; Nogueira, V.; Gavina, A.; Fernandes, S.; Rodrigues, J.L.; Pinto, V.V.; Ferreira, M.J.; Silva, A.M.; Pereira, C.M.; Pereira, R. Ecotoxicity to Freshwater Organisms and Cytotoxicity of Nanomaterials: Are We Generating Sufficient Data for Their Risk Assessment? Nanomaterials 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Eghan, K.; Lee, S.; Kim, W.K. Cardio- and neuro-toxic effects of four parabens on Daphnia magna. Ecotoxicol. Environ. Saf. 2023, 268, 115670. [Google Scholar] [CrossRef]
- Silva, M.L.N.d.; Nogueira, D.J.; Vicentini, D.S.; Puerari, R.C.; Alves, P.R.L.; Fuzinatto, C.F.; Matias, W.G. Rapid Communication: Oxidative stress induced by mixed exposure to glyphosate and silver nanoparticles. J. Toxicol. Environ. Health A 2022, 85, 586–590. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.L.N.; Nogueira, D.J.; Köerich, J.S.; Vaz, V.P.; Justino, N.M.; Schmidt, J.R.A.; Vicentini, D.S.; Matias, M.S.; de Castilhos, A.B.; Fuzinatto, C.F.; et al. Multigenerational Toxic Effects on Daphnia magna Induced by Silver Nanoparticles and Glyphosate Mixture. Environ. Toxicol. Chem. 2021, 40, 1123–1131. [Google Scholar] [CrossRef]
- Shen, C.; Wei, J.; Wang, T.; Wang, Y. Acute toxicity and responses of antioxidant systems to dibutyl phthalate in neonate and adult Daphnia magna. PeerJ 2019, 7, e6584. [Google Scholar] [CrossRef]
- Religia, P.; Nguyen, N.D.; Nong, Q.D.; Matsuura, T.; Kato, Y.; Watanabe, H. Mutation of the Cytochrome P450 CYP360A8 Gene Increases Sensitivity to Paraquat in Daphnia magna. Environ. Toxicol. Chem. 2021, 40, 1279–1288. [Google Scholar] [CrossRef]
- Ohnuki, S.; Tokishita, S.; Kojima, M.; Fujiwara, S. Effect of chlorpyrifos-exposure on the expression levels of CYP genes in Daphnia magna and examination of a possibility that an up-regulated clan 3 CYP, CYP360A8, reacts with pesticides. Environ. Toxicol. 2024, 39, 3641–3653. [Google Scholar] [CrossRef]
- Im, H.; Kang, J.; Jacob, M.F.; Bae, H.; Oh, J.-E. Transgenerational effects of benzotriazole on the gene expression, growth, and reproduction of Daphnia magna. Environ. Pollut. 2023, 323, 121211. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gong, Q.; Zhao, C.; Wang, D.; Ye, X.; Zheng, G.; Wang, Y.; Cao, J.; Sun, C. Genome-wide analysis of cytochrome P450 genes in Citrus clementina and characterization of a CYP gene encoding flavonoid 3′-hydroxylase. Hortic. Res. 2023, 10, uhac283. [Google Scholar] [CrossRef] [PubMed]
- Garaud, M.; Trapp, J.; Devin, S.; Cossu-Leguille, C.; Pain-Devin, S.; Felten, V.; Giamberini, L. Multibiomarker assessment of cerium dioxide nanoparticle (nCeO2) sublethal effects on two freshwater invertebrates, Dreissena polymorpha and Gammarus roeseli. Aquat. Toxicol. 2015, 158, 63–74. [Google Scholar] [CrossRef]
- Rundle, A.; Robertson, A.B.; Blay, A.M.; Butler, K.M.A.; Callaghan, N.I.; Dieni, C.A.; MacCormack, T.J. Cerium oxide nanoparticles exhibit minimal cardiac and cytotoxicity in the freshwater fish Catostomus commersonii. Comp. Biochem. Physiol. C 2016, 181–182, 19–26. [Google Scholar] [CrossRef]
- Guo, H.; Kuang, Y.; Ouyang, K.; Zhang, C.; Yang, H.; Chen, S.; Tang, R.; Zhang, X.; Li, D.; Li, L. Ammonia in the presence of nano titanium dioxide (nano-TiO2) induces greater oxidative damage in the gill and liver of female zebrafish. Ecotoxicol. Environ. Saf. 2022, 236, 113458. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.Y.M.; Alshagga, M.; Kong, C.; Alshawsh, M.A.; Alshehade, S.A.; Pan, Y. CYP35 family in Caenorhabditis elegans biological processes: Fatty acid synthesis, xenobiotic metabolism, and stress responses. Arch. Toxicol. 2022, 96, 3163–3174. [Google Scholar] [CrossRef]
- Dermauw, W.; Van Leeuwen, T.; Feyereisen, R. Diversity and evolution of the P450 family in arthropods. Insect Biochem. Mol. Biol. 2020, 127, 103490. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Liu, Z.; Cai, M.; Jiao, Y.; Li, Y.; Chen, Q.; Zhao, Y. Molecular characterisation of cytochrome P450 enzymes in waterflea (Daphnia pulex) and their expression regulation by polystyrene nanoplastics. Aquat. Toxicol. 2019, 217, 105350. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Ong, C.E.; Pung, Y.F.; Chieng, J.Y. The current understanding of the interactions between nanoparticles and cytochrome P450 enzymes—A literature-based review. Xenobiotica 2018, 49, 863–876. [Google Scholar] [CrossRef]
- Melo de Almeida, E.; Tisserand, F.; Faria, M.; Chèvre, N. Efficiency of Several Cytochrome P450 Biomarkers in Highlighting the Exposure of Daphnia magna to an Organophosphate Pesticide. Toxics 2022, 10, 482. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, X.; Liu, S.; Zhao, C.; Miao, Y.; Jin, L.; Wang, D.; Zhou, L. Cyp17a1 is Required for Female Sex Determination and Male Fertility by Regulating Sex Steroid Biosynthesis in Fish. Endocrinology 2021, 162, bqab205. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Wang, H.; Zeng, Q. Comparative transcriptome reveals the response of oriental river prawn (Macrobrachium nipponense) to sulfide toxicity at molecular level. Aquat. Toxicol. 2021, 230, 105700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, P.; Sun, L.; Cao, C. Integration of miRNA and mRNA expression profiles in Asian spongy moth Lymantria dispar in response to cyantraniliprole. Pestic. Biochem. Physiol. 2023, 191, 105364. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, W.; Chen, X.; Wang, M.; Zhong, L.; Liu, J.; Bian, W.; Zhang, S. Genome-wide identification and expression analysis of mitogen-activated protein kinase (MAPK) genes in response to salinity stress in channel catfish (Ictalurus punctatus). J. Fish Biol. 2022, 101, 972–984. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clan | Gene Name | Gene ID | Molecular Function |
|---|---|---|---|
| Clan 2 | CYP2C15 | LOC116918329 | steroid hydroxylase activity |
| CYP2K4 | LOC116923437 | steroid hydroxylase activity | |
| CYP18a1 | LOC116919046 | steroid hydroxylase activity | |
| CYP306a1 | LOC116918853 | steroid hydroxylase activity; steroid hydroxylase activity | |
| Clan 3 | CYP3A19 | LOC116928063 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen |
| Clan 4 | CYP4C1.1 | LOC116927068 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen |
| CYP4C1.2 | LOC116926713 | heme binding, iron ion binding, methyltransferase activity, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen, | |
| CYP4C1.3 | LOC116926407 | iron ion binding; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; oxidation-reduction process | |
| CYP4C1.4 | LOC116926406 | iron ion binding; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; oxidation-reduction process; oxidation-reduction process | |
| CYP4C1.5 | LOC116926459 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4C1.6 | LOC116928122 | iron ion binding; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; oxidation-reduction process | |
| CYP4C1.7 | LOC116927585 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4C1.8 | LOC116928121 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4C1.9 | LOC116927935 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4C1.10 | LOC116930969 | iron ion binding; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; oxidation-reduction process; oxidation-reduction process | |
| CYP4c3.1 | LOC116929157 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4c3.2 | LOC116922314 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4c3.3 | LOC116925875 | iron ion binding; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; oxidation-reduction process | |
| CYP4c3.4 | LOC116926314 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4c3.5 | LOC116934506 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4c3.6 | LOC116934248 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4c3.7 | LOC116926402 | iron ion binding; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; oxidation-reduction process | |
| CYP4c3.8 | LOC116926463 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| CYP4c3.9 | LOC116926461 | heme binding, iron ion binding, monooxygenase activity, oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen | |
| Mitochondrial clan | CYP302a1 | LOC116927720 | iron ion binding; mitochondrion; ecdysone biosynthetic process; dorsal closure; central nervous system development; midgut development; head involution; chitin-based embryonic cuticle biosynthetic process; electron transfer activity; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; ecdysteroid 22-hydroxylase activity; oxidation-reduction process |
| CYP315a1 | LOC116926680 | iron ion binding; mitochondrion; ecdysone biosynthetic process; dorsal closure; central nervous system development; midgut development; motor neuron axon guidance; head involution; oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen; heme binding; ecdysteroid 2-hydroxylase activity; oxidation-reduction process |
| Gene Name | Chromosome Location | Length (aa) | Isoelectric Points | Molecular Weight/Da | Subcellular Localizations | Exon Count | Gravy |
|---|---|---|---|---|---|---|---|
| CYP18a1 | LG1 | 551 | 8.35 | 63,170.02 | extracellular | 6 | −0.122 |
| CYP4C1.9 | LG7 | 506 | 8.72 | 58,694.89 | plasma membrane | 18 | −0.17 |
| CYP2C15 | LG3 | 507 | 9.19 | 58,789.94 | endoplasmic reticulum | 9 | −0.181 |
| CYP4C1.6 | LG7 | 504 | 6.55 | 58,039.1 | plasma membrane | 12 | −0.119 |
| CYP4c3.1 | Unknown | 514 | 8.23 | 58,912.73 | mitochondrion | 13 | −0.13 |
| CYP4C1.4 | LG7 | 444 | 8.43 | 50,380.05 | mitochondrion | 14 | −0.15 |
| CYP4c3.4 | LG7 | 528 | 7.17 | 60,877.57 | endoplasmic reticulum | 13 | −0.141 |
| CYP4c3.7 | LG7 | 526 | 6.3 | 59,991.96 | plasma membrane | 14 | −0.18 |
| CYP306a1 | LG1 | 585 | 8.45 | 66,441.76 | plasma membrane | 9 | −0.159 |
| CYP4C1.1 | LG7 | 520 | 6.22 | 60,095.44 | plasma membrane | 16 | −0.114 |
| CYP4C1.2 | LG7 | 509 | 6.75 | 58,800.96 | endoplasmic reticulum | 11 | −0.173 |
| CYP4c3.5 | LG7 | 490 | 7.87 | 55,885.2 | plasma membrane | 15 | −0.097 |
| CYP4C1.10 | LG9 | 527 | 7.61 | 60,180.6 | endoplasmic reticulum | 12 | −0.054 |
| CYP3A19 | LG7 | 99 | 9.16 | 11,683.6 | extracellular | 4 | −0.363 |
| CYP4c3.9 | LG7 | 528 | 6.1 | 60,656.91 | endoplasmic reticulum | 11 | −0.15 |
| CYP4C1.3 | LG7 | 509 | 5.93 | 58,157.05 | plasma membrane | 12 | −0.053 |
| CYP4c3.8 | LG7 | 534 | 8.58 | 62,191.37 | endoplasmic reticulum | 13 | −0.159 |
| CYP4c3.3 | LG6 | 514 | 6.9 | 59,866.09 | endoplasmic reticulum | 11 | −0.263 |
| CYP2K4 | LG5 | 503 | 7.17 | 58,164.88 | endoplasmic reticulum | 9 | −0.191 |
| CYP4c3.6 | LG7 | 520 | 6.44 | 58,821.43 | plasma membrane | 14 | 0.058 |
| CYP4C1.8 | LG7 | 507 | 7.6 | 58,335.33 | plasma membrane | 11 | −0.173 |
| CYP4C1.7 | LG7 | 513 | 8.43 | 58,957.01 | endoplasmic reticulum | 13 | −0.209 |
| CYP4C1.5 | LG7 | 533 | 6.65 | 60,999.32 | plasma membrane | 11 | −0.135 |
| CYP4c3.2 | LG5 | 256 | 5.98 | 28,663.33 | extracellular | 6 | −0.001 |
| CYP315a1 | LG7 | 491 | 8.77 | 56,522.29 | mitochondrion | 8 | −0.229 |
| CYP302a1 | LG7 | 532 | 9.43 | 60,986.62 | mitochondrion | 8 | −0.269 |
| Gene ID | Description | Gene | Forward Primer (5′-3′) Reverse Primer (3′-5′) |
|---|---|---|---|
| Beta-actin | Beta-actin | CCCCATTTATGAAGGTTACGC CCTTGATGTCACGGACGATTT | |
| 116919046 | cytochrome P450 18a1 | CYP18a1 | TCACCATACCGAAAGGCACC ACGCCGAACGGAATGAAGTA |
| 116927935 | cytochrome P450 4C1.9 | CYP4C1.9 | ATATGCCCTCCACCACAACG TCCAATGCAGTTCCTCGGTC |
| 116918329 | cytochrome P450 2C15 | CYP2C15 | TCATACTCACAGCGAACGCA ATCATGCCAAAGGGCAGTGT |
| 116928122 | cytochrome P450 4C1.6 | CYP4C1.6 | TTCGGCAAGTGGACGACATC CCTCGACATCGGAAGGAGAC |
| 116929157 | cytochrome P450 4c3.1 | CYP4c3.1 | TGGCTTCGATTGGATTGGCT ACCAATTCACGGCGATGTCT |
| 116926406 | cytochrome P450 4C1.4 | CYP4C1.4 | ATCACCAGAATTTATGGAGGTAAGT CCGGGACTAATAAGAAGCCCT |
| 116926314 | cytochrome P450 4c3.4 | CYP4c3.4 | TACCTGATGGCGAAGCATCC GGATGCCAGGGTACAACCTC |
| 116926402 | cytochrome P450 4c3.7 | CYP4c3.7 | CAGAGAACAGCATCGGTCGT CATAGCTGGATCAGAGGCGG |
| 116918853 | cytochrome P450 306a1 | CYP306a1 | TTTGCCCAGTTACCCAGTTGT TGCAACCACCATACGGCGA |
| 116927068 | cytochrome P450 4C1.1 | CYP4C1.1 | TAATAACGCGGGGCTGAGTG TCGCTGATGTTGTCGTGTCA |
| 116926713 | cytochrome P450 4C1.2 | CYP4C1.2 | CATCAAGCGCCGTGTTAGTG ACGGAACGAAGGCAAATGGA |
| 116934506 | cytochrome P450 4c3.5 | CYP4c3.5 | GTCGGTGCTGTGGGATTGTT CGTATCTTCCCGTCCACGTT |
| 116930969 | cytochrome P450 4C1.10 | CYP4C1.10 | TGGAAACCCAAAAGGACGGT ACAGCTTGAGGGTATTCGCC |
| 116928063 | cytochrome P450 3A19 | CYP3A19 | ATGCCGGCGTATGCTCTC TTTCAGCGCACCATCTTTCG |
| 116926461 | cytochrome P450 4c3.9 | CYP4c3.9 | CCGGAGAACAGTATCGGACG TCATGGGCTGTGAGGAATCG |
| 116926407 | cytochrome P450 4C1.3 | CYP4C1.3 | TCTTGATGATATGTGCGCGTC GCGCGTTTATTCTTGTCCCC |
| 116926463 | cytochrome P450 4c3.8 | CYP4c3.8 | TAACACCGGCATTCCACGTT ATGTCAAGGGCGTGTTCCAT |
| 116925875 | cytochrome P450 4c3.3 | CYP4c3.3 | CTGACTCCGGCCTTCCATTT ATGTCGAGGGTGCATCGTTT |
| 116923437 | cytochrome P450 2K4 | CYP2K4 | CTGAAGCTCGCCAAATGGTC CTCGCTAAACTTGTCCGCCT |
| 116934248 | cytochrome P450 4c3.6 | CYP4c3.6 | GGTCCCAGGT TCTACTTGCC TAGCAATGCGAGCCAAGGAA |
| 116928121 | cytochrome P450 4C1.8 | CYP4C1.8 | AATTTGGCCGCATCTACCGA TTCCACTTAGCCCCTGTTGC |
| 116927585 | cytochrome P450 4C1.7 | CYP4C1.7 | AGGGTGGAGTGGAGCTTAGT TGATGCCCTGGATTCGTAGC |
| 116926459 | cytochrome P450 4C1.5 | CYP4C1.5 | CAAGCGTCGGAGAATGGAGA AGAAACCAACTCATCGCCGT |
| 116922314 | cytochrome P450 4c3.2 | CYP4c3.2 | AGAACATGCACGCTGAACAAC CTGGATCAGGGCTAACCTCC |
| 116926680 | cytochrome P450 315a1 | CYP315a1 | AATGGAACGACCACCACCAT TGCACTTGGAACGTGCAATC |
| 16927720 | cytochrome P450 302a1 | CYP302a1 | TTGGGACGGTCTCTGTGTTG GGTAGTGGTTTAGGGCAAGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, X.; Zhou, Y.; Liu, Q.; Liu, M.; Chen, J.; Zhang, Y.; Wei, J.; Wang, Y. Characterization and Expression of the Cytochrome P450 Genes in Daphnia magna Exposed to Cerium Oxide Nanoparticles. Int. J. Mol. Sci. 2024, 25, 10812. https://doi.org/10.3390/ijms251910812
Kang X, Zhou Y, Liu Q, Liu M, Chen J, Zhang Y, Wei J, Wang Y. Characterization and Expression of the Cytochrome P450 Genes in Daphnia magna Exposed to Cerium Oxide Nanoparticles. International Journal of Molecular Sciences. 2024; 25(19):10812. https://doi.org/10.3390/ijms251910812
Chicago/Turabian StyleKang, Xinyi, Yan Zhou, Qi Liu, Miao Liu, Jing Chen, Yuanwen Zhang, Jie Wei, and Yuan Wang. 2024. "Characterization and Expression of the Cytochrome P450 Genes in Daphnia magna Exposed to Cerium Oxide Nanoparticles" International Journal of Molecular Sciences 25, no. 19: 10812. https://doi.org/10.3390/ijms251910812